
Growth Characteristics, Phytochemical Contents, and Antioxidant Capacity of Trachyandra ciliata (L.f) Kunth Grown in Hydroponics under Varying Degrees of Salinity


Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Design
2.1.1. Plant Material
2.1.2. Nutrient Solutions
2.1.3. Medium Treatments
- Silica sand (100% sand),
- PPV (1:1:1—Peat: Perlite: vermiculite),
- Clay (100% Leca clay).
2.1.4. Salinity Treatments
2.2. Determination of Growth Parameters
2.2.1. Plant Weight
2.2.2. Shoot Length
2.2.3. Number of Leaves
2.2.4. Chlorophyll Content of Leaves
2.2.5. Formulation of Crude Extract
2.3. Determination of Phytochemical and Antioxidant Contents
2.3.1. Total Polyphenol Assay
2.3.2. Estimation of Flavonol Content
2.3.3. Determination of Ferric Reducing Antioxidant Power (FRAP)
2.3.4. Determination of ABTS Antioxidant Capacity
2.3.5. Antioxidant Capacity of DPPH Radicals
2.4. Statistical Analysis
3. Results
3.1. Effects of Salt Stress and Soilless Media on Vegetative Growth
3.1.1. The Effects of Salinity and Growth Media on Leaf Number
3.1.2. The Effects of Salinity and Growth Media on Plant Height
3.1.3. The Effects of Salinity and Growth Media on the Total Fresh Weight
3.1.4. The Effects of Salinity and Growth Media on the Total Dry Weight
3.1.5. The Effects of Salinity and Growth Media on Chlorophyll Content
3.1.6. The Effects of Salinity and Growth Media on the Number of Flowers
3.1.7. The Effects of Salinity and Growth Media on Inflorescence Fresh Weight
3.1.8. The Effects of Salinity and Growth Media on Inflorescence Dry Weight
3.2. Effects of Salt Stress on Phenolic Content and Antioxidant Capacity
3.2.1. Effect of Salinity and Soilless Media on the Accumulation of Polyphenols
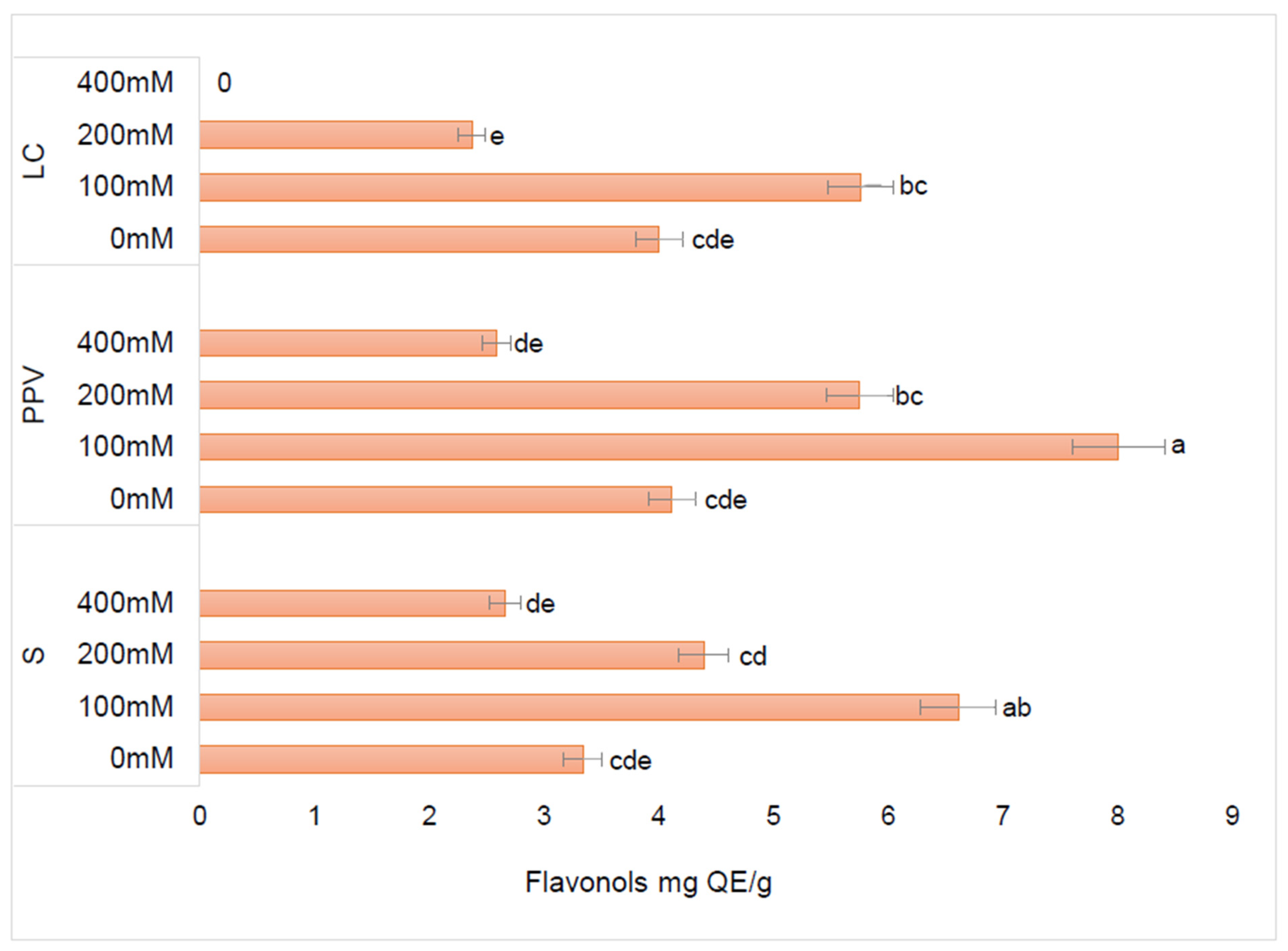
3.2.2. Effect of Different Salinity Levels and Soilless Media on Flavonols
3.2.3. The Effect of Salinity and Soilless Media on FRAP
3.2.4. Influence of Different Salinity Levels and Soilless Media on ABTS
3.2.5. Effect of Different Salinity Concentrations and Soilless Media on DPPH
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Abid, M.; Ali, S.; Qi, L.K.; Zahoor, R.; Tian, Z.; Jiang, D.; Snider, J.L.; Dai, T. Physiological and biochemical changes during drought and recovery periods at tillering and jointing stages in wheat (Triticum aestivum L.). Sci. Rep. 2018, 8, 1–15. [Google Scholar] [CrossRef]
- Bhusal, N.; Han, S.G.; Yoon, T.M. Impact of drought stress on photosynthetic response, leaf water potential, and stem sap flow in two cultivars of bi-leader apple trees (Malus × domestica Borkh.). Sci. Hortic. 2019, 246, 535–543. [Google Scholar] [CrossRef]
- Turton, A.; Mucheleng, C.; Mbao, E.; Musonda, W.; Mukubesa, L.; Ngoma, M. Water Demand Management, Natural Resource Reconstruction and Social Adaptive Capacity: A Case Study from Kolomo, Zambia. In Proceedings of the 2nd WARFSA/WaterNet Symposium Integrated Water Resource Management: Theory, Practice, Cases, Cape Town, South Africa, 30–31 October 2001; pp. 30–31. [Google Scholar]
- Singh, A. Conjunctive use of water resources for sustainable irrigated agriculture. J. Hydrol. 2014, 519, 1688–1697. [Google Scholar] [CrossRef]
- Gleick, P.H. Global Freshwater Resources: Soft-Path Solutions for the 21st Century. Science 2003, 302, 1524–1528. [Google Scholar] [CrossRef] [Green Version]
- Colvin, C.; Muruven, D. Scenarios for the Future of Water in South Africa; WWF South Africa: Cape Town, South Africa, 2017. [Google Scholar]
- Stefanov, M.; Yotsova, E.; Rashkov, G.; Ivanova, K.; Markovska, Y.; Apostolova, E.L. Effects of salinity on the photosynthetic apparatus of two Paulownia lines. Plant Physiol. Biochem. 2016, 101, 54–59. [Google Scholar] [CrossRef]
- Silveira, J.A.G.; Araújo, S.A.M.; Lima, J.P.M.S.; Viégas, R.A. Roots and leaves display contrasting osmotic adjustment mechanisms in response to NaCl-salinity in Atriplex nummularia. Environ. Exp. Bot. 2009, 66, 1–8. [Google Scholar] [CrossRef]
- Penella, C.; Landi, M.; Guidi, L.; Nebauer, S.G.; Pellegrini, E.; Bautista, A.S.; Remorini, D.; Nali, C.; López-Galarza, S.; Calatayud, A. Salt-tolerant rootstock increases yield of pepper under salinity through maintenance of photosynthetic performance and sinks strength. J. Plant Physiol. 2016, 193, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Daniels, C.W.; Rautenbach, F.; Mabusela, W.T.; Valentine, A.J.; Marnewick, J.L. Comparative antioxidant-capacity and -content of leaves, bulbs, roots, flowers and fruit of Gethyllis multifolia L. Bolus and G. villosa Thunb. species. S. Afr. J. Bot. 2011, 77, 711–717. [Google Scholar] [CrossRef]
- Ksouri, R.; Megdiche, W.; Debez, A.; Falleh, H.; Grignon, C.; Abdelly, C. Salinity effects on polyphenol content and antioxidant activities in leaves of the halophyte Cakile maritima. Plant Physiol. Biochem. 2007, 45, 244–249. [Google Scholar] [CrossRef]
- Jimoh, M.O.; Afolayan, A.J.; Lewu, F.B. Nutrients and antinutrient constituents of Amaranthus caudatus L. Cultivated on different soils. Saudi J. Biol. Sci. 2020, 27, 3570–3580. [Google Scholar] [CrossRef] [PubMed]
- Mazouz, W.; Haouli, N.E.H.; Gali, L.; Vezza, T.; Bensouici, C.; Mebrek, S.; Hamel, T.; Galvez, J.; Djeddi, S. Antioxidant, anti-alzheimer, anti-diabetic, and anti-inflammatory activities of the endemic halophyte Limonium spathulatum (Desf.) kuntze on LPS-stimulated RAW264 macrophages. S. Afr. J. Bot. 2020, 135, 101–108. [Google Scholar] [CrossRef]
- Mittler, R. ROS Are Good. Trends Plant Sci. 2017, 22, 11–19. [Google Scholar] [CrossRef] [Green Version]
- Randhir, R.; Lin, Y.-T.; Shetty, K. Phenolics, their antioxidant and antimicrobial activity in dark germinated fenugreek sprouts in response to peptide and phytochemical elicitors. Asian Pac. J. Clin. Nutr. 2004, 13, 295–307. [Google Scholar]
- Jimoh, M.O.; Afolayan, A.J.; Lewu, F.B. Antioxidant and phytochemical activities of Amaranthus caudatus L. harvested from different soils at various growth stages. Sci. Rep. 2019, 9, 12965. [Google Scholar] [CrossRef] [PubMed]
- Kambizi, L.; Bakare-Odunola, M.T.; Oladiji, A.T.; Kola-Mustapha, A.T.; Amusa, T.O.; Atolani, O.; Njinga, N.S.; Quadri, A.L. Proteinease inhibition, membrane stabilization, antioxidant and phytochemical evaluations of leaves, seeds and calyces of four selected edible medicinal plants. Cogent Chem. 2017, 3, 1–15. [Google Scholar] [CrossRef]
- Adegbaju, O.D.; Otunola, G.A.; Afolayan, A.J. Effects of growth stage and seasons on the phytochemical content and antioxidant activities of crude extracts of Celosia argentea L. Heliyon 2020, 6, e04086. [Google Scholar] [CrossRef]
- Salami, S.O.; Afolayan, A.J. Evaluation of nutritional and elemental compositions of green and red cultivars of roselle: Hibiscus sabdariffa L. Sci. Rep. 2021, 11, 1–13. [Google Scholar] [CrossRef]
- Jimoh, M.A.; Idris, O.A.; Jimoh, M.O. Cytotoxicity, Phytochemical, Antiparasitic Screening, and Antioxidant Activities of Mucuna pruriens (Fabaceae). Plants 2020, 9, 1249. [Google Scholar] [CrossRef]
- Finley, J.W. Phenolic antioxidants and prevention of chronic inflammation. Food Technol. 2004, 58, 42–46. [Google Scholar]
- Idris, O.A.; Wintola, O.A.; Afolayan, A.J. Comparison of the proximate composition, Vitamins (Ascorbic acid, α-Tocopherol and retinol), anti-nutrients (phytate and oxalate) and the GC-MS analysis of the essential oil of the root and leaf of Rumex Crispus L. Plants 2019, 8, 51. [Google Scholar] [CrossRef] [Green Version]
- Ventura, Y.; Wuddineh, W.A.; Myrzabayeva, M.; Alikulov, Z.; Khozin-goldberg, I.; Shpigel, M.; Samocha, T.M.; Sagi, M. Effect of seawater concentration on the productivity and nutritional value of annual Salicornia and perennial Sarcocornia halophytes as leafy vegetable crops. Sci. Hortic. 2011, 128, 189–196. [Google Scholar] [CrossRef]
- Klados, E.; Tzortzakis, N. Effects of substrate and salinity in hydroponically grown Cichorium spinosum. J. Soil Sci. Plant Nutr. 2014, 14, 211–222. [Google Scholar] [CrossRef] [Green Version]
- Botía, P.; Carvajal, M.; Cerdá, A.; Martínez, V. Response of eight Cucumis melo cultivars to salinity during germination and early vegetative growth. Agronomie 1998, 18, 503–513. [Google Scholar] [CrossRef] [Green Version]
- Ortiz, A.; Rotatori, H.; Schreiber, L.; von Roth, G. Hydroponic Farming in Mahasarakham: Integrating Hydroponics into the Agricultural Curriculum while Promoting Entrepreneurial Skills; Worcester Polytechnic Institute: Worcester, MA, USA, 2009. [Google Scholar]
- Wahome, P.K.; Oseni, T.; Masarirambi, M. Effects of different hydroponics systems and growing media on the vegetative growth, yield and cut flower quality of gypsophila (Gypsophila paniculata L.). J. Agric. Sci. 2011, 7, 692–698. [Google Scholar]
- Manning, J.C.; Goldblatt, P. Trachyandra arenicola and Trachyandra montana, two new species from South Africa. Bothalia 2007, 37, 27–30. [Google Scholar]
- Van Jaarsveld, E. Trachyandra Asphodelaceae. In Monocotyledons; Springer: Berlin/Heidelberg, Germany, 2020; pp. 829–832. [Google Scholar]
- Van Wyk, B.-E. The potential of South African plants in the development of new medicinal products. S. Afr. J. Bot. 2011, 77, 812–829. [Google Scholar] [CrossRef] [Green Version]
- De Vynck, J.C.; Van Wyk, B.E.; Cowling, R.M. Indigenous edible plant use by contemporary Khoe-San descendants of South Africa’s Cape South Coast. S. Afr. J. Bot. 2016, 102, 60–69. [Google Scholar] [CrossRef]
- Faber, R.J.; Laubscher, C.P.; Rautenbach, F.; Jimoh, M.O. Variabilities in alkaloid concentration of Sceletium tortuosum (L.) N.E. Br in response to different soilless growing media and fertigation regimes in hydroponics. Heliyon 2020, 6, e05479. [Google Scholar] [CrossRef] [PubMed]
- Sogoni, A.; Jimoh, M.; Kambizi, L.; Laubscher, C. The Impact of Salt Stress on Plant Growth, Mineral Composition, and Antioxidant Activity in Tetragonia decumbens Mill.: An Underutilized Edible Halophyte in South Africa. Horticulturae 2021, 7, 140. [Google Scholar] [CrossRef]
- Nkcukankcuka, M.; Jimoh, M.O.; Griesel, G.; Laubscher, C.P. Growth characteristics, chlorophyll content and nutrients uptake in Tetragonia decumbens Mill. cultivated under different fertigation regimes in hydroponics. Crop Pasture Sci. 2021, 72, 1–12. [Google Scholar] [CrossRef]
- Ainsworth, E.A.; Gillespie, K.M. Estimation of total phenolic content and other oxidation substrates in plant tissues using Folin-Ciocalteu reagent. Nat. Protoc. 2007, 2, 875–877. [Google Scholar] [CrossRef]
- Singleton, V.L.; Orthofer, R.; Lamuela-Raventós, R.M. Analysis of total phenols and other oxidation substrates and antioxidants by means of folin-ciocalteu reagent. Methods Enzymol. 1999, 299, 152–178. [Google Scholar] [CrossRef]
- Mazza, G.; Fukumoto, L.; Delaquis, P.; Girard, B.; Ewert, B. Anthocyanins, phenolics, and color of Cabernet Franc, Merlot, and Pinot Noir wines from British Columbia. J. Agric. Food Chem. 1999, 47, 4009–4017. [Google Scholar] [CrossRef]
- Benzie, I.F.F.; Strain, J.J. Ferric reducing/antioxidant power assay: Direct measure of total antioxidant activity of biological fluids and modified version for simultaneous measurement of total antioxidant power and ascorbic acid concentration. Methods Enzymol. 1999, 299, 15–27. [Google Scholar] [CrossRef] [PubMed]
- Idris, O.A.; Wintola, O.A.; Afolayan, A.J. Phytochemical and antioxidant activities of Rumex crispus L. in treatment of gastrointestinal helminths in Eastern Cape Province, South Africa. Asian Pac. J. Trop. Biomed. 2017, 7, 1071–1078. [Google Scholar] [CrossRef]
- Unuofin, J.O.; Otunola, G.A.; Afolayan, A.J. Nutritional evaluation of Kedrostis africana (L.) Cogn: An edible wild plant of South Africa. Asian Pac. J. Trop. Biomed. 2017, 7, 443–449. [Google Scholar] [CrossRef]
- Koryo, H. Effect of salinity on growth, photosynthesis and solute composition of the potential cash crop halophyte plantago. Environ. Exp. Bot. 2006, 56, 136–146. [Google Scholar] [CrossRef]
- Rengasamy, P.; Chittleborough, D.; Helyar, K. Root-zone constraints and plant-based solutions for dryland salinity. Plant Soil 2003, 257, 249–260. [Google Scholar] [CrossRef]
- Hussain, A.; Iqbal, K.; Aziem, S.; Mahato, P.; Negi, A.K. A Review on the Science of Growing Crops Without Soil (Soilless Culture)—A Novel Alternative For Growing Crops. Int. J. Agric. Crop Sci. 2014, 7, 833–842. [Google Scholar]
- Adams, P. Effects of increasing the salinity of the nutrient solution with major nutrients or sodium chloride on the yield, quality and composition of tomatoes grown in rockwool. J. Hortic. Sci. 1991, 66, 201–207. [Google Scholar] [CrossRef]
- Sayyad-Amin, P.; Jahansooz, M.R.; Borzouei, A.; Ajili, F. Changes in photosynthetic pigments and chlorophyll-a fluorescence attributes of sweet-forage and grain sorghum cultivars under salt stress. J. Biol. Phys. 2016, 42, 601–620. [Google Scholar] [CrossRef] [Green Version]
- Khan, M.A.; Ungar, I.A.; Showalter, A.M. The effect of salinity on the growth, water status, and ion content of a leaf succulent perennial halophyte, Suaeda fruticosa (L.) Forssk. J. Arid Environ. 2000, 45, 73–84. [Google Scholar] [CrossRef] [Green Version]
- Guo, J.; Li, Y.; Han, G.; Song, J.; Wang, B. NaCl markedly improved the reproductive capacity of the euhalophyte Suaeda salsa. Funct. Plant Biol. 2018, 45, 350–361. [Google Scholar] [CrossRef]
- Shalhevet, J.; Heuer, B.; Meiri, A. Irrigation interval as a factor in the salt tolerance of eggplant. Irrig. Sci. 1983, 4, 83–93. [Google Scholar] [CrossRef]
- Gama, P.B.S.; Inanaga, S.; Tanaka, K.; Nakazawa, R. Physiological response of common bean (Phaseolus vulgaris L.) seedlings to salinity stress. Afr. J. Biotechnol. 2007, 6, 079–088. [Google Scholar] [CrossRef]
- Yurtseven, E.; Kesmez, G.D.; Ünlükara, A. The effects of water salinity and potassium levels on yield, fruit quality and water consumption of a native central anatolian tomato species (Lycopersicon esculantum). Agric. Water Manag. 2005, 78, 128–135. [Google Scholar] [CrossRef]
- Lyons, A.; Kass-Hanna, J.; Greenlee, A. Impacts of Financial and Digital Inclusion on Poverty in South Asia and Sub-Saharan Africa. SSRN Electron. J. 2020, 1, 0–39. [Google Scholar] [CrossRef]
- Venter, G. Successful Hydroponics: 21st Century Technology for Commercial and Home Applications: A Comprehensive Practical Guide to Scientifically Based Hydroponic Crop; Xlibris Corporation: Dartford, UK, 2010. [Google Scholar]
- Zapryanova, N.; Atanassova, B. Effects of salt stress on growth and flowering of ornamental annual species. Biotechnol. Biotechnol. Equip. 2009, 23, 177–179. [Google Scholar] [CrossRef] [Green Version]
- Ventura, Y.; Myrzabayeva, M.; Alikulov, Z.; Omarov, R.; Khozin-Goldberg, I.; Sagi, M. Effects of salinity on flowering, morphology, biomass accumulation and leaf metabolites in an edible halophyte. AoB Plants 2014, 6, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Stanton, M.L.; Galen, C.; Shore, J. Population structure along a steep environmental gradient: Consequences of flowering time and habitat variation in the snow buttercup, Ranunculus adoneus. Evolution 1997, 51, 79–94. [Google Scholar] [CrossRef]
- Pardossi, A.; Carmassi, G.; Diara, C.; Incrocci, L.; Maggini, R.; Massa, D. Fertigation and Substrate Management in Closed Soilless Culture; University of Pisa: Pisa, Italy, 2011. [Google Scholar]
- Rezazadeh, A.A.; Ghasemnezhad, M.B.; Telmadarrehei, T. Effect of salinity on phenolic composition and antioxidant activity of artichoke (Cynara scolymus L.) leaves. Res. J. Med. Plants 2012, 6, 245–252. [Google Scholar] [CrossRef] [Green Version]
- Ben-Abdallah, S.; Zorrig, W.; Amyot, L.; Renaud, J.; Hannoufa, A.; Lachâal, M.; Karray-Bouraoui, N. Potential production of polyphenols, carotenoids and glycoalkaloids in Solanum villosum Mill. under salt stress. Biologia 2019, 74, 309–324. [Google Scholar] [CrossRef]
- Parvez, S.; Abbas, G.; Shahid, M.; Amjad, M.; Hussain, M.; Asad, S.A.; Imran, M.; Naeem, M.A. Effect of salinity on physiological, biochemical and photostabilizing attributes of two genotypes of quinoa (Chenopodium quinoa Willd.) exposed to arsenic stress. Ecotoxicol. Environ. Saf. 2020, 187, 109814. [Google Scholar] [CrossRef]
- Shabala, S.; Pottosin, I. Regulation of potassium transport in plants under hostile conditions: Implications for abiotic and biotic stress tolerance. Physiol. Plant. 2014, 151, 257–279. [Google Scholar] [CrossRef]
- Ben Abdallah, S.; Aung, B.; Amyot, L.; Lalin, I.; Lachâal, M.; Karray-Bouraoui, N.; Hannoufa, A. Salt stress (NaCl) affects plant growth and branch pathways of carotenoid and flavonoid biosyntheses in Solanum nigrum. Acta Physiol. Plant. 2016, 38, 1–13. [Google Scholar] [CrossRef]
- Wong, C.C.; Li, H.B.; Cheng, K.W.; Chen, F. A systematic survey of antioxidant activity of 30 Chinese medicinal plants using the ferric reducing antioxidant power assay. Food Chem. 2006, 97, 705–711. [Google Scholar] [CrossRef]
- Machado, R.M.A.; Serralheiro, R.P. Soil salinity: Effect on vegetable crop growth. Management practices to prevent and mitigate soil salinization. Horticulturae 2017, 3, 30. [Google Scholar] [CrossRef]
- Sharma, V.; Ramawat, K.G. Salinity-induced modulation of growth and antioxidant activity in the callus cultures of miswak (Salvadora persica). 3 Biotech 2013, 3, 11–17. [Google Scholar] [CrossRef] [Green Version]
- Sharma, V.; Ramawat, K.G. Salt stress enhanced antioxidant response in callus of three halophytes (Salsola baryosma, Trianthema triquetra, Zygophyllum simplex) of Thar Desert. Biologia 2014, 69, 178–185. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Soil-Less Medium | NaCl Conc. (mM) | Total Dry Weight (g) | Total Fresh Weight (g) | Leaf No. | Shoot Length (cm) |
|---|---|---|---|---|---|
| Silica Sand | 0 | 19.62 ± 2.56 a | 131.02 ± 20.2 bc | 37.90 ± 3.78 a | 73.50 ± 1.61 e |
| 100 | 12.24 ± 1.79 b | 136.40 ± 21.44 bc | 34.10 ± 5.7 a | 68.70 ± 8.72 e | |
| 200 | 1.97 ± 0.76 c | 22.15 ± 7.54 d | 7.30 ± 2.1 b | 31.00 ± 8.66 ab | |
| 400 | 0.3 ± 0.19 c | 2.15 ± 1.13 d | 2.00 ± 1.04 b | 5.70 ± 3.02 cd | |
| PPV | 0 | 15.92 ± 2.08 ab | 142.16 ± 18.47 ac | 34.30 ± 6.71 a | 77.50 ± 8.81 e |
| 100 | 13.72 ± 0.98 b | 156.31 ± 14.16 ac | 42.80 ± 3.89 a | 72.90 ± 3.34 e | |
| 200 | 3.62 ± 1.11 c | 34.91 ± 11.22 d | 9.60 ± 2.66 b | 34.80 ± 9.67 a | |
| 400 | 0.78 ± 0.3 c | 6.88 ± 2.81 d | 4.10 ± 1.64 b | 14.60 ± 5.04 bcd | |
| LECA Clay | 0 | 14.66 ± 1.58 b | 99.68 ± 13.31 b | 35.70 ± 3.01 a | 67.00 ± 1.91 e |
| 100 | 15.40 ± 2.53 b | 183.11 ± 29.95 a | 36.30 ± 6.59 a | 68.00 ± 8 e | |
| 200 | 1.26 ± 0.44 c | 11.17 ± 3.76 d | 5.20 ± 1.81 b | 19.70 ± 6.62 abc | |
| 400 | 0.00 ± 0 c | 0.00 ± 0 d | 0.00 ± 0 b | 0.00 ± 0 d | |
| Two-way ANOVA F-Statistics | |||||
| Soil-less Medium | 0.30 ns | 0.84 ns | 0.82 ns | 3.17 * | |
| NaCl Conc. | 92.02 * | 77.24 * | 70.74 * | 77.62 * | |
| Soil-less Medium * NaCl Conc. | 1.55 ns | 0.19 ns | 0.46 ns | 0.20 ns | |
| Soil-Less Medium | NaCl Conc. (mM) | Number of Flowers | Inflorescence Fresh Weight (g) | Inflorescence Dry Weight (g) | Chlorophyll |
|---|---|---|---|---|---|
| Silica sand | 0 | 1.20 ± 0.42 b | 33.2 ± 13.17 ab | 2.08 ± 0.99 bc | 283.40 ± 9.89 ab |
| 100 | 0.30 ± 0.15 c | 7.07 ± 4.02 c | 0.74 ± 0.41 cd | 257.30 ± 29.7 ab | |
| 200 | 0.00 ± 0 c | 0.00 ± 0 c | 0.00 ± 0 d | 166.60 ± 46.42 bcd | |
| 400 | 0.00 ± 0 c | 0.00 ± 0 c | 0.00 ± 0 d | 69.00 ± 35.9 de | |
| PPV | 0 | 0.90 ± 0.28 b | 40.14 ± 13.19 ab | 3.47 ± 1.15 b | 270.60 ± 33.5 ab |
| 100 | 1.80 ± 0.25 a | 46.73 ± 7.9 a | 5.14 ± 0.79 a | 278.30 ± 7.91 ab | |
| 200 | 0.00 ± 0 c | 0.00 ± 0 c | 0.00 ± 0 d | 141.00 ± 39.52 bcde | |
| 400 | 0.00 ± 0 c | 0.00 ± 0 c | 0.00 ± 0 d | 103.70 ± 36.64 de | |
| LECA Clay | 0 | 0.90 ± 0.23 b | 27.77 ± 6.86 b | 2.67 ± 0.67 b | 324.00 ± 16.14 a |
| 100 | 0.30 ± 0.21 c | 7.20 ± 6.27 c | 0.60 ± 0.52 cd | 254.20 ± 28.91 abc | |
| 200 | 0.00 ± 0 c | 0.00 ± 0 c | 0.00 ± 0 d | 111.30 ± 38.18 cde | |
| 400 | 0.00 ± 0 c | 0.00 ± 0 c | 0.00 ± 0 d | 0.00 ± 0 f | |
| Two-way ANOVA F-Statistics | |||||
| Soil-less Medium | 4.34 ns | 4.78 ns | 8.15 * | 0.84 ns | |
| NaCl Conc. | 22.87 * | 19.2 * | 19.38 * | 38.99 * | |
| Soil-less Medium * NaCl Conc. | 5.72 * | 2.79 ns | 4.78 * | 1.35 ns | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ngxabi, S.; Jimoh, M.O.; Kambizi, L.; Laubscher, C.P. Growth Characteristics, Phytochemical Contents, and Antioxidant Capacity of Trachyandra ciliata (L.f) Kunth Grown in Hydroponics under Varying Degrees of Salinity. Horticulturae 2021, 7, 244. https://doi.org/10.3390/horticulturae7080244
Ngxabi S, Jimoh MO, Kambizi L, Laubscher CP. Growth Characteristics, Phytochemical Contents, and Antioxidant Capacity of Trachyandra ciliata (L.f) Kunth Grown in Hydroponics under Varying Degrees of Salinity. Horticulturae. 2021; 7(8):244. https://doi.org/10.3390/horticulturae7080244
Chicago/Turabian StyleNgxabi, Sihle, Muhali Olaide Jimoh, Learnmore Kambizi, and Charles Petrus Laubscher. 2021. "Growth Characteristics, Phytochemical Contents, and Antioxidant Capacity of Trachyandra ciliata (L.f) Kunth Grown in Hydroponics under Varying Degrees of Salinity" Horticulturae 7, no. 8: 244. https://doi.org/10.3390/horticulturae7080244