
Typha lepechinii Mavrodiev et Kapit. sp. nov. (Typhaceae Juss.)—A New Endangered Endemic Cattail in the Outmost East of European Russia †

 ,
,  and
and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
3. Results
3.1. Taxonomic Treatment
3.2. Diagnosis
3.3. Etymology
3.4. Material Examined
- Type
- Paratypes
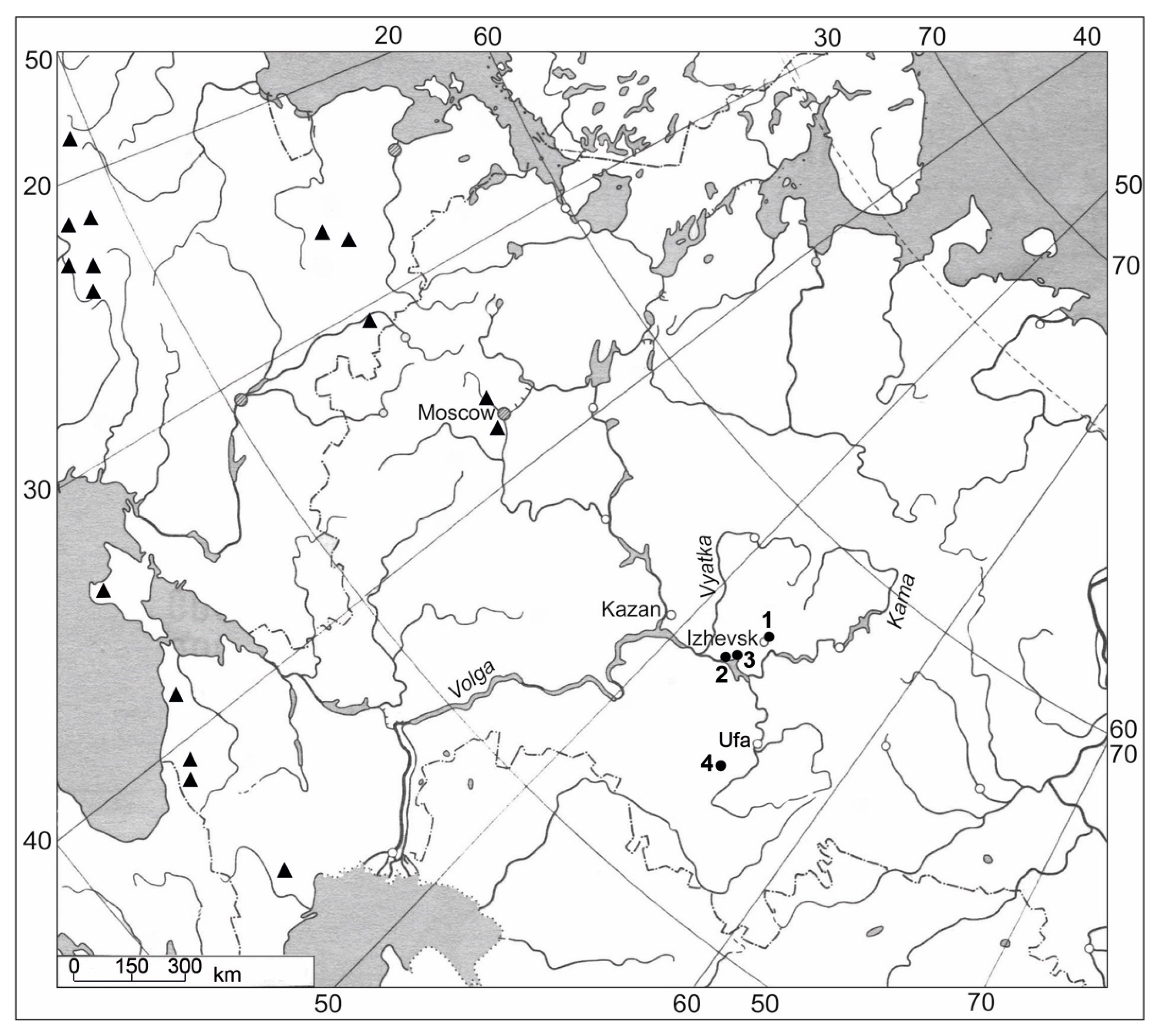
- RUSSIA—Udmurt Republic • Alnashsky district, five km south of the village Muvazhi; ~56°01′ N, 52°42′ E; shallow water of a small pond on river Golyushurminka; 1 July 2006; V. I. Kapitonov, O. A. Kapitonova (TOB, n 6540);
- RUSSIA—Republic of Tatarstan • Mendeleev district, vicinity of village Izhevka (1.5 km N-NW from the village), small creek dam, in shallow riverside water and on the damp shore; 28 June 2006; V. I. Kapitonov, O. A. Kapitonova (TOB, n 6539);
- RUSSIA—Republic of Bashkortostan • Miyakinsky district; 53°31′57.70″ N, 54°50′28.73″ E; on the damp shore along the stream Kurmanay, 1.9 km from the village Kurmanaibash; 23 July 2014; A. A. Muldashev (UFA, n 289).
- Description
3.5. Distribution and Ecology
3.6. Conservation Status
3.7. Cladistic Analyses of cpDNA Sequence Data
3.7.1. Standard cpDNA Matrix (No Indels Included)
- Typha lepechinii sp. nov. (Pattern I) is sister of T. shuttleworthii (Pattern II);
- Clade T. lepechinii sp. nov. plus T. shuttleworthii defined as a sister of all the remaining analyzed taxa of T. sect. Ebracteolatae;
- The clade that corresponds to Pattern III, is a sister of the clade that includes sequences of Patterns IV and V, but Pattern IV is defined as a polytomy by Fitch parsimony as well as by 3TA.
- In all the analyses T. latifolia appears as clearly polyphyletic.
3.7.2. Indel-Based Binary Matrix
- Typha shuttleworthii, not the clade (T. shuttleworthii + T. lepechinii sp. nov.), is defined as a sister of all the remaining taxa of T. sect. Ebracteolatae;
- Typha lepechinii sp. nov. is a sister of all the remaining analyzed taxa of sect. Ebracteolatae, excluding T. shuttleworthii;
- The clade that corresponds to Pattern III, is a sister of the clade that includes most sequences of Patterns IV (but not T. capsica) and V, but both Patterns are again defined as polytomies in all analyses;
- Clade (Pattern IV + Pattern V) is defined as a polytomy by Fitch parsimony as well as by 3TA;
- Pattern IV appears as paraphyletic due to the sisterhood of T. capsica and the clade (Patterns III + IV + V);
- In all the analyses of the indel-based binary matrix, T. latifolia appears as clearly polyphyletic (Figure 11B).
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Mavrodiev, E.V.; Kapitonova, O.A. Taxonomic composition of Typhaceae of the flora of European Russia. Novit. Syst. Plant. Vasc. 2015, 46, 5–24. Available online: https://www.binran.ru/files/journals/Novitates/2015_46/NSPV-46_1-Mavrodiev.pdf (accessed on 1 November 2020).
- Mavrodiev, E.V.; Majorov, S.R. Floristic records in Central and North-Western regions of East Europe. Bull. Mosc. Soc. Nat. Biol. Ser. 1999, 104, 61–62. [Google Scholar]
- Kozłowska, K.; Nobisk, A.; Nobis, M. Typha shuttleworthii (Typhaceae): New for Poland. Pol. Bot. J. 2011, 56, 299–305. [Google Scholar]
- Nobis, M.; Nobis, A.; Jędrzejczak, E.; Klichowska, E. A new record of Typha shuttleworthii (Typhaceae) in Poland. Acta Musei Sil. Sci. Nat. 2015, 64, 107–109. [Google Scholar] [CrossRef]
- Hrivnák, R.; Slezák, M.; Blanár, D.; Širka, P.; Šumberová, K. Vegetation affinity of species Typha shuttleworthii in the western part of the Carpathians, with Typhetum shuttleworthii as a new association to Slovakia. Biodivers. Data J. 2020, 8, e52151. [Google Scholar] [CrossRef]
- Hamdi, S.M.M.; Assadi, M.; Ebadi, M. Revision of study of Typha genus: Three new records species of the genus Typha (Typhaceae) in Iran and their micromorphological pollen and capsule studies. Asian J. Plant Sci. 2009, 8, 455–464. [Google Scholar] [CrossRef]
- Ghahreman, A.; Chariat-Panahi, M.S. Une espece nouvelle de Typha (Typhaceae) en Iran. Bull. de la Soc. Bot. de France. Lett. Bot. 1979, 126, 373–375. [Google Scholar]
- Mavrodiev, E.V. A new species of cat-tail (Typha L.) from sect. Engleria (Leonova) Tzvel. Feddes Repert. 2000, 111, 571–575. [Google Scholar] [CrossRef]
- Dubovik, D.V. Typhaceae Juss. In Flora of Belarus: Vascular Plants; Parfenov, V.I., Ed.; Belarusian Science Press: Minsk, Belarus, 2013; Volume 2, pp. 13–23. [Google Scholar]
- Kapitonova, O.A.; Platunova, G.R.; Kapitonov, V.I. The distribution, biological and ecological features of Typha shuttleworthii (Typhaceae) in the Vyatka-Kama Cis-Urals, Russia. Am. J. Plant Sci. 2015, 6, 283–288. [Google Scholar] [CrossRef] [Green Version]
- Kapitonova, O.A. Macrophyte Flora of the Vyatka-Kama Cis-Urals; Filigran: Yaroslavl, Russia, 2021. [Google Scholar]
- Zhou, B.; Tu, T.; Kong, F.; Wen, J.; Xu, X. Revised phylogeny and historical biogeography of the cosmopolitan aquatic plant genus Typha (Typhaceae). Sci. Rep. 2018, 8, 8813. [Google Scholar] [CrossRef]
- Volkova, P.A.; Bobrov, A.A. Easier than it looks: Notes on the taxonomy of Typha L. (Typhaceae) in East Europe. Aquat. Bot. 2022, 176, 103453. [Google Scholar] [CrossRef]
- Guisinger, M.M.; Chumley, T.W.; Kuehl, J.V.; Boore, J.L.; Jansen, R.K. Implications of the plastid genome sequence of Typha (Typhaceae Poales) for understanding genome evolution in Poaceae. J. Mol. Evol. 2010, 70, 149–166. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krasnova, A.N.; Efremov, A.N. New species hydrophilic genus Typha L. (Typhaceae) of Central Yakutia. Nat. Resour. Arct. Subarct. Reg. 2018, 23, 120–125. [Google Scholar]
- Gouy, M.; Guindon, S.; Gascuel, O. SeaView version 4: A multiplatform graphical user interface for sequence alignment and phylogenetic tree building. Mol. Biol. Evol. 2010, 27, 221–224. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maddison, W.P.; Maddison, D.R. Mesquite: A Modular System for Evolutionary Analysis. Version 3.70. 2021. Available online: http://www.mesquiteproject.org (accessed on 11 November 2021).
- Edgar, R.C. MUSCLE: Multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Res. 2004, 32, 1792–1797. [Google Scholar] [CrossRef] [Green Version]
- Mavrodiev, E.V.; Madorsky, A. TAXODIUM Version 1.0: A simple way to generate uniform and fractionally weighted three-item matrices from various kinds of biological data. PLoS ONE 2012, 7, e48813. [Google Scholar] [CrossRef]
- Kitching, I.J.; Forey, P.; Forey, P.L.; Humphries, C.; Williams, D. Cladistics: The Theory and Practice of Parsimony Analysis; Oxford University Press: Oxford, UK, 1998. [Google Scholar]
- Nelson, G.; Platnick, N.I. Three-taxon statements—A more precise use of parsimony? Cladistics 1991, 7, 351–366. [Google Scholar] [CrossRef]
- Lapointe, F.J.; Cucumel, G. The average consensus procedure: Combination of weighted trees containing identical or overlapping sets of taxa. Syst. Biol. 1997, 46, 306–312. [Google Scholar] [CrossRef]
- Lapointe, F.J.; Levasseur, C. Everything you always wanted to know about the average consensus, and more. In Phylogenetic Supertrees: Combining Information to Reveal the Tree of Life; Bininda-Emonds, O.R.P., Ed.; Kluwer Academic Publisher: Dordrecht, Germany; Boston, MA, USA; London, UK, 2004; pp. 87–105. [Google Scholar]
- Mavrodiev, E.V.; Dell, C.; Schroder, L. A laid-back trip through the Hennigian Forests. PeerJ 2017, 5, e3578. [Google Scholar] [CrossRef] [Green Version]
- Mavrodiev, E.V.; Williams, D.M.; Ebach, M.C. On the Typology of Relations. Evol. Biol. 2019, 46, 71–89. [Google Scholar] [CrossRef]
- Swofford, D.L. PAUP*. Phylogenetic Analysis Using Parsimony (*And Other Methods); Sinauer Associates Inc.: Sunderland, MA, USA, 2002. [Google Scholar]
- Creevey, C.J.; McInerney, J.O. Trees from trees: Construction of phylogenetic supertrees using Clann. In Bioinformatics for DNA Sequence Analysis (Methods in Molecular Biology); Posada, D., Ed.; Springer: Berlin/Heidelberg, Germany; Humana Press: Totowa, NJ, USA, 2009; pp. 139–161. [Google Scholar]
- Muldashev, A.A. Typha shuttleworthii Koch and Sond. In Red List of the Republic of Bashkortostan, 237; Martynenko, V.B., Ed.; Studio Online: Moscow, Russia, 2021. [Google Scholar]
- Williams, D.M.; Ebach, M.C. Cladistics. A Guide to Biological Classification, 3rd ed.; Systematics Association Special, Volume Series 88; Cambridge University Press: Cambridge, UK, 2020. [Google Scholar]
- Shaw, J.; Lickey, E.B.; Schilling, E.E.; Small, R.L. Comparison of whole chloroplast genome sequences to choose noncoding regions for phylogenetic studies in angiosperms: The tortoise and the hare III. Am. J. Bot. 2007, 94, 275–288. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bilz, M.; Kell, S.; Maxted, N.; Lansdown, R. European Red List of Vascular Plants; Publication Office of the European Union: Luxembourg, 2011. [Google Scholar]
- Tilman, D.; May, R.M.; Lehman, C.L.; Nowak, M.A. Habitat destruction and the extinction debt. Nature 1994, 371, 65–66. [Google Scholar] [CrossRef]
- Chazdon, R.L.; Peres, C.A.; Dent, D.; Sheil, D.; Lugo, A.E.; Lamb, D.; Stork, N.E.; Miller, S.E. The potential for species conservation in tropical secondary forests. Conserv. Biol. 2009, 23, 1406–1417. [Google Scholar] [CrossRef] [PubMed]











Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kapitonova, O.A.; Muldashev, A.A.; Platunova, G.R.; Mavrodiev, E.V. Typha lepechinii Mavrodiev et Kapit. sp. nov. (Typhaceae Juss.)—A New Endangered Endemic Cattail in the Outmost East of European Russia. Taxonomy 2022, 2, 180-195. https://doi.org/10.3390/taxonomy2020014
Kapitonova OA, Muldashev AA, Platunova GR, Mavrodiev EV. Typha lepechinii Mavrodiev et Kapit. sp. nov. (Typhaceae Juss.)—A New Endangered Endemic Cattail in the Outmost East of European Russia. Taxonomy. 2022; 2(2):180-195. https://doi.org/10.3390/taxonomy2020014
Chicago/Turabian StyleKapitonova, Olga A., Albert A. Muldashev, Guzel R. Platunova, and Evgeny V. Mavrodiev. 2022. "Typha lepechinii Mavrodiev et Kapit. sp. nov. (Typhaceae Juss.)—A New Endangered Endemic Cattail in the Outmost East of European Russia" Taxonomy 2, no. 2: 180-195. https://doi.org/10.3390/taxonomy2020014