
Evolution of the Secondary Metabolites in Invasive Plant Species Chromolaena odorata for the Defense and Allelopathic Functions

Department of Applied Biological Science, Faculty of Agriculture, Kagawa University, Miki, Kagawa 761-0795, Japan
*
Author to whom correspondence should be addressed.
Plants 2023, 12(3), 521; https://doi.org/10.3390/plants12030521
Submission received: 5 January 2023
/
Revised: 16 January 2023
/
Accepted: 20 January 2023
/
Published: 23 January 2023
(This article belongs to the Special Issue Plant Invasion 2022)
Abstract
:Chromolaena odorata (L.) R.M. King & H. Robinson is native to tropical America, and has naturalized in many other countries in tropical Asia, Austria, and West Africa. The species often forms dense thickets and reduces the native species diversity and population in the invasive ranges. The species is also considered as a noxious weed in agriculture fields, and listed in the 100 of the world’s worst invasive alien species. The characteristics of its life-history such as the seed production rate, growth pattern, and adaptative ability to the environmental conditions may contribute to the invasiveness of the species. Possible evidence of the defense capacity against the natural enemy, and the allelopathic potential against the competitive plant species for C. odorata has been accumulated in the literature over three decades. The extracts, residues, and/or rhizosphere soil of C. odorata increased the mortality of various insects and parasitic nematodes, and decreased their population. The extracts, residues, and/or rhizosphere soil of C. odorata also inhibited the germination and growth of several plant species including the indigenous plant species in the invasive ranges of C. odorata. Toxic substances, pyrrolizidine alkaloids were found in the leaves and flowers of C. odorata. These pyrrolizidine alkaloids may work as the defense agents against the natural enemies. Several potential allelochemicals such as flavonoids, phenolic acids, and terpenoids were also found in the plant extracts of C. odorata. Some of these compounds may work as allelopathic agents of C. odorata and inhibit the germination and growth of the competitive plant species. These characteristics of C. odorata for the defense function against their natural enemies such as insects and parasitic nematodes, and allelopathic potential against the competitive native plant species may contribute to the invasiveness and naturalization of C. odorata in the new habitats as invasive plant species. However, it is necessary to determine the concentration of these allelochemicals in the neighboring environment of C. odorata such as the rhizosphere soil since allelochemicals are able to work only when they are released into the neighboring environment. It is the first review article focusing on the defense function and allelopathy of C. odorata.
Keywords:
allelochemical; herbivore; invasive species; monospecific stand; natural enemy; nematode; pathogen1. Introduction
Chromolaena odorata (L.) R.M. King & H. Robinson (synonym, Eupatorium odoratum L.), belonging to Asteraceae, is a perennial shrub, and grows to 2–3 m in height, but it can scramble up other plants and reach 5–10 m in length. Its stems are cylindrical and pithy, become woody, and often branch in pairs from the axillary buds. The opposite leaves are ovate-triangular, 6–12 cm long and 3–7 cm wide with a 1–3 cm petiole, and smell a strong odor when they are crushed [1] (Figure 1). It has an abundantly branched lateral fine root system supported by corms which are underground swollen stems and storage nutrients [1,2,3]. Capitula are generated in panicles at the end of the twigs, and a single capitulum contains 15–35 florets. The corollas of the florets vary in color ranging from white to pale-lilac. The species grows on all types of well-drained soil, and where the temperature ranges 20–37 °C, and the minimum annual rainfall is 1500 mm [4,5]. However, it grows best in sunny and open areas such as roadsides, riverbanks, and vacant land [6,7,8].
The native range of C. odorata is from southern Florida and Texas (30° N) to north-western Argentina (30° S) including the Caribbean islands [9,10,11,12,13]. The species is thought to be introduced into Asia as an ornamental plant in 1840s through the Botanical garden in Kolkata, India. The first record of naturalization of C. odorata was in the 1870s in Indo-Gangetic Plain [1]. The species has also been introduced, spread rapidly, and naturalized in many other countries in eastern and southern Asia, Austria, and southern and western Africa [1,11,12].
C. odorata infests in a wide range of natural vegetation such as grassland, savanna, bush, forest margins and gaps, dry deciduous and degraded forests. The species scrambles up to the canopy of trees, and spreads over the trees, eventually reducing the vigor of the trees [14,15,16]. After infestation, it grows rapidly and often forms a dense thicket, extending its multiple branches and twisting them around the existing vegetation. The thickets of the species prevent the movement of the wildlife and livestock, which affects their breeding and population [1,17,18]. It was reported that the species reduced the biodiversity and population in its introduced vegetation [19,20]. Average number of the plant species in the infested areas of C. odorata was reduced by 31% [21]. C. odorata also infests as a weed in agricultural lands such as oil palm, rubber, cacao, coffee, coconut and banana plantations, pastures, crop fields, and abandoned agriculture fields. The species is considered as a noxious weed in agricultural fields and commercial plantations [22,23], and listed in the 100 of the world’s worst invasive alien species [11,12]. In its native ranges, C. odorata is only weedy on some occasions such as after the fire, hurricane, human activity, and other destruction events. It is then forced out by the successional vegetation and disappears [24,25]. However, the species forms dense monospecific stands and shrives over 15 years in the introduced ranges [1,26].
It was estimated that approximately 10% of the introduced plant species could be established in the introduced ranges, and 10% of the established plants become invasive [27], which indicates only 1% of the introduced plants could be invasive. The characteristics of life-history, such as the high reproduction rate, phenotypic plasticity, and competitive ability of the plants are important for the naturalization of invasive plants into the introduced ranges [28,29,30,31].
C. odorata is also a prolific seed producer. It was reported to produce 2000 (1 year old C. odorata)–260,000 (10 year old) seeds per m2 with 20–46% of seeds being viable. The germination rate of the seeds collected from the seed banks was 5–20% [26,32,33]. The foliage of the species, which contains essential oil, is flammable and increases the wild fire occurrence and its maximum temperature [34,35]. The intense fire increases the mortality of indigenous herbaceous plant species as well as the juvenile forest woody plants. The development of the stump sprouts of C. odorata from its underground stems, corms, arises quickly after the fire, and the regeneration of the species occurs [2,36,37]. Therefore, the wild fire may convert a native ecosystem of the woody plants and herbaceous into an C. odorata dominated ecosystem.
Genetic diversity of C. odorata in the introduced ranges was lower than that in the native ranges [38,39,40,41]. The morphological variability of C. odorata such as flower color, leaf shape, and plant shape is high in the native range, while there are only two main biotypes, Asia/West Africa biotype and South Africa biotype, in the invasive ranges [1]. Asia/West Africa biotype is thought to be originating from Trinidad and Tobago [40,42], while South Africa biotype is thought to be originating from Jamaica and/or Cuba [42,43]. However, the adaptative potential of the species was higher in the invasive range than that of the native ranges in the response to the environmental factors such as light conditions, mean annual temperature and precipitation [39,44,45]. Drought tolerance of the species in the invasive ranges was also higher than that in the native ranges [44,45]. It was reported that total biomass and plant height of C. odorata obtained from invasive ranges were larger than those from the native ranges [41]. On the contrary, the aboveground biomass and plant height of C. odorata in the invasive ranges were smaller than those of the native ranges [39]. The aboveground biomass of C. odorata in the native ranges was 30% greater than that in the invasive ranges [46].
Plants produce large number of secondary metabolites in many chemical classes. The biosynthesis of some secondary metabolites is induced or synthesized de-novo under certain circumstances. Many of these secondary metabolites in the invasive plants have been reported to show multiple functions such as anti-herbivore, anti-fungal, anti-microbial, and allelopathic activity, and contribute to increasing the fitness of the plants in the invasive ranges [47,48,49,50,51,52,53,54,55,56]. Available information from a large number of publications suggests that C. odorata is allelopathic, and contains the compounds involved in the allelopathy. The plant species was also reported to show the defense response against the natural enemies such as herbivores and pathogens, and contain the compounds involved in the defense function. However, there has been no review paper focusing on the allelopathy and defense response of C. odorata, and compounds involved in these functions. This review provides an overview of the defense response and allelopathy of the species and compounds involved in the defense and allelopathy. Then, their possible involvement in the invasiveness of the species is discussed.
2. Interaction of C. odorata with the Natural Enemy
The interaction between the invasive plants and their natural enemies such as herbivores and pathogens, is one of the important factors for the naturalization of the invasive plants [29,30,31,57,58]. The population of C. odorata is controlled by many insects and pathogens in its native ranges [59,60]. More than 200 species of the herbivores were counted in the native ranges of C. odorata, and 25% of them are specific species in the native ranges [61]. There may be fewer specific herbivores in the invasive ranges. In fact, very few specific insect species for C. odorata in the invasive range (South Africa) were counted [62]. According to the evolution of increased competitive ability hypothesis, the success of the invasive species is due to fewer specialized predators in the invasive ranges. The invasive plants can allocate the resources from the high-cost defense strategy to the low-cost defense strategy and plant growth, leading to the successful naturalization [63,64].
2.1. Interaction of C. odorata with Insects
Powder of the roots, stems, and leaves of C. odorata increased the mortality of leaf beetle, Callosobruchus maculatus Fabricius [65,66]. Essential oil obtained from C. odorata leaves also increased the mortality of adult weevil, Sitophilus zeamais Motschulsky [67,68]. Aqueous leaf extract of C. odorata induced the increasing larval mortality of black fly, Simulium spp. [69], and an adult stage of cockroach, Periplaneta americana Linnaeus [70]. Aqueous ethanol leaf extracts of C. odorata were applied to Aabelmoschus esculentus (L.) Moench once a week for 4–7 weeks after its planning. The treatments resulted in the reduction of the population of whitefly, Bemissa tabaci Gennadius, and leafhopper, Amrasca biguttula Ishida on Aabelmoschus esculentus [71]. The methanol extracts of C. odorata leaves also showed ovicidal, antifeedant, and larvicidal activity on a leaf-eating insect, Spodoptera litura Fabricius [72]. These observations suggest that C. odorata possess anti-insect activity and contain certain compounds involved in the activity.
Pyrrolizidine alkaloids such as 7- and 9-angeloylretronecine, intermedine, rinderine, and 3′-acetylrinderine were isolated from roots and mature flower heads of C. odorata [73], and rinderine N-oxide and intermedine N-oxide were identified in its roots [74] (Figure 2). Pyrrolizidine alkaloids are amino alcohols, esterified with mono- or dicarboxylic acids [75], and act as chemical defense agents against herbivores such as insects and mammals [76,77,78,79]. The compounds are highly toxic including hepatotoxicity, and disturb several metabolisms in the cell functions [80,81,82,83]. Therefore, these pyrrolizidine alkaloids in C. odorata may be involved in the anti-insect activity caused by the extracts and power of the species as describe above, and contribute to the protection of the species from herbivore attacks.
Some specialist insects obtain pyrrolizidine alkaloids from plants and store them. Those stored pyrrolizidine alkaloids are used for their protection from their predators as poison, and for the precursors to synthesize their mail pheromones [75,84,85]. However, C. odorata may scarcely meet these specialist insects in its invasive ranges since the host plants of the sspecialist insects are narrow and there may be no such co-evolutional history between C. odorata and the insects in the invasive ranges (Figure 2, Table 1).
2.2. Interaction of C. odorata with Nematodes
Plant parasitic nematodes such as Meloidogyne spp. (root-knot nematode), Helicotylenchus spp., and Pratylenchus spp. feed on the roots of plants, and their feeding process causes serious injuries and reduces the ability of the plants to absorb nutrients and water, leading to losing plant vigor and defense capability against other pathogen attacks [86,87,88]. The population density of nematodes such as Meloidogyne spp., Helicotylenchus spp. and Pratylenchus spp., was suppressed by the C. odorata infestation into the invasive ranges with various soil conditions [89]. It was also reported that C. dorata reduced by 77–81% of plant parasitic nematode population; Meloidogyne spp., Helicotylenchus spp., and Pratylenchus spp. in the soils after two years invention [90]. C. odorata also suppressed the increasing population of Meloidogyne incoginita Kofoid & White in the pot experiments [91]. These observations suggest that some compounds may be released from C. odorata and accumulated in the soils, and these compounds may suppress the population of plant parasitic nematodes in the soils.
The incorporation of plant powder of C. odorata into the field soil prevented the increasing population of Meloidogyne incoginita [92]. Aqueous root extracts and root mulch of C. odorata showed the suppression of the parasitism of Meloidogyne incoginita into the roots of Lactuca sativa L. [93]. Therefore, certain compounds in the plant powder, roots, and extracts of C. odorata may work for the suppression.
1,2-Dehydropyrrolizine alkaloid was identified in the root extracts of C. odorata and the compound showed anti-nematode activity [93] (Figure 2). 1,2-Dehydropyrrolizine alkaloid was reported to be synthesized and stored in vacuole in the roots of C. odorata [94]. As described in the Section 2.1, pyrrolizidine alkaloids are highly toxic and act as chemical defense agents against natural enemies [76,77,78]. Those observations suggest that the extracts and powder of C. odorata, and the soil under C. odorata may suppress the population of the nematodes, and prevented the hatch and parasitism of the nematodes. Certain compounds including 1,2-dehydropyrrolizine alkaloid may cause the suppression and acts as anti-nematode agents of C. odorata (Table 2).
2.3. Interaction of C. odorata with Microbial
The invasion of C. odorata into the forest and savanna in West Africa increased the soil microbial activity, and the amount of available N and P in the soil. C. odorata altered the soil microbial community in the invasion ranges. The altered microbial community suppressed the growth of the native plant species such as Eupatorium japonicum Thunb. and Eupatorium heterophyllum DC., and stimulated the growth of C. odorata [95]. The population of an arbuscular mycorrhizal fungus Paraglomus spp. was also increased in the soil under C. odorata [96]. Arbuscular mycorrhizal fungi increase the ability of their host plants to absorb water and nutrients, and enhance the defense function against several stress conditions and pathogen attacks [97,98,99]. The observations suggest that certain compounds from C. odorata may alter the soil microbial community in the invasion ranges and the alteration may contribute the invasion of C. odorata. However, an arbuscular mycorrhizal fungus Paraglomus spp. colonizes with a wide range of plant species [100,101]. The colonization may occur with other plant species and promote their growth in the invasive ranges of C. odorata. In addition, the abundance of the arbuscular mycorrhizal colonization of C. odorata in its invasive range (South Africa) was reported to be 50% of its native ranges (Puerto Rico) [102].
The rhizosphere soil of C. odorata increased the population of the soil borne fungal pathogen, Fusarium ssp., and inhibited the growth of Amaranthus spinosus L. and Bambusa bambos (L.) Voss. Sterilization of the soil eliminated these effects. The root leachate of C. odorata increased the spore density of Fusarium spp. in C. odorata-free soil. The increases were illuminated by adding activated carbon into the soil [103]. It was also reported that the root exudates of Sorghum bicolor (L.) Moench. and Vigna unguiculata (L.) Walp. increased the population of Fusarium spp. [104], and phenolics in the root exudates of Glycine max (L.) Merr. increased the population of Fusarium spp. [105]. The observations suggest that certain compounds in the root exudate of these plant species including C. odorata may stimulate the increasing population of Fusarium spp. However, it is not clear if the increased Fusarium spp. population affects the growth of C. odorata.
On the contrary, it was reported that the extracts of C. odorata suppressed the growth of some soil borne fungal pathogens including Fusarium spp. Aqueous methanol leaf extracts of C. odorata significantly suppressed the colony growth of the pathogens, Lasiodiplodia theobromae (Pat.) Griffon & Maubl. and Lasiodiplodia pseudothobromae A.J.L. Phillips, A. Alves & Crous [106]. Both Lasiodiplodia spp. are members of the Botryosphaeriaceae family and cause leaf necrosis, canker, and dieback in many plant species [107,108]. The methanol leaf extracts of C. odorata suppressed the growth of Bacillus subtilis Cohn, and Bacillus cereus Frankland & Frankland [109]. The ethanol plant extracts of C. odorata suppressed the growth of soil borne pathogen fungi, Phytophthora colocasiae Racib., and Fusarium oxysporum Schlecht. emend. Snyder & Hansen [110]. Acetone extracts of C. odorata also suppressed the colony growth of the pathogen fungi, Pythium ultimum Trow, Rhizoctonia solani J.G. Kühn, Fusarium oxysporium Schlecht. emend. Snyder & Hansen, and Phytophthora nicotianae Breda de Haan [111] and Pyricularia oryzae Cavara [112]. Essential oil of C. odorata suppressed the growth of Rhizoctonia solani J.G. Kühn, Fusarium graminearum Schwabe, Exserohilum turcicum (Pass.) K.J. Leonard & Suggs, Botrytis cinereal Pers., and Sclerotinia sclerotiorum (Lib.) de Bary [113]. These observations suggest that the extracts of C. odorata possess the anti-fungal activity, and may contain certain compounds involved in the activity.
Some compounds in the extracts and/or the rhizosphere soil of C. odorata may be involved in the alteration of the microbial community as the observations in those publications (Table 3). The identification of these compounds is also necessary. In addition, the observations described in this section are controversial that whether C. odorata increases the population of fungal pathogen such as Fusarium spp. or suppressed the population. More sophisticate investigations are necessary in the future to explain the interaction of C. odorata with the microbial population.
3. Allelopathy of C. odorata
The interaction between the invasive plants and the indigenous plant species in the invasive ranges is also one of the important factors for the naturalization of the invasive plants [30,31,48,50,51]. According to the novel weapons hypothesis, the competitive ability of the invasive plants against the indigenous plants are high because allelochemicals (weapons) released from the invasive plants inhibit the germination and growth of the indigenous plant species. The inhibitory effect of the allelochemicals was greater on the indigenous plant species in the invasive ranges than that on the neighboring plant species in the native ranges of the invasive plant species. These allelochemicals were new to the indigenous plant species in the invasive ranges. However, the co-evolutional history of the neighboring plant species with the invasive plant species allows for those neighboring plant species to gain the tolerance to these allelochemicals. Therefore, these allelochemicals are more effective on the indigenous plant species in the invasive ranges than the neighboring plant species in the native ranges of the invasive plants, and contribute to the invasions [48,49,50]
The seeds of C. odorata, which were obtained from the population in the native (Mexico) and invasive (China) ranges, were germinated and grown in a field in China under natural condition for 10 months. The biomass of C. odorata seeds obtained from the invasive range was greater than that from the native range. Both the C. odorata from Mexico and China were grown together with other plant species from Mexican origin and those from Chinese origin. The biomass of the C. odorata seed variety grown with Chinese original plant species was greater than that grown with Mexican original plant species, while these Mexican original plant species grew well with both C. odorata than the Chinese original plant species [114,115,116]. These observations suggest that the Mexican original plant species may have stronger resistance to C. odorata than the Chinese original plant species. The co-evolutional history of the neighboring plant species in the native ranges of C. odorata are longer than the plant species in the invasive ranges, and those neighboring plant species in the native ranges may have more competitive ability to C. odorata than the plant species in the invasive ranges. These observations may be consistent with the novel weapons hypothesis described above.
Allelopathy is the chemical interaction between donner plants and their neighboring plants through certain secondary substances defied as allelochemicals. Allelochemicals are synthesized in the donner plants and released into the vicinity of the donner plants either by root exudation, volatilization, rainfall leachates or decomposition of plant residues and litter. Since allelochemicals are thought to be stored in certain plant tissues until their releasing into the vicinity of donner plants [117,118,119,120], many researchers determined the allelopathic activity of C. odorata in its plant residues and extracts from different plant parts of C. odorata (Table 3).
3.1. Allelopathic Activity of the Residue of C. odorata
The incorporation of C. odorata leaves into the crop field soil resulted in the suppression of the growth of Capsicum annuum L. and Solanum melongena L. [121]. The leaf residues of C. odorata incorporated into soil under greenhouse conditions inhibited the growth of Eleusine indica (L.) Gaertn., Cyperus iria L. and Ageratum conyzoides L. [122], and Lycopersicon esculentum Mill. [123]. The leaf powder of C. odorata also suppressed the growth of Crassocephalum crepidioides (Benth.) S. Moore [124]. These observations suggest that C. odorata leaves contain some allelochemicals, and those allelochemical were released into the soil during their decomposition process.
3.2. Allelopathic Activity of the Extracts of C. odorata
Aqueous extracts of leaves and roots of C. odorata suppressed the germination and growth of five plant species from the native ranges of C. odorata such as Bidens pilosa L. (tropical America origin), Ageratum conyzoides L. (tropical America), Amaranthus spinosus L. (tropical America), Conyza sumatrensis (Retz.) E. Walker (South America), and Chenopodium ambrosioides L. (Central and South America), and other five plant species from the invasive ranges such as Rottboellia exaltata (Lour.) Clayton (tropical Africa and Asia), Digitaria sanguinalis (L.) Scop. (South Europa, North Africa and Asia), Hemisteptia lyrata Bunge. (Eurasia and north Africa), Youngia japonica (L.) DC. (Eastern Asia), and Dicliptera chinensis (L.) Juss. (Eastern Asia). Growth inhibitory effect on these ten test plant species was higher with the leaf extracts than the root extracts of C. odorata, and on plant species from the invasive ranges than that from native ranges of C. odorata [125]. It was also reported that the growth inhibitory activity of the leaf extracts of C. odorata collected from invasive ranges (China) was higher than that collected from native ranges (Mexico) against the growth of the indigenous plant species in China; Eupatorium japonicum Thunb., Eupatorium stoechadosmum Hance and Eupatorium lindleyanum DC [114]. Those observations suggest that the inhibitory activity of C. odorata extracts from the invasive ranges was greater than that from the native ranges, and the plant species from its invasive ranges were more susceptible to the extracts than those from the native ranges of C. odorata.
The seeds of Lathyrus sativus L. were dipped in the aqueous leaf extract of C. odorata for 8 h, washed with distill water, and dried. Then, the seeds were sown and the germination and growth of Lathyrus sativus were determined after 10 days and 30 days, respectively. The treatments resulted in the suppression of the germination and the growth of Lathyrus sativus [126]. The observation suggests that some allelochemicals may be absorbed into the seeds and suppress the seed germination and growth processes.
Aqueous leaf extracts of C. odorata showed the suppression of the germination of weed plant species, Cynodon dactylon L., Crassocephalum crepidioides (Benth.) S.Moore, and Ageratum conyzoides L. [124], and the growth of Eleusine indica (L.) Gaertn., Cyperus iria L., and Ageratum conyzoides L. [122]. Aqueous extracts of whole plants of C. odorata inhibited the germination and growth of Echinochloa crus-galli (L.) P.Beauv., and Amaranthus viridis L. [127].
Aqueous leaf extracts of C. odorata also inhibited the germination and growth of crop plant species, Sorghum bicolor (L.) Moench, Zea mays L., Phaseolus vulgaris L., Vigna radiata (L.) R.Wilczek, and Centrosema pubescens Benth. [128,129,130], Brassica chinensis L. [131], Glycine max L., and Gossypium hirsutum L. [132], and Lycopersicum esculentum Mill. [123]. Aqueous extracts of the aerial parts of C. odorata inhibited the germination and growth of Sesame indicum L., Brassica nigra (L.), Brassica juncea (L.) Czern., and Raphenus raphanistrm L. [133]. The leaves of C. odorata were extracted with methanol, and the extract was sprayed to Amaranthus spinosus L. and Amaranthus spinosus L. The treatments resulted in the reduction of the growth parameters of both plant species such as their plant high, leaf areas, root length, and plant masses [134].
These observations suggest that the aqueous and methanol extracts of all parts of C. odorata possess allelopathic activity on the germination and growth of both weed and crop plant species, and may contain water and/or methanol extractable allelochemicals. Effectiveness of these allelochemicals was greater on the plant species from the invasive ranges than the native ranges of C. odorata.
3.3. Mechanism of the Inhibition
The inhibitory mechanism of the extracts of C. odorata on the germination and growth of several plant species was also investigated. Aqueous leaf extracts of C. odorata suppressed amylase activity in the seeds of Cicer arietinum L., and Cajanus cajan (L.) Millsp. [135]. Aqueous extracts of leaves, stems, and roots of C. odorata were sprayed onto 21-day-old plants of Ageratum conyzoides L. every two days for 28 days. The treatments resulted in the reduction in the contents of chlorophyll, carotenoids, and stomata number in the leaves of A. conyzoides [136]. The leaves of C. odorata were soaked in water for 48 h, and the soaking water also reduced the contents of chlorophyll and protein, and cell division of Allium cepa L. [137]. The hexane fraction obtained from the aqueous methanol leaf extracts of C. odorata suppressed chlorophylls and carotenoid contents in the leaves of Echinochloa crus-galli (L.) P.Beauv. [127]. These observations suggest that the extracts disturb amylase activity in the seeds, cell division, and reduce the pigments of photosynthesis, which may cause the suppression of the photosynthesis and growth. The induction of amylase is essential for seed germination because this enzyme triggers starch degradation of the reserve starch in seeds and enable the seeds to germinate and grow [138,139]. Total concentrations of terpenoids, flavonoids, tannins, steroids, and alkaloids were determined in the aqueous extracts of C. odorata (Hamidi et al., 2014). Total terpenoids, flavonoids, phenols, and steroids were also determined in the ethanol extracts of C. odorata [134]. However specific compounds did not identify in these extracts. The observations described in this section suggest that the extracts of C. odorata are allelopathic and contain allelochemicals. Allelochemicals in the extracts of C. odorata should be identified in the furfure. Table 4 shows the allelopathic activity of C. odorata described in the Section 3.
4. Diversity of the Secondary Metabolites in C. odorata
Some of the plant secondary metabolites function as defense molecules against herbivores, pathogens, and competing neighboring plants. These compounds are important for the plant’s survival and fitness, and represent the adaptive characters of the plant species that have been subjected to natural selection during the evolution. The pattern of the plant secondary metabolites is complex, and changes in an organ- and tissue-specific way, and during the developmental stages of the plants. The biosynthesis of some secondary metabolites is induced or synthesized de-novo upon herbivore-attack, pathogen–infection and competition with neighboring plants [117,140,141,142,143,144].
The significant difference in the metabolomic profiles between C. odorata obtain from the native ranges (USA) and that from the invasive ranges (South Africa) were observed by the ultra-performance liquid chromatography-mass spectrometry. The major differences were high concentrations of flavonoids and flavone glycosides in the C. odorata from the invasive ranges comparing to those from the native ranges [145]. Although the function of these metabolites and their significance to the invasive behaviors of C. odorata are not apparent, the observation suggests that the invasive plant species may employ the metabolic flexibility and/or rapid adaptive evolution to succeed as the invasive plant species.
As already described in the Section 3.2, the inhibitory activity of the extracts of C. odorata collected from invasive ranges was higher than that collected from the native ranges [114,125]. Total phenolic concentration in the leaves and stems of C. odorata obtained from the invasive ranges (China) was greater than that from the native ranges (Mexico) [39]. Concentrations of flavonoids; dihydrokaempferol-3-methoxy ether, isosakuranetin, kaempferide-4’-methoxy ether, 3,5-dihy-droxy-7,4’-dimethoxyflavone, acutellerin-4’,6,7-trimethy ether and 4’,5,6,7-tetramethoxyflavone were also greater in C. odorata in the invasive ranges (China, Laos, Thailand, Vietnam, Philippines, Sri Lanca, Malaysia) than those in the native ranges (USA, Mexico, Puerto Rico, Trinidad, and Tobago) [41]. The concentration of a flavonoid, odoratin in C. odorata obtained from an invasive range was 2.4-fold greater than that from the native ranges. However, the specific compounds of the increasing phenolics were not identified. The function of these flavonoids on the allelopathy of C. odorata is also not clear. In addition, the chemical structure of odratin in the publication is not correct [46] (Figure 3).
Pharmacological investigations showed that C. odorata contains secondary metabolites in many chemical classes, such as flavonoids, phenolic acids, saponins, terpenoids and tannins. Some of those compounds were related to the pharmacological activity such as analgesic, antipyretic, anti-inflammatory, anti-diabetic, anticancer, and antioxidant activity [146,147,148,149,150,151,152,153,154]. Although most of those identified compounds have not yet been related to the invasiveness of the plant species, some of them may be involved in the allelopathy and defense functions against herbivores, nematodes, and fungal pathogens.
Benzoic acid and cinnamic acid derivatives such as p-hydroxybenzoic acid, protocatechuic acid, and p-coumaric acid were identified as the major compounds in the leaves of C. odorata [146]. These compounds are synthesized by shikimic acid pathway [155,156]. Benzoic acid and cinnamic acid derivatives have been found in a wide range of plants, decomposition products of plants, and plant rhizosphere soil. The involvement of those compounds in allelopathy and their mechanisms of the action have been investigated in other plant species [157,158,159]. Benzoic acid and cinnamic acid derivatives reduced the transmembrane electrochemical potential of the plasma membrane of the cells. The depolarization of the membranes caused a nonspecific efflux of both cations and anions, and affected the membrane permeability and the uptake of ions and nutrients. These compounds caused structural alteration in the membranes including a variety of membrane proteins. The changes in ion flux through the membranes affected plant water status including the stomatal functions. These compounds also suppressed several enzyme activities involved in several physiological processes such as respiration, phytohormone synthesis, protein synthesis, and synthesis of some other secondary metabolites [157,158,159]. In addition, their derivative, chlorogenic acid was also isolated from the C. odorata leaves [160]. Chlorogenic acid was reported to inhibit blue-green alga Microcystis aerugonisa Kützing [161] (Figure 4).
Several flavonoids were identified in the leaf and flower extracts of C. odorata [147,149,150,152]. Flavonoids are polyphenolic secondary metabolites having a 15-carbon skeleton, consisting of two benzene rings and a heterocycling ring, and synthesized from chalcone. Many of the flavonoids showed anti-herbivore, anti-fungal, and anti-bacteria activity [162,163,164]. Sakuranetin was isolated from the leaf extracts of C. odorata [165]. The compound is known to act as phytoalexin against pathogen infection [166,167]. Quercetin and kaempferol were also identified in the leaf extracts C. odorata [145,168] and both compounds were reported to work as allelopathic agents in the other plant species. Quercetin inhibited the growth of several pant species [169,170], and the plant mitochondrial function [171,172]. Kaempferol showed the growth inhibitory activity on blue-green alga Microcystis aerugonisa Kützing [173]. Kaempferol reduced the efficiency of the photosystem II in the chloroplasts [171] (Figure 4).
Several terpenoids were identified in the essential oil and leaf extracts of C. odorata [68,148]. Terpenoids in the most plants are synthesized through the mevalonate pathway from acetyl-CoA, and many terpenoids were reported to be involved in the defense function of plants such as anti-fungal, anti-bacterial, and anti-feeding activities, and in the interaction with insects such as for the pollination, and the attracting predators of their natural enemy [174,175,176,177,178,179]. The essential oil mixture, of which major constituents were monoterpenes and sesquiterpens, showed allelopathic activity [180,181,182]. Among terpenoids found in C. odorata, monoterpenes, α-pinene and 1,8-cineole were identified in its leaf extracts [68]. α-Pinene and 1,8-cineole showed allelopathic activity and a mixture of both compounds exhibited synergistic effect for the allelopathic activity on the growth of Solanum elaeagnifolium Cav. [183]. 1,8-Cineole strongly inhibits the roots and coleoptile growth of Echinochloa crus-galli (L.) P.Beauv. and Senna obtusifolia (L.) H.S.Irwin et Barneby, and reduced all stages of mitosis of the root tips of Allium cepa L. [184] (Figure 4). Several pyrrolizine alkaloids were also identified in the leaf extracts of C. odorata and showed anti-herbivore and anti-nematode activity as already described in the Section 2.1.
As describe above, some of cinnamic acid and benzoic acid derivatives, flavonoids, and terpenoids found in C. odorata may affect the physiological processes of the neighboring plant species, causing growth inhibition and reducing their fitness as allelochemicals. Pyrrolizine alkaloids found in C. odorata may also have defense functions against herbivores, and pathogenic fungi, microbes, and nematodes. Therefore, these compounds may contribute to the invasiveness and naturalization in the introduced rages of C. odorata.
5. Conclusions
C. odorata is highly invasive and has naturalized in many countries. The paper described the interaction between the species and their natural enemies, and the species and the indigenous plant species in the invasive ranges for the naturalization of the invasive plants. The extracts of all plant parts, residues, and rhizosphere soil of C. odorata showed the inhibitory activity against various insects, parasitic nematodes, and fungal pathogens, and several pyrrolizidine alkaloids were identified in the roots and flower heads of C. odorata. Pyrrolizidine alkaloids are toxic against insects and other organisms. Therefore, pyrrolizidine alkaloids of C. odorata may act as chemical protection agents against natural enemies including herbivores.
The rhizosphere soil, extracts, and residues of all plant parts showed the suppression of the germination of growth of many plant species including indigenous plant species in the invasive ranges. The indigenous plant species from the invasive ranges of C. odorata were more susceptible to the extracts than plant species from the native ranges of C. odorata. Those observations suggest that C. odorata possess allelopathic activity, and may contain certain allelochemicals. These allelochemicals may be more effective on the plant species from the invasive ranges of C. odorata than on the plant species from its native ranges.
Pharmacological investigations showed that C. odorata contains secondary metabolites in many chemical classes, such as phenolic acids, flavonoids, saponins, tannins, and terpenoids. Although many of these compounds have not been related to the allelopathy of C. odorata, benzoic acid and cinnamic acid derivatives such as p-hydroxybenzoic acid, protocatechuic acid, and p-coumaric acid; flavonoids such as sakuranetin, quercetin and kaempferol; and monoterpenes such as α-pinene and 1,8-cineole identified in the leaves of C. odorata were reported to be involved in the allelopathy of other plant species. Therefore, these compounds may also work as allelopathic agents for C. odorata and inhibit the germination and growth of the neighboring plant species.
These characteristics of C. odorata for the defense function against their natural enemies such as insects, parasitic nematodes, and fungal pathogens, and allelopathic potential may contribute to the invasiveness and naturalization of C. odorata in the new habitats as invasive plant species. However, allelochemicals are able to work only when they are released into the neighboring environment of the donner plants either by root exudation, volatilization, rainfall leachates, or decomposition of plant litter and residues. Therefore, it is necessary to determine the concentration of these allelochemicals in the neighboring environment of C. odorata such as the rhizosphere soil.
Funding
This research received no external funding.
Institutional Review Statement
Not applicable.
Informed Consent Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Zachariades, C.; Day, M.; Muniappan, R.; Reddy, G.V.P. Chromolaena odorata (L.) King and Robinson (Asteraceae). In Biological Control of Tropical Weeds Using Arthropods; Muniappan, R., Reddy, G.V.P., Raman, A., Eds.; Cambridge University Press: London, UK, 2009; pp. 130–140. [Google Scholar]
- Joshi, C.; van Andel, J.; Skidmore, A.; De Leeuw, J.; Duren, I.V. Morphological plasticity of corms in enhancing invasion of Chromolaena odorata. Banko Janakari 2011, 21, 3–12. [Google Scholar] [CrossRef] [Green Version]
- Tripathi, R.S.; Yadav, A.S.; Kushwaha, S.P.S. Biology of Chromolaena odorata, Ageratina adenophora and Ageratina riparia: A Review. In Invasive Alien Plants: An Ecological Appraisal for the Indian Subcontinent; Bhatt, J.R., Singh, J.S., Singh, S.P., Tripathi, R.S., Kohli, R.K., Eds.; CAB International: Croydon, UK, 2012; pp. 43–56. [Google Scholar]
- McFadyen, R.E.C.; Skarratt, B. Potential distribution of Chromolaena odorata (Siam Weed) in Australia, Africa and Oceania. Agric. Ecosyst. Environ. 1996, 59, 89–96. [Google Scholar] [CrossRef]
- Timbilla, J.A.; Braimah, H. Highlights from work on Chromolaena odorata in Ghana. In Proceedings of the Second International Workshop on Biological Control and Management of Chromolaena odorata, Bogor, Indonesia, 4–8 February 1991; Muniappan, R., Ferrar, P., Eds.; Biotrop: Bogor, Indonesia, 2002; pp. 105–112. [Google Scholar]
- Kushwaha, S.P.S.; Ramakrishnan, P.S. Observations on growth of Eupatorium odoratum L. and Imperata cylindrica (L.) Beauv. var. major under different light and moisture regimes. Proc. Indian Natl. Sci. Acad. 1982, B48, 689–693. [Google Scholar]
- Tripathi, R.S.; Kushwaha, S.P.S.; Yadav, A.S. Ecology of three invasive species of Eupatorium: A review. Int. J. Ecol. Environ. Sci. 2006, 32, 301–326. [Google Scholar]
- Zhang, L.L.; Wen, D.Z. Structural and physiological responses of two invasive weeds, Mikania micrantha and Chromolaena odorata, to contrasting light and soil water conditions. J. Plant Res. 2009, 122, 69–79. [Google Scholar] [CrossRef] [PubMed]
- Scott, L.J.; Lange, C.L.; Graham, G.C.; Yeates, D.K. Genetic diversity and origin of Siam weed (Chromolaena odorata) in Australia. Weed Technol. 1998, 12, 27–31. [Google Scholar] [CrossRef]
- Kriticos, D.J.; Yonow, T.; McFadyen, R.E. The potential distribution of Chromolaena odorata (Siam weed) in relation to climate. Weed Res. 2005, 45, 246–254. [Google Scholar] [CrossRef]
- Invasive Species Compendium, Chromolaena odorata (Siam Weed). Available online: https://www.cabidigitallibrary.org/doi/full/10.1079/cabicompendium.23248 (accessed on 12 December 2022).
- Global Invasive Species Databas, Species Profile: Chromolaena odorata. Available online: http://www.iucngisd.org/gisd/speciesname/Chromolaena+odorata (accessed on 12 December 2022).
- Royal Botanical Gardens, Kew, Chromolaena odorata. Available online: https://powo.science.kew.org/taxon/urn:lsid:ipni.org:names:57443-2 (accessed on 12 December 2022).
- Goodall, J.M.; Erasmus, D.J. Review of the status and integrated control of the invasive alien weed, Chromolaena odorata, in South Africa. Agric. Ecosyst. Environ. 1996, 56, 151–164. [Google Scholar] [CrossRef]
- Prasad, U.K.; Muniappan, R.; Ferrar, P.; Aeschliman, J.P.; De Foresta, H. Distribution, Ecology and Management of Chromolaena odorata; University of Guam: Mangilao, Guam, 1996; Publication no. 202; pp. 1–203. [Google Scholar]
- Prashanthi, S.K.; Kulkarni, S. Aureobasidium pullulans, a potential mycoherbicide for biocontrol of Eupatorium [Chromolaena odorata (L.) King and Robinson] weed. Curr. Sci. 2005, 88, 18–21. [Google Scholar]
- Leslie, A.J.; Spotila, J.R. Alien plant threatens Nile crocodile (Crocodylus niloticus) breeding in Lake St. Lucia, South Africa. Biol. Conserv. 2001, 98, 347–355. [Google Scholar] [CrossRef]
- Dumalisile, L. Effects of Chromolaena odorata on Mammalian Biodiversity in Hluhluwe-iMfolozi Park, South Africa. Ph.D. Dissertation, University of Pretoria, Pretoria, South Africa, 2018; pp. 1–58. Available online: https://repository.up.ac.za/handle/2263/26178 (accessed on 12 December 2022).
- Norgrove, L.; Tueche, R.; Dux, J.; Yonghachea, P. Chromolaena odorata: The benevolent dictator. Chromolaena odorata Newsletter. 2008, 17, 1–3. [Google Scholar]
- Thapa, L.B.; Kaewchumnong, L.; Sinkkonen, A.; Sridith, L. Impacts of invasive Chromolaena odorata on species richness, composition and seedling recruitment of Shorea robusta in a tropical Sal forest, Nepal. Songklanakarin J. Sci. Technol. 2016, 38, 683–689. [Google Scholar]
- Agboola, O.O.; Muoghalu, J.I. Changes in species diversity, composition, growth and reproductive parameters of native vegetation invaded by Chromolaena odorata and Tithonia diversifolia in osun state, southwest Nigeria. FUTA J. Res. Sci. 2015, 11, 217–230. [Google Scholar]
- Rao, R.R. Changing pattern in the Indian flora. Bull. Bot. Surv. India 1977, 19, 156–166. [Google Scholar]
- Azmi, M. Status and Biological Control of Chromolaena odorata in Malaysia. In Proceedings of the Fifth International Workshop on Biological Control and Management of Chromolaena odorata, Durban, South Africa, 23–25 October 2000; Zachariades, C., Muniappan, R., Strathie, R.W., Eds.; Plant Protection Research Institute: Pretoria, South Africa, 2000; pp. 23–25. [Google Scholar]
- McFadyen, R.E.C. Ecology of Chromolaena odorata in the Neotropics. In Proceedings of the First International Workshop on Biological Control of Chromolaena odorata, Bangkok, Thailand, 29 February–4 March 1988; Prasad, U., Muniappan, R., Aeschliman, E.F.J.P., de Foresta, H., Eds.; Agricultural Experiment Station, University of Guam: Mangilao, Guam, 1988; pp. 13–20. [Google Scholar]
- McFadyen, R.E.C. Siam weed: A new threat to Australia’s north. Plant Prot. Qtly. 1989, 4, 3–7. [Google Scholar]
- Witkowski, E.T.F.; Wilson, M. Changes in density, biomass, seed production and soil seed banks of the non-native invasive plant, Chromolaena odorata, along a 15 year chronosequence. Plant Ecol. 2001, 152, 13–27. [Google Scholar] [CrossRef]
- Groves, R. A Short History of Biological Invasion in Australia. In Biogeography of Mediterranean Invasion; Groves., R., di Castri, F., Eds.; Cambridge University Press: Cambridge, UK, 1991; pp. 59–63. [Google Scholar]
- Thompson, J.D.; McNeilly, T.; Gray, A.J. Population variation in Spartina anglica C.E. Hubbard. I. Evidence from a common garden experiment. New Phytol. 1991, 117, 115–128. [Google Scholar] [CrossRef]
- Mack, R.M. Predicting the identity and fate of plant invaders: Emergent and emerging approaches. Biol. Conserv. 1996, 78, 107–121. [Google Scholar] [CrossRef]
- Chengxu, W.; Mingxing, Z.; Xuhui, C.; Bo, Q. Review on allelopathy of exotic invasive plants. Procedia Eng. 2011, 18, 240–246. [Google Scholar] [CrossRef] [Green Version]
- Warren, R.J.; Candeias, M.M.; Labatore, A.; Olejniczak, M.; Yang, L. Multiple mechanisms in woodland plant species invasion. J. Plant Ecol. 2019, 12, 201–209. [Google Scholar] [CrossRef]
- Yadav, A.S.; Tripathi, R.S. Population dynamics of the ruderal weed Eupatorium odoratum and its natural regulation. Oikos 1981, 36, 355–361. [Google Scholar] [CrossRef] [Green Version]
- Ramakrishnan, P.S. Shifting Agriculture and Sustainable Development—An Interdisciplinary Study from North-Eastern India. In Man and The Biosphere Series; Unesco: Paris, France, 1992; Volume 10, pp. 1–441. [Google Scholar]
- Moni, N.S.; Subramoniam, R. Essential oil from Eupatorium odoratum—A common weed in Kerala. Indian For. 1960, 86, 209. [Google Scholar]
- Hoevers, R.; M’Boob, S.S. The status of Chromolaena odorata (L.) R.M. King and H. Robinson in West and Central Africa. In Proceedings of the Third International Workshop on Biological Control and Management of Chromolaena odorata, Abidjan, Côte d’Ivoire, 15–19 November 1993; Prasad, U.K., Muniappan, R., Aeschliman, P.F.J.P., de Foresta, H., Eds.; Agricultural Experiment Station, University of Guam: Mangilao, Guam, 1996; pp. 1–5. [Google Scholar]
- Macdonald, I.A.W. Alien trees, shrubs and creepers invading indigenous vegetation in the Hluhluwe-Umfolozi Game Reserve Complex in Natal. Bothalia 1983, 14, 949–959. [Google Scholar] [CrossRef] [Green Version]
- Muniappan, R.; Marutani, M. Ecology and distribution of C. odorata in Asia and the Pacific. In Proceedings of the First International Workshop on Biological Control of Chromolaena odorata, Bangkok, Thailand, 29 February–4 March 1988; Prasad, U., Muniappan, R., Aeschliman, E.F.J.P., de Foresta, H., Eds.; Agricultural Experiment Station, University of Guam: Mangilao, Guam, 1988; pp. 21–24. [Google Scholar]
- Ye, W.H.; Mu, H.P.; Cao, H.L.; Ge, X.J. Genetic structure of the invasive Chromolaena odorata in China. Weed Res. 2004, 44, 129–135. [Google Scholar] [CrossRef]
- Zheng, Y.; Feng, Y.; Liao, Z.; Li, W.; Xiao, H.; Sui, H. Invasive Chromolaena odorata has similar size but higher phenolic concentration than native conspecifics. Evol. Ecol. Res. 2013, 15, 769–781. [Google Scholar]
- Yu, X.Q.; He, T.H.; Zhao, J.L.; Li, Q.M. Invasion genetics of Chromolaena odorata (Asteraceae): Extremely low diversity across Asia. Biol. Invasions 2014, 16, 2351–2366. [Google Scholar] [CrossRef]
- Li, W.; Zheng, Y.; Zhang, L.; Lei, Y.; Li, Y.; Liao, Z.; Li, Y.; Feng, Y. Postintroduction evolution contributes to the successful invasion of Chromolaena odorata. Ecol. Evol. 2020, 10, 1252–1263. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shao, X.; Li, Q.; Lin, L.; He, T. On the origin and genetic variability of the two invasive biotypes of C. odorata. Biol. Invasions 2018, 20, 2033–2046. [Google Scholar] [CrossRef]
- Paterson, I.D.; Zachariades, C. ISSRs indicate that Chromolaena odorata invading southern Africa originates in Jamaica or Cuba. Biol. Control 2013, 66, 132–139. [Google Scholar] [CrossRef]
- Liao, Z.Y.; Scheepens, J.F.; Li, W.T.; Wang, R.F.; Zheng, Y.L.; Feng, Y.L. Biomass reallocation and increased plasticity might contribute to successful invasion of Chromolaena odorata. Flora 2019, 256, 79–84. [Google Scholar] [CrossRef]
- Li, W.T.; Zheng, Y.L.; Wang, R.F. Extension of the EICA hypothesis for invasive Chromolaena odorata. Acta Oecol. 2022, 114, 103803. [Google Scholar] [CrossRef]
- Zheng, Y.L.; Feng, Y.L.; Zhang, L.K.; Callaway, R.M.; Valiente-Banuet, A.; Luo, D.Q.; Liao, Z.Y.; Lei, Y.B.; Barclay, G.F.; Silva-Pereyra, C. Integrating novel chemical weapons and evolutionarily increased competitive ability in success of a tropical invader. New Phytol. 2015, 205, 1350–1359. [Google Scholar] [CrossRef]
- Lockwood, J.L.; Simberloff, D.; McKinney, M.L.; Von Holle, B. How many, and which, plants will invade natural areas. Biol. Invasions 2001, 3, 1–8. [Google Scholar] [CrossRef]
- Callaway, R.M.; Aschehoug, E.T. Invasive plants versus their new and old neighbors: A mechanism for exotic invasion. Science 2000, 290, 521–523. [Google Scholar] [CrossRef] [PubMed]
- Callaway, R.M.; Ridenour, W.M. Novel weapons: Invasive success and the evolution of increased competitive ability. Front. Ecol. Environ. 2004, 2, 419–426. [Google Scholar] [CrossRef]
- Cappuccino, N.; Arnason, J.T. Novel chemistry of invasive exotic plants. Biol. Lett. 2006, 2, 189–193. [Google Scholar] [CrossRef] [Green Version]
- Meiners, S.J.; Kong, C.H.; Ladwig, L.M.; Pisula, N.L.; Lang, K.A. Developing an ecological context for allelopathy. Plant. Ecol. 2012, 213, 1861–1867. [Google Scholar] [CrossRef]
- Kato-Noguchi, H. Involvement of allelopathy in the invasive potential of Tithonia diversifolia. Plants 2020, 9, 766. [Google Scholar] [CrossRef]
- Kato-Noguchi, H.; Kurniadie, D. Allelopathy of Lantana camara as an invasive plant. Plants 2021, 10, 1028. [Google Scholar] [CrossRef]
- Kato-Noguchi, H. Allelopathy of knotweeds as invasive plants. Plants 2022, 11, 3. [Google Scholar] [CrossRef]
- Kato-Noguchi, H.; Kurniadie, D. Allelopathy and allelochemicals of Leucaena leucocephala as an invasive plant species. Plants 2022, 11, 1672. [Google Scholar] [CrossRef]
- Kato-Noguchi, H. Allelopathy and allelochemicals of Imperata cylindrica as an invasive plant species. Plants 2022, 11, 2551. [Google Scholar] [CrossRef]
- Keane, R.M.; Crawley, M.L. Exotic plant invasions and the enemy release hypothesis. Trend. Ecol. Evol. 2002, 17, 164–170. [Google Scholar] [CrossRef]
- Blumenthal, D.M. Interactions between resource availability and enemy release in plant invasion. Ecol. Let. 2006, 9, 887–895. [Google Scholar] [CrossRef]
- McFadyen, R.E.C. Phytophagous insects recorded from Chromolaena odorata. Chromolaena odorata Newsletter. 1988, 2, 5–23. [Google Scholar]
- Barreto, R.W.; Evans, H.C. The mycobiota of the weed Chromolaena odorata in southern Brazil with particular reference to fungal pathogens for biological control. Mycol. Res. 1994, 98, 1107–1116. [Google Scholar] [CrossRef]
- Zhang, L.H.; Feng, Y.L. Potential biological control agents of Chromolaena odorata. Chin. J. Biol. Control 2007, 23, 83–88. [Google Scholar]
- Kluge, R.L.; Caldwell, P.M. Phytophagous insects and mites on Chromolaena odorata (Compositae, Eupatoreae) in Southern Africa. J. Entomol. Soc. South. Afr. 1992, 55, 159–161. [Google Scholar]
- Blossey, B.; Notzold, R. Evolution of increased competitive ability in invasive nonindigenous plants—A hypothesis. J. Ecol. 1995, 83, 887–889. [Google Scholar] [CrossRef]
- Muller-Scharer, H.; Schaffner, U.; Steinger, T. Evolution in invasive plants: Implications for biological control. Trends Ecol. Evol. 2004, 19, 417–422. [Google Scholar] [CrossRef]
- Uyi, O.O.; Obi, B.N. Evaluation of the repellent and insecticidal activities of the leaf, stem and root powders of Siam weed (Chromolaena odorata) against the cowpea beetle, Callosobruchus maculatus. J. Appl. Sci. Environ. Manag. 2017, 21, 511–518. [Google Scholar]
- Uyi, O.O.; Igbinoba, O.G. Repellence and toxicological activity of the root powder of an invasive alien plant, Chromolaena odorata (L.)(Asteraceae) against Callosobruchus maculatus (FAB.) (Coleoptera: Chrysomelidae). Anim. Res. Int. 2016, 13, 2510–2517. [Google Scholar]
- Bouda, H.; Tapondjou, L.A.; Fontem, D.A.; Gumedzoe, M.Y.D. Effect of essential oils from leaves of Ageratum conyzoides, Lantana camara and Chromolaena odorata on the mortality of Sitophilus zeamais (Coleoptera, Curculionidae). J. Stored Prod. Res. 2001, 37, 103–109. [Google Scholar] [CrossRef]
- Lawal, O.A.; Opoku, A.R.; Ogunwande, I.A. Phytoconstituents and insecticidal activity of different solvent leaf extracts of Chromolaena odorata L., against Sitophilus zeamais (Coleoptera: Curculionidae). Eur. J. Med. Plants 2015, 5, 237–247. [Google Scholar] [CrossRef]
- Mathur, B.M.; Davou, B.J. Comparative larvicidal property of leaf extract of Chromolaena odorata L. (Composidae) and Chlopyrifos (Organophosphorus Compound) on Simulium larvae. Biomed. Environ. Sci. 2007, 20, 313–316. [Google Scholar]
- Udebuani, A.C.; Abara, P.C.; Obasi, K.O.; Okuh, S.U. Studies on the insecticidal properties of Chromolaena odorata (Asteraceae) against adult stage of Periplaneta americana. J. Entomol. Zool. Stud. 2015, 3, 318–321. [Google Scholar]
- Nneji, E.T.; Uddin, I.I.R.O.; Musa, A.A. Effects of Boerhavia diffusa L. nom. cons. and Chromolaena odorata (L.). M. King & H. Rob. extracts on some field insect pests of okra (Aabelmoschus esculentus (L.) Moench). Acta Agric. Slov. 2020, 115, 133–139. [Google Scholar]
- Gorawade, V.; Attar, U.; Shiragave, P. Bioefficacy and GC-MS analysis of Chromolaena odorata and Leonotis nepetifolia leaf extracts against Spodoptera litura. J. Crop Prot. 2022, 11, 361–375. [Google Scholar]
- Biller, A.; Boppré, M.; Witte, L.; Hartmann, T. Pyrrolizidine alkaloids in Chromolaena odorata. Chemical and chemoecological aspects. Phytochemistry 1994, 35, 615–619. [Google Scholar] [CrossRef]
- Dube, N.; van Heerden, F.R.; Zachariades, C.; Uyi, O.O.; Munyai, T.C. First record of pyrrolizidine alkaloids in the southern African biotype of Chromolaena odorata (Asteraceae). S. Afr. J. Bot. 2021, 139, 19–25. [Google Scholar] [CrossRef]
- Boppré, M. Lepidoptera and pyrrolizidine alkaloids. Exemplification of complexity in chemical ecology. J. Chem. Ecol. 1990, 16, 165–185. [Google Scholar] [CrossRef] [PubMed]
- Van Dam, N.M.; Vuister, L.W.; Bergshoeff, C.; de Vos, H.; van der Meijden, E.D. The “Raison D’être” of pyrrolizidine alkaloids in Cynoglossum officinale: Deterrent effects against generalist herbivores. J. Chem. Ecol. 1995, 21, 507–523. [Google Scholar] [CrossRef] [PubMed]
- Joshi, J.; Vrieling, K. The enemy release and EICA hypothesis revisited: Incorporating the fundamental difference between specialist and generalist herbivores. Ecol. Lett. 2005, 8, 704–714. [Google Scholar] [CrossRef]
- Gardner, D.R.; Thorne, M.S.; Molyneux, R.J.; Pfister, J.A.; Seawright, A.A. Pyrrolizidine alkaloids in Senecio madagascariensis from Australia and Hawaii and assessment of possible livestock poisoning. Biochem. Syst. Ecol. 2006, 34, 736–744. [Google Scholar] [CrossRef]
- Stegelmeier, B.L.; Colegate, S.M.; Brown, A.W. Dehydropyrrolizidine alkaloid toxicity, cytotoxicity, and carcinogenicity. Toxins 2016, 8, 356. [Google Scholar] [CrossRef]
- Cheeke, P.R. Toxicity and metabolism of pyrrolizidine alkaloids. J. Anim. Sci. 1988, 66, 2343–2350. [Google Scholar] [CrossRef]
- Tamariz, J.; Burgueño-Tapia, E.; Vázquez, M.A.; Delgado, F. Pyrrolizidine Alkaloids. In The Alkaloids: Chemistry and Biology; Hans-Joachim Knölker, H.J., Ed.; Academic Press: Cambridge, UK, 2018; Volume 80, pp. 1–314. [Google Scholar]
- Griffiths, M.R.; Strobel, B.W.; Hama, J.R.; Cedergreen, N. Toxicity and risk of plant-produced alkaloids to Daphnia magna. Environ. Sci. Eur. 2021, 33, 10. [Google Scholar] [CrossRef]
- Kisielius, V.; Hama, J.R.; Skrbic, N.; Hansen, H.C.B.; Strobel, B.J.; Rasmussen, L.H. The invasive butterbur contaminates stream and seepage water in groundwater wells with toxic pyrrolizidine alkaloids. Sci. Rep. 2020, 10, 19784. [Google Scholar] [CrossRef]
- Conner, W.E. Tiger Moths and Woolly Bears: Behavior, Ecology, and Evolution of the Arctiidae; Oxford University Press: New York, NY, USA, 2009; pp. 1–303. [Google Scholar]
- Macel, M. Attract and deter: A dual role for pyrrolizidine alkaloids in plant-insect interactions. Phytochem. Rev. 2011, 10, 75–82. [Google Scholar] [CrossRef] [Green Version]
- Lambert, K.; Bekal, S. Introduction to Plant-Parasitic Nematodes. The Plant Health Instructor. Available online: https://www.apsnet.org/edcenter/disandpath/nematode/intro/Pages/IntroNematodes.aspx (accessed on 12 December 2022).
- Den Akker, S.E. Plant—Nematode interactions. Curr. Opin. Plant Biol. 2021, 62, 102035. [Google Scholar] [CrossRef]
- Pires, D.; Vicente, C.S.L.; Menéndez, E.; Faria, J.M.S.; Rusinque, L.; Camacho, M.J.; Inácio, M.L. The fight against plant-parasitic nematodes: Current status of bacterial and fungal biocontrol agents. Pathogens 2022, 11, 1178. [Google Scholar] [CrossRef]
- Adediran, J.A.; Adegbite, A.A.; Akinlosotu, T.A.; Agbaje, G.O.; Taiwo, L.B.; Owolade, O.F.; Oluwatosin, G.A. Evaluation of fallow and cover crops for nematode suppression in three agroecologies of south western Nigeria. Afr. J. Biotechnol. 2005, 4, 1034–1039. [Google Scholar]
- Odeyemi, I.S.; Afolami, S.O.; Adigun, J.A. Plant parasitic nematode relative abundance and population suppression under Chromolaena odorata (Asteraceae) fallow. Int. J. Pest Manag. 2013, 59, 79–88. [Google Scholar] [CrossRef]
- Sturhan, M.D. Studies on effects of Chromolaena odorata roots on root-knot nematodes (Meloidogyne incognita). Nematol. Mediterr. 1994, 22, 273–274. [Google Scholar]
- Odeyemi, I.S.; Afolami, S.O.; Adekoyejo, A.B. Integration of Glomus mosseae with Chromolaena odorata powder for suppression of Meloidogyne incognita on maize (Zea mays L.). Arch. Phytopathol. Plant Prot. 2013, 46, 1589–1597. [Google Scholar] [CrossRef]
- Thoden, T.C.; Boppré, M.; Hallmann, J. Pyrrolizidine alkaloids of Chromolaena odorata act as nematicidal agents and reduce infection of lettuce roots by Meloidogyne incognita. Nematology 2007, 9, 343–349. [Google Scholar]
- Hartmann, T.; Ehmke, A.; Eilert, U.; von Borstel, K.; Theuring, C. Sites of synthesis, translocation and accumulation of pyrrolizidine alkaloid N-oxides in Senecio vulgaris L. Planta 1989, 177, 98–107. [Google Scholar] [CrossRef]
- Koné, A.W.; Kassi, S.P.A.; Koffi, B.Y.; Masse, D.; Maïga, A.A.; Tondoh, J.E.; Kisaka, O.M.; Touré, G.P.T. Chromolaena odorata (L.) K&R (Asteraceae) invasion effects on soil microbial biomass and activities in a forest-savanna mosaic. Catena 2021, 207, 105619. [Google Scholar]
- Xiao, H.F.; Feng, Y.L.; Schaefer, D.A.; Yang, X.D. Soil fungi rather than bacteria were modified by invasive plants, and that benefited invasive plant growth. Plant Soil 2014, 378, 253–264. [Google Scholar] [CrossRef]
- Smith, S.E.; Read, D.J. Mycorrhizal Symbiosis, 3rd ed.; Academic Press: London, UK, 2008; pp. 1–815. [Google Scholar]
- Diagne, N.; Ngom, M.; Djighaly, P.I.; Fall, D.; Hocher, V.; Svistoonoff, S. Roles of Arbuscular Mycorrhizal Fungi on Plant Growth and Performance: Importance in Biotic and Abiotic Stressed Regulation. Diversity 2020, 12, 370. [Google Scholar] [CrossRef]
- Tang, H.; Hassan, M.U.; Feng, L.; Nawaz, M.; Shah, A.N.; Qari, S.H.; Liu, Y.; Miao, J. The critical role of arbuscular mycorrhizal fungi to improve drought tolerance and nitrogen use efficiency in crops. Front. Plant Sci. 2022, 13, 919166. [Google Scholar] [CrossRef]
- Schüβler, A.; Schwarzott, D.; Walker, C. A new fungal phylum, the Glomeromycota: Phylogeny and evolution. Mycol. Res. 2001, 105, 1413–1421. [Google Scholar] [CrossRef] [Green Version]
- Reynolds, H.L.; Packer, A.; Bever, J.D.; Clay, K. Grassroots ecology: Plant-microbe-soil interactions as drivers of plant community structure and dynamics. Ecology 2003, 84, 2281–2291. [Google Scholar] [CrossRef] [Green Version]
- Te Beest, M.; Stevens, N.; Olff, H.; Van Der Putten, W.H. Plant-soil feedback induces shifts in biomass allocation in the invasive plant Chromolaena odorata. J. Ecol. 2009, 97, 1281–1290. [Google Scholar] [CrossRef] [Green Version]
- Mangla, S.; Inderjit; Callaway, R.M. Exotic invasive plant accumulates native soil pathogens which inhibit native plants. J. Ecol. 2008, 96, 58–67. [Google Scholar] [CrossRef]
- Odunfa, V.S.A. Root exudation in cowpea and sorghum and the effect on spore germination and growth of some soil Fusaria. New Phytol. 1987, 80, 607–612. [Google Scholar] [CrossRef]
- Ju, H.; Han, L.; Wang, S.; Cong, D. Allelopathic effect of root exudates on pathogenic fungi of root rot in continuous soybean. J. Appl. Ecol. 2002, 13, 723–727. [Google Scholar]
- Asman, A.; Rosmana, A.; Sjam, S.; Fakhruddin, A.; Natsir, N.U. Antifungal activity of crude extracts of Ageratum conyzoides and Chromolaena odorata for management of Lasiodiplodia theobromae and Lasiodiplodia pseudotheobromae through in vitro evaluation. IOP Conf. Ser. Earth Environ. Sci. 2021, 886, 012008. [Google Scholar] [CrossRef]
- Ismail, A.M.; Cirvilleri, G.; Polizzi, G.; Crous, P.; Groenewald, W.J.Z.; Lombard, L. Lasiodiplodia species associated with dieback disease of mango (Mangifera indica) in Egypt Australas. Plant Pathol. 2012, 41, 649–660. [Google Scholar]
- Li, H.L.; Jayawardena, R.S.; Xu, W.; Hu, M.; Li, X.H.; Liu, J.H.; Hyde, K.D.; Yan, J. Lasiodiplodia theobromae and L. pseudotheobromae causing leaf necrosis on Camellia sinensis in Fujian Province, China. Can. J. Plant Pathol. 2019, 41, 277–284. [Google Scholar] [CrossRef]
- Naidoo, K.K.; Coopoosamy, R.M.; Naidoo, G. Screening of Chromolaeana odorata (L.) King and Robinson for antibacterial and antifungal properties. J. Med. Plant Res. 2011, 5, 4859–4862. [Google Scholar]
- Okoro, A.; Onaebi, C. Sensitivity assessment of soil-borne pathogens to ethanolic and methanolic leaf extract of Chromolaena odorata (L.) using dilution and diffusion assay methods. J. Phytopathol. 2020, 32, 71–78. [Google Scholar] [CrossRef]
- Meela, M.M.; Mdee, L.K.; Masoko, P.; Eloff, J.N. Acetone leaf extracts of seven invasive weeds have promising activity against eight important plant fungal pathogens. S. Afr. J. Bot. 2019, 121, 442–446. [Google Scholar] [CrossRef]
- Manjappa, K. Evaluation of antifungal properties of Eupatorium (Chromolaena odorata L.) plant extract against Pyricularia oryzae causing blast disease in rice crop. Asian J. Pharm. Sci. Technol. 2015, 5, 79–81. [Google Scholar]
- Wang, L.; Li, Y.X.; Ge, W.C.; Yuan, K. Chemical composition and antifungal activity of essential oils of Eupatorium catarium and Eupatorium odoratum from China. Asian J. Chem. 2014, 26, 7168–7172. [Google Scholar] [CrossRef]
- Qin, R.M.; Zheng, Y.L.; Valiente-Banuet, A.; Callaway, R.M.; Barclay, G.F.; Silva-Pereyra, C.; Feng, Y.L. The evolution of increased competitive ability, innate competitive advantage, and novel biochemical weapons act in concert to promote invasion for a tropical invader. New Phytol. 2013, 197, 979–988. [Google Scholar] [CrossRef]
- Zheng, Y.; Liao, Z. High-density native-range species affects the invasive plant Chromolaena odorata more strongly than species from its invasive range. Sci. Rep. 2017, 7, 16075. [Google Scholar] [CrossRef] [Green Version]
- Zheng, L.; Burns, J.H.; Liao, Z.Y.; Li, Y.P.; Yang, J.; Chen, Y.J.; Zheng, Y.G.; Zhang, J.I.; Zheng, U.G. Species composition, functional and phylogenetic distances correlate with success of invasive Chromolaena odorata in an experimental test. Ecol. Lett. 2018, 21, 1211–1220. [Google Scholar] [CrossRef]
- Rice, E.L. Allelopathy, 2nd ed.; Academic Press: Orlando, FL, USA, 1984; pp. 1–422. [Google Scholar]
- Bais, H.P.; Weir, T.L.; Perry, L.G.; Gilroy, S.; Vivanco, J.M. The role of root exudates in rhizosphere interactions with plants and other organisms. Annu. Rev. Plant Biol. 2006, 57, 233–266. [Google Scholar] [CrossRef] [Green Version]
- Bonanomi, G.; Sicurezza, M.G.; Caporaso, S.; Esposito, A.; Mazzoleni, S. Phytotoxicity dynamics of decaying plant materials. New Phytol. 2006, 169, 571–578. [Google Scholar] [CrossRef]
- Belz, R.G. Allelopathy in crop/weed interactions—An update. Pest. Manag. Sci. 2007, 63, 308–326. [Google Scholar] [CrossRef]
- Sangakkara, U.R.; Attanayake, K.B.; Dissanayake, U.; Bandaranayake, P.R.S.D. Allelopathic impact of Chromolaena odorata (L.) King and Robinson on germination and growth of selected tropical crops. J. Plant Dis. Proct. 2008, 21, 323–326. [Google Scholar]
- Sahid, I.; Yusoff, N. Allelopathic effects of Chromolaena odorata (L.) King and Robinson and Mikania micrantha H.B.K. on three selected weed species. Aust. J. Crop Sci. 2014, 8, 1024–1028. [Google Scholar]
- Onwugbuta-Enyi, J. Allelopathic effects of Chromolaena odorata L. (RM King and Robinson—(Awolowo Plant)) toxin on tomatoes (Lycopersicum esculentum Mill). J. Appl. Sci. Environ. Manag. 2001, 5, 69–73. [Google Scholar]
- Nakamura, N.; Nemoto, M. Allelopathic potential of Eupatorium odoratum in abandoned shifting cultivation fields in the tropics. Weed Res. 1993, 38, 103–108. [Google Scholar]
- Hu, G.; Zhang, Z. Allelopathic effects of Chromolaena odorata on native and non-native invasive herbs. J. Food Agric. Environ. 2013, 11, 878–882. [Google Scholar]
- Jash, A.; Halder, S.; Bhattacharjee, A. Assessment of allelopathic potential of Chromolaena odorata (L.) King and Robinson by physiobiochemical approach. Life Sci. Infor. Pub. 2019, 5, 2. [Google Scholar]
- Poonpaiboonpipat, T.; Krumsri, R.; Kato-Noguchi, H. Allelopathic and herbicidal effects of crude extract from Chromolaena odorata (L.) R.M. King and H. Rob. on Echinochloa crus-galli and Amaranthus viridis. Plants 2021, 10, 1609. [Google Scholar] [CrossRef]
- Hamidi, F.W.A.; Ismail, A.M.; Zainuddin, F.H.I.; Hasan, M.Y. Preliminary study on allelopathic effect from Chromolaena odarata (siam weed) leaves extract towards Vigna radiata. Int. J. Eng. Res. Technol. 2014, 3, 406–411. [Google Scholar]
- Muzzo, B.I.; Mwilawa, A.J.; Maleko, D.D.; Mtengeti, E.J. Allelopathic effect of Chromolaena odorata aqueous leaf extracts on seed germination and seedling growth of selected crop and pasture species in Tanzania. Int. J. Bot. Stud. 2018, 3, 41–48. [Google Scholar]
- Rusdy, M.; Ako, A. Allelopathic effect of Lantana camara and Chromolaena odorata on germination and seedling growth of Centroma pubescens. Int. J. Appl. Environ. Sci. 2017, 12, 1769–1776. [Google Scholar]
- Yusoff, N.; Ismail, B.S. Allelopathic potential of Chromolaena odorata and Mikania micrantha on Brassica chinensis var. parachinensis. AIP Conf. Proc. 2015, 1678, 020024. [Google Scholar]
- Masum, S.M.; Ali, M.H.; Mandal, M.S.H.; Haque, M.N.; Mahto, A.K. Influence of Parthenium hysterophorus, Chromolaena odorata and PRH on seed germination and seedling growth of maize, soybean and cotton. Bangladesh J. 2012, 3, 83–90. [Google Scholar]
- Pertin, N.; Sutradhar, J.; Das, A.P. Allelopathic effects of two important exotic weeds, Ageratum houstonianum Mill. and Chromolaena odorata (L.) R.M. King & H. Rob. on some crop plants. Pleione 2018, 12, 45–51. [Google Scholar]
- Erida, G.; Saidi, N. Allelopathic screening of several weed species as potential bioherbicides. In IOP Conference Series: Earth and Environmental Science; IOP Publishing: Bristol, UK, 2019; Volume 334, p. 012034. [Google Scholar]
- Madane, A.N.; Patil, B.J. Allelopathic effect of Eupatorium odoratum L. on amylase activity during seed germination of Cicer arietinum L. and Cajanus cajan (L) Millsp. Biosci. Dis. 2017, 8, 82–86. [Google Scholar]
- Yuliyani, E.D.; Darmanti, S.; Hastuti, E.D. Allelochemical effects of Chromolaena odorata L. against photosynthetic pigments and stomata of Ageratum conyzoides L. leaves. J. Phys. Conf. Ser. 2019, 1217, 012149. [Google Scholar] [CrossRef]
- Nandi, A.K.; Mandal, G.D. Allelotoxicity of Chromolaena odorata (L.) King & Robinson on growth, cytology and biochemicals of Allium cepa L. Nat. Environ. Pollut. Technol. 2009, 8, 395–398. [Google Scholar]
- Vartapetian, B.B.; Jackson, M.B. Plant adaptations to anaerobic stress. Ann. Bot. 1997, 79 (Suppl. S1), 3–20. [Google Scholar] [CrossRef]
- Perata, P.; Guglielminetti, L.; Alpi, A. Mobilization of endosperm reserves in cereal seeds under anoxia. Ann. Bot. 1997, 79 (Suppl. S1), 49–56. [Google Scholar] [CrossRef] [Green Version]
- Harborne, J.B.; Turner, B.L. Plant Chemosystematics; Academic Press: London, UK, 1984; pp. 1–562. [Google Scholar]
- Harborne, J.B.; Baxter, H. Phytochemical Dictionary. In A Handbook of Bioactive Compounds from Plants; Taylor & Francis: London, UK, 1993; pp. 1–791. [Google Scholar]
- Jain, D.C.; Tripathi, A.K. Potential of natural products as insect antifeedants. Phytother. Res. 1993, 7, 327–334. [Google Scholar] [CrossRef]
- Wink, M. Evolution of secondary metabolites from an ecological and molecular phylogenetic perspective. Phytochemistry 2003, 64, 3–19. [Google Scholar] [CrossRef] [PubMed]
- Ahuja, I.; Kissen, R.; Bones, A.M. Phytoalexins in defense against pathogens. Trends Plant Sci. 2012, 17, 73–90. [Google Scholar] [CrossRef] [PubMed]
- Skubel, S.A.; Su, X.; Poulev, A.; Foxcroft, L.C.; Dushenkov, V.; Raskin, I. Metabolomic differences between invasive alien plants from native and invaded habitats. Sci. Rep. 2020, 10, 9749. [Google Scholar] [CrossRef] [PubMed]
- Phan, T.T.; Wang, L.; See, P.; Grayer, R.J.; Chan, S.Y.; Lee, S.T. Phenolic compounds of Chromolaena odorata protect cultured skin cells from oxidative damage: Implication for cutaneous wound healing. Biol. Pharm. Bull. 2001, 24, 1373–1379. [Google Scholar] [CrossRef] [Green Version]
- Suksamrarn, A.; Chotipong, A.; Suavansri, T.; Boongird, S.; Timsuksai, P.; Vimuttipong, S.; Chuaynugul, A. Antimycobacterial activity and cytotoxicity of flavonoids from the flowers of Chromolaena odorata. Arch. Pharm. Res. 2004, 27, 507–511. [Google Scholar] [CrossRef] [PubMed]
- Joshi, R.K. Chemical composition of the essential oil of Chromolaena odorata (L.) R.M. King & H. Rob. roots from India. J. Chem. 2013, 2013, 195057. [Google Scholar]
- Liu, P.Y.; Liu, D.; Li, W.H.; Zhao, T.; Sauriol, F.; Gu, Y.C.; Shi, Q.W.; Zhang, M.L. Chemical constituents of plants from the genus Eupatorium (1904–2014). Chem. Biodivers. 2015, 12, 1481–1515. [Google Scholar] [CrossRef]
- Kanase, V.; Shaikh, S. A pharmacognostic and pharmacological review on Chromolaena odorata (Siam Weed). Asian J. Pharm. Clin. Res. 2018, 11, 34–38. [Google Scholar] [CrossRef]
- Abubakar, M.A.; Etonihu, A.C.; Kigbu, P.E.; Owuna, J.E.; Audu, S.I. Phytochemical and antimicrobial analyses of leaf extracts of Cerathoteca sesamoides and Chromolaena odorata. Int. J. Res. 2020, 8, 65–74. [Google Scholar]
- Omokhua-Uyi, A.G.; Abdalla, M.A.; Leonard, C.M.; Aro, A.; Uyi, O.O.; van Staden, J.; McGaw, L.J. Flavonoids isolated from the South African weed Chromolaena odorata (Asteraceae) have pharmacological activity against uropathogens. BMC Complement. Med. Ther. 2020, 20, 233. [Google Scholar] [CrossRef]
- Eze, F.N.; Jayeoye, T.J. Chromolaena odorata (Siam weed): A natural reservoir of bioactive compounds with potent anti-fibrillogenic, antioxidative, and cytocompatible properties. Biomed. Pharmacother. 2021, 141, 111811. [Google Scholar] [CrossRef] [PubMed]
- Olawale, F.; Olofinsan, K.; Iwaloye, O. Biological activities of Chromolaena odorata: A mechanistic review. S. Afr. J. Bot. 2022, 144, 44–57. [Google Scholar] [CrossRef]
- Li, Z.H.; Wang, Q.; Ruan, X.; Pan, C.D.; Jiang, D.A. Phenolics and plant allelopathy. Molecules 2010, 15, 8933–8952. [Google Scholar] [CrossRef] [Green Version]
- Widhalm, J.R.; Dudareva, N. A familiar ring to it: Biosynthesis of plant benzoic acids. Mol. Plant 2015, 8, 83–97. [Google Scholar] [CrossRef] [Green Version]
- Dalton, B.R. The occurrence and behavior of plant phenolic acids in soil environments and their potential involvement in allelochemical interference interactions: Methodological limitations in establishing conclusive proof of allelopathy. In Principals and Practices in Plant Ecology: Allelochemical Interactions; Inderjit, Dakshini, K.M.M., Foy, C.L., Eds.; CRC Press: Boca Raton, FL, USA, 1999; pp. 57–74. [Google Scholar]
- Inderjit. Plant phenolics in allelopathy. Bot. Rev. 1996, 62, 186–202.
- Einhellig, F.A. Mode of Action of Allelochemical Action of Phenolic Compounds. In Chemistry and Mode of Action of Allelochemicals; Macías, F.A., Galindo, J.C.G., Molino, J.M.G., Cutler, H.G., Eds.; CRC Press: Boca Raton, FL, USA; London, UK; New York, NY, USA; Washington, DC, USA, 2004; pp. 217–238. [Google Scholar]
- Pitakpawasutthi, Y.; Thitikornpong, W.; Palanuvej, C.; Ruangrungsi, N. Chlorogenic acid content, essential oil compositions, and in vitro antioxidant activities of Chromolaena odorata leaves. J. Adv. Pharm. Technol. Res. 2016, 7, 37–42. [Google Scholar]
- Nakai, S.; Inoue, Y.; Hosomi, M.; Murakami, A. Myriophyllum spicatum-released allelopathic polyphenols inhibiting growth of blue-green algae Microcystis aeruginosa. Water Res. 2000, 34, 3026–3032. [Google Scholar] [CrossRef]
- Treutter, D. Significance of flavonoids in plant resistance: A review. Environ. Chem. Lett. 2006, 4, 147. [Google Scholar] [CrossRef]
- Weston, L.A.; Mathesius, U. Flavonoids: Their structure, biosynthesis and role in the rhizosphere, including allelopathy. J. Chem. Ecol. 2013, 39, 283–297. [Google Scholar] [CrossRef]
- Mierziak, J.; Kostyn, K.; Kulma, A. Flavonoids as important molecules of plant interactions with the environment. Molecules 2014, 19, 16240–16265. [Google Scholar] [CrossRef] [Green Version]
- Ohtsuki, T.; Hiraka, T.; Kikuchi, H. Flavonoids from Eupatorium odoratum with death receptor 5 promoter enhancing activity. Heterocycles 2009, 77, 1379–1388. [Google Scholar]
- Kodama, O.; Miyakawa, J.; Akatsuka, T.; Kiyosawa, S. Sakuranetin, a flavanone phytoalexin from ultraviolet-irradiated rice leaves. Phytochemistry 1992, 31, 3807–3809. [Google Scholar] [CrossRef]
- Stompor, M. A review on sources and pharmacological aspects of sakuranetin. Nutrients 2020, 12, 513. [Google Scholar] [CrossRef] [Green Version]
- Wollenweber, E.; Dörr, M.; Muniappan, R. Exudate flavonoids in a tropical weed, Chromolaena odorata (L.) R.M. King et H. Robinson. Biochem. Syst. Ecol. 1995, 23, 873–874. [Google Scholar] [CrossRef]
- Parvez, M.M.K.; Yokotani, T.; Fujii, Y.; Konishi, T.; Iwashina, T. Effects of quercetin and its seven derivatives on the growth of Arabidopsis thaliana and Neurospora crassa. Biochem. Syst. Ecol. 2004, 32, 631–635. [Google Scholar] [CrossRef]
- Okada, S.; Iwasaki, A.; Kataoka, I.; Suenaga, K.; Kato-Noguchi, H. Isolation and identification of a phytotoxic substance in kiwifruit leaves. Acta Hortic. 2018, 1218, 207–212. [Google Scholar] [CrossRef]
- Moreland, D.E.; Novitzky, W.P. Effects of Phenolic Acids, Coumarins, and Flavonoids on Isolated Chloroplasts and Mitochondria. In Allelochemicals: Role in Agriculture and Forestry; Waller, E.D., Ed.; ACS publications: Washington, DC, USA, 1987; Series 330; pp. 247–274. [Google Scholar]
- Einhellig, F.A. Mechanisms of Action of Allelochemicals in Allelopathy. In Allelopathy; Inderjit, Dakshini, K.M.M., Einhellig, F.A., Eds.; ACS publications: Washington, DC, USA, 1995; Series 582; pp. 96–116. [Google Scholar]
- Li, J.; Cao, L.; Guo, Z.; An, G.; Li, B.; Li, J. Time- and dose-dependent allelopathic effects and mechanisms of kaempferol on toxigenic Microcystis growth. Ecotoxicol. Environ. Saf. 2021, 222, 112508. [Google Scholar] [CrossRef]
- Gershenzon, J.; Dudareva, N. The function of terpene natural products in the natural world. Nat. Chem. Biol. 2007, 3, 408–414. [Google Scholar] [CrossRef]
- M’sou, S.; Alifriqui, M.; Romane, A. Phytochemical study and biological effects of the essential oil of Fraxinus dimorpha Coss & Durieu. Nat. Prod. Res. 2017, 31, 2797–2800. [Google Scholar]
- Riccobono, L.; Maggio, A.; Bruno, M.; Spadaro, V.; Raimondo, F.M. Chemical composition and antimicrobial activity of the essential oils of some species of Anthemis sect. Anthemis (Asteraceae) from Sicily. Nat. Prod. Res. 2017, 31, 2759–2767. [Google Scholar] [CrossRef]
- Winters, A.E.; White, A.M.; Dewi, A.S.; Mudianta, I.W.; Wilson, N.G.; Forster, L.C.; Garson, M.J.; Cheney, K.L. Distribution of defensive metabolites in nudibranchmolluscs. J. Chem. Ecol. 2018, 44, 384–396. [Google Scholar] [CrossRef]
- Sarles, L.; Boullis, A.; Fassotte, B.; Lognay, G.; Verhaeghe, A.; Francis, F.; Verheggen, F.J. Identification of walnut husk (Juglans regia L.) volatiles and the behavioural response of the invasive walnut husk fly, Rhagoletis completa Cresson. Pest. Manag. Sci. 2017, 73, 2100–2104. [Google Scholar] [CrossRef]
- Boncan, D.A.T.; Tsang, S.S.K.; Li, C.; Lee, I.H.T.; Lam, H.M.; Chan, T.F.; Hui, J.H.L. Terpenes and Terpenoids in Plants: Interactions with Environment and Insects. Int. J. Mol. Sci. 2020, 21, 7382. [Google Scholar] [CrossRef]
- Angelini, L.G.; Carpanese, G.; Cioni, P.L.; Morelli, I.; Macchia, M.; Flamini, G. Essential oils from Mediterranean Lamiaceae as weed germination inhibitors. J. Agric. Food. Chem. 2003, 51, 6158–6164. [Google Scholar] [CrossRef]
- Al-Rowaily, S.L.; Abd-ElGawad, A.M.; Assaeed, A.M.; Elgamal, A.M.; Gendy, A.E.N.G.E.; Mohamed, T.A.; Dar, B.A.; Mohamed, T.K.; Elshamy, A.I. Essential Oil of Calotropis procera: Comparative Chemical Profiles, Antimicrobial Activity, and Allelopathic Potential on Weeds. Molecules 2020, 25, 5203. [Google Scholar] [CrossRef]
- Gharibvandi, A.; Karimmojeni, H.; Ehsanzadeh, P.; Rahimmalek, M.; Mastinu, A. Weed management by allelopathic activity of Foeniculum vulgare essential oil. Plant Biosyst. 2022, 156, 1298–1306. [Google Scholar] [CrossRef]
- Zhang, J.; An, M.; Wu, H.; Liu, D.L.; Stanton, R. Chemical composition of essential oils of four Eucalyptus species and their phytotoxicity on silverleaf nightshade (Solanum elaeagnifolium Cav.) in Australia. Plant Growth Regul. 2012, 68, 231–237. [Google Scholar] [CrossRef]
- Romagni, J.G.; Allen, S.N.; Dayan, F.E. Allelopathic effects of volatile cineoles on two weedy plant species. J. Chem. Ecol. 2000, 26, 303–313. [Google Scholar] [CrossRef]
Figure 1.
C. odorata; photos were kindly provided by Dr. Poonpaiboonpipat, T.

Figure 2.
Pyrrolizidine alkaloids; 1: 7-angeloylretronecine, 2: 9-angeloylretronecine, 3: 3′-acetylrinderine, 4; rinderine, 5: intermedine, 6: rinderine N-oxide, 7: intermedine N-oxide, 8: 1,2-dehydropyrrolizine alkaloid.
Figure 2.
Pyrrolizidine alkaloids; 1: 7-angeloylretronecine, 2: 9-angeloylretronecine, 3: 3′-acetylrinderine, 4; rinderine, 5: intermedine, 6: rinderine N-oxide, 7: intermedine N-oxide, 8: 1,2-dehydropyrrolizine alkaloid.

Figure 3.
Flavonoids; 9: dihydrokaempferol-3-methoxy ether, 10: isosakuranetin, 11: kaempferide-4’-methoxy ether, 12: 3,5-dihy-droxy-7,4’-dimethoxyflavone, 13: acutellerin-4’,6,7-trimethy ether, 14: 4’,5,6,7-tetramethoxyflavone, 15: odoratin. The concentration of these compounds was grater in C. odorata collected from the invasive ranges than that from the native ranges.
Figure 3.
Flavonoids; 9: dihydrokaempferol-3-methoxy ether, 10: isosakuranetin, 11: kaempferide-4’-methoxy ether, 12: 3,5-dihy-droxy-7,4’-dimethoxyflavone, 13: acutellerin-4’,6,7-trimethy ether, 14: 4’,5,6,7-tetramethoxyflavone, 15: odoratin. The concentration of these compounds was grater in C. odorata collected from the invasive ranges than that from the native ranges.

Figure 4.
Potential allelochemicals; 16: p-hydroxybenzoic acid, 17: protocatechuic acid, 18: p-coumaric acid, 19: sakuranetin, 20: quercetin, 21: kaempferol, 22: α-pinene, 23: 1,8-cineole.
Figure 4.
Potential allelochemicals; 16: p-hydroxybenzoic acid, 17: protocatechuic acid, 18: p-coumaric acid, 19: sakuranetin, 20: quercetin, 21: kaempferol, 22: α-pinene, 23: 1,8-cineole.


{kind=link}
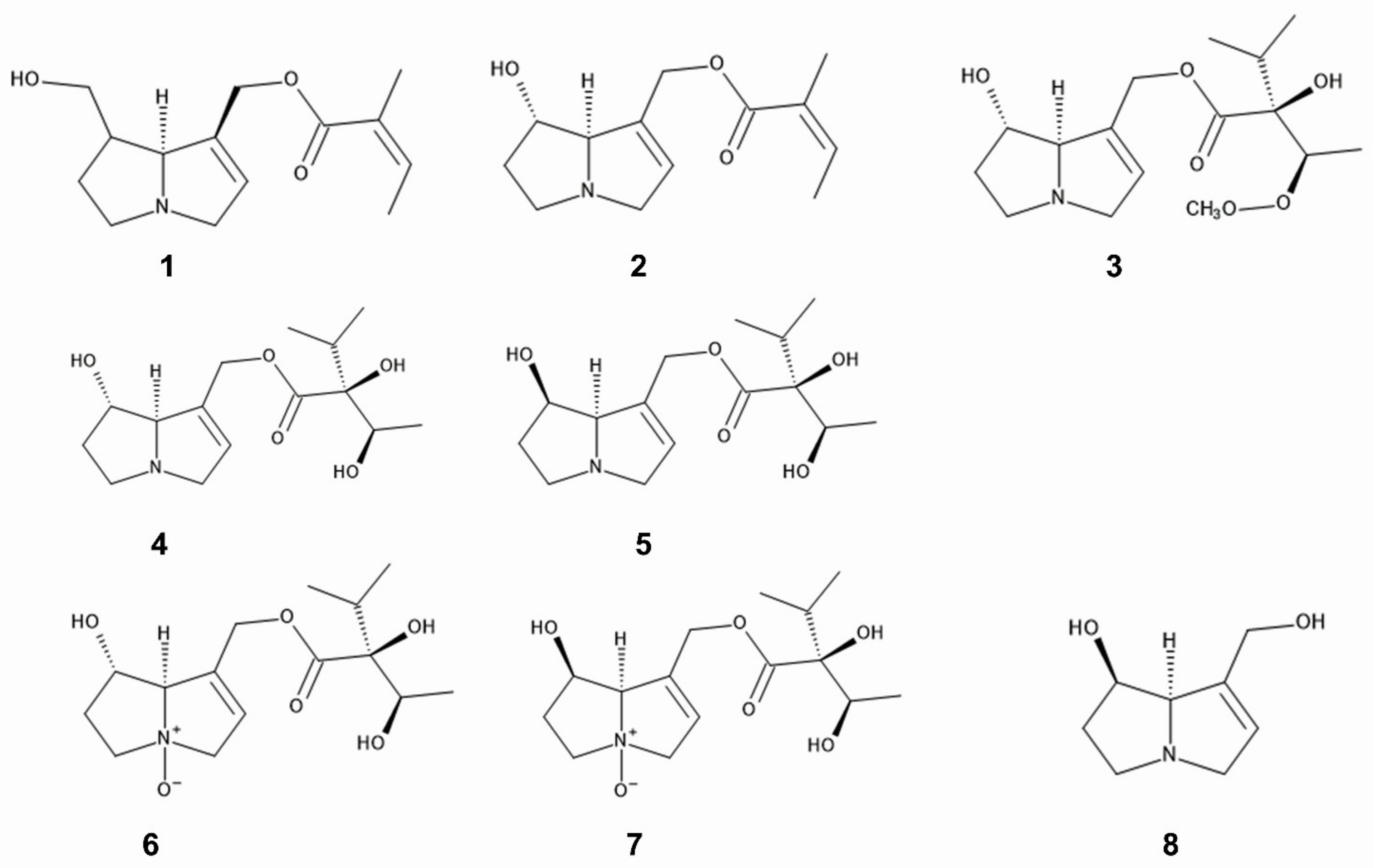{kind=link}
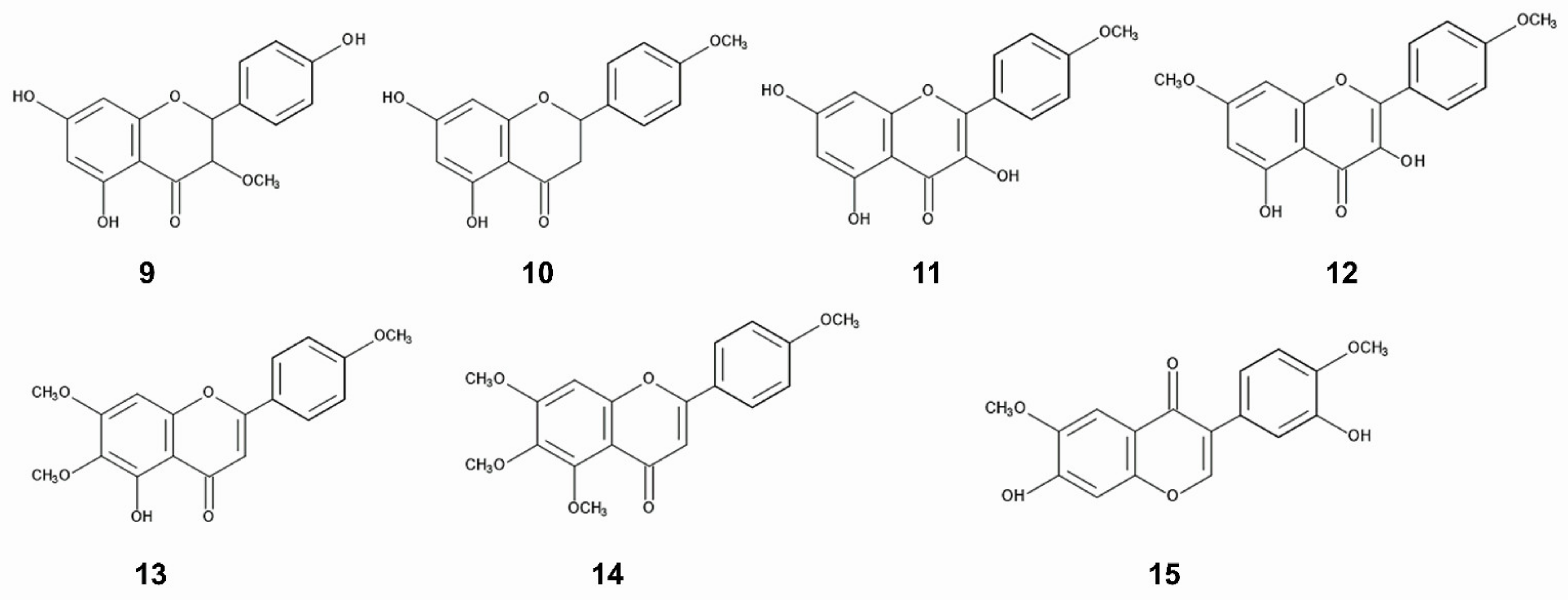{kind=link}
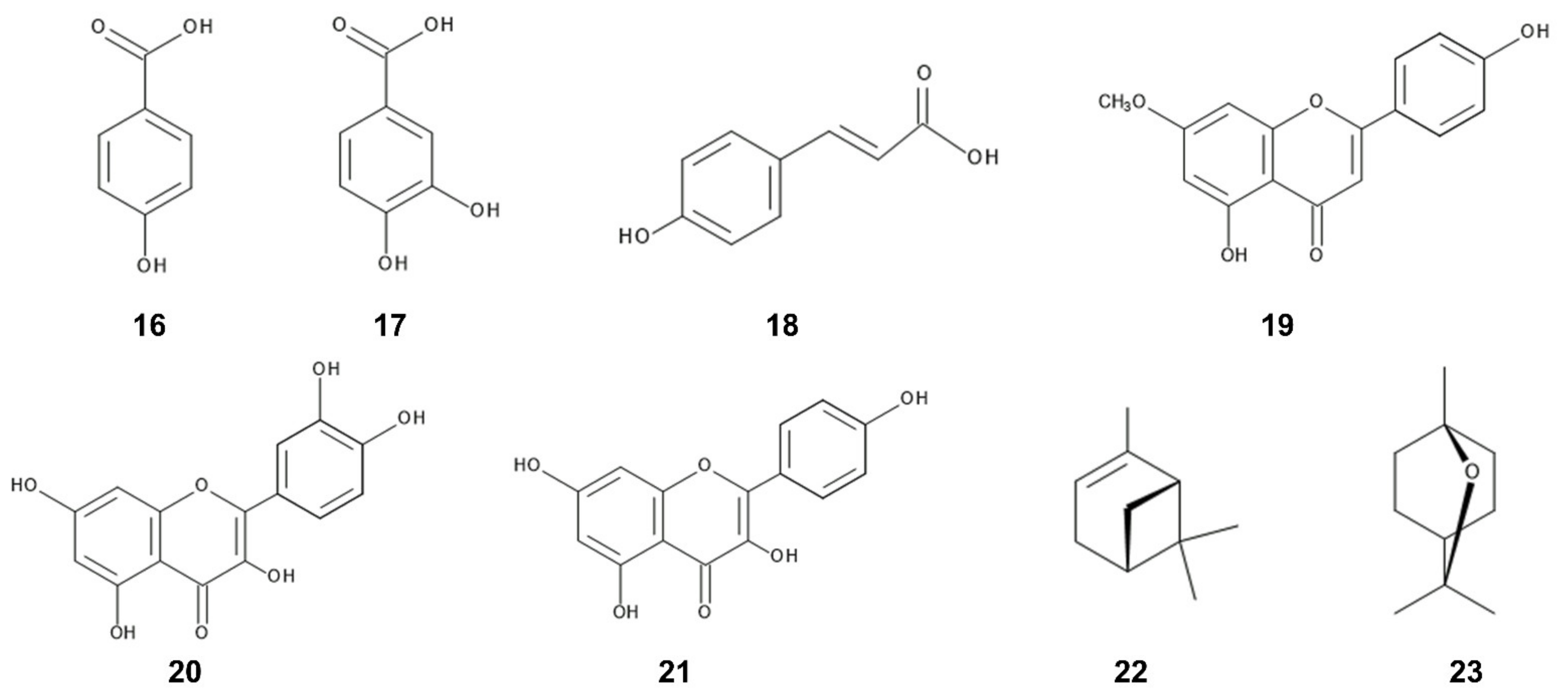{kind=link}
Table 1.
Interaction of C. odorata with insects.
| Source | Insect | Action | Reference |
|---|---|---|---|
| Root powder | Callosobruchus maculatus | Mortality (100%) at 8% (weight of powder/weight of crop grain) | [65] |
| Leaf powder | Callosobruchus maculatus | Mortality (51%) at 1.7% (weight of powder/weight of crop grain) | [66] |
| Essential oil from leaves | Sitophilus zeamais | Mortality (100%) at 0.5% (volume of essential oil/weight of crop grain) | [67] |
| Leaf extract | Sitophilus zeamais | Mortality (50%) at 40 μg/mL methanol extract | [68] |
| Simulium spp. | Mortality (100%) at 1 mg dry mass of aqueous extract/mL | [69] | |
| Periplaneta americana | Mortality (36%) at 1 mL dose of 20% (w/v) aqueous extract | [70] | |
| Bemissa tabaci, Amrasca biguttula | Population 25–50% decrease of control at 2.5% (w/v) ethanol extract | [71] | |
| Spodoptera litura | Mortality 68% at 5% (w/v) methanol extract | [72] |
Table 2.
Interaction of C. odorata with nematodes.
| Source | Nematode | Action | Reference |
|---|---|---|---|
| Plant infestation soil | Meloidogyne spp., Helicotylenchus spp., Pratylenchus spp., | Population decrease by 67–79% of control after 30 months | [89] |
| Meloidogyne spp., Helicotylenchus spp., Pratylenchus spp., | Population decrease by 77–81% after 24 months | [90] | |
| Meloidogyne incoginita | Population decrease by 92% after 2 months | [91] | |
| Plant residues | Meloidogyne incoginita | Population decrease significantly at 1% (weight of powder/weight of soil) | [92] |
| Root extract | Meloidogyne incoginita | Parasitism suppression by 0.4–65% of control at 0.1–5% (w/v) aqueous extract | [93] |
| Root mulch | Meloidogyne incoginita | Parasitism suppression by 0–22% at 0.1–5% (weight of plant/weight of soil) | [93] |
Table 3.
Interaction of C. odorata with microbial population.
| Source | Species | Population | Reference |
|---|---|---|---|
| Plant infestation soil | Paraglomus spp. | Increase biomass significantly after 10 months incubation | [96] |
| Rhizosphere soil | Fusarium ssp. | Increase spore number 25 times after 5 days incubation | [103] |
| Leaf extract | Lasiodiplodia theobromae, Lasiodiplodia pseudothobromae | Decrease biomass by 6–17% of control at 3% (w/v; extract residue/medium) | [106] |
| Bacillus subtilis, Bacillus cereus | Decrease population significantly at 10% (w/v; plant weight/solvent) | [109] | |
| Phytophthora colocasiae, Fusarium oxysporum | Decrease of the colony expansion by 20–33% of control at 100 mg leaves/mL methanol | [110] | |
| Pythium ultimum, Rhizoctonia solani, Fusarium oxysporium, Phytophthora nicotianae | Inhibitory activity at concentration of 0.09 mg residue of extract/mL of medium | [111] | |
| Pyricularia oryzae | Decrease by 12–43% of control at 10% (w/v; extract residue/solvent) | [112] |
Table 4.
Allelopathic activities of C. odorata.
| Source | Inhibition | Target Plant Species | Reference | |||
|---|---|---|---|---|---|---|
| Germination | Growth | Chlorophyll | Amylase | |||
| Plan residue | ✔ | Capsicum annuum, Solanum melongena | [121] | |||
| ✔ | ✔ | Eleusine indica, Cyperus iria, Ageratum conyzoides | [122] | |||
| ✔ | Lycopersicon esculentum | [123] | ||||
| Leaf powder | ✔ | Crassocephalum crepidioides | [124] | |||
| Extract | ||||||
| Leaf | ✔ | Eupatorium japonicum, Eupatorium stoechadosmum, Eupatorium lindleyanum | [114] | |||
| ✔ | ✔ | Lathyrus sativus | [126] | |||
| ✔ | ✔ | Cynodon dactylon, Crassocephalum crepidioides, Ageratum conyzoides | [124] | |||
| ✔ | ✔ | Eleusine indica, Cyperus iria, Ageratum conyzoides | [122] | |||
| ✔ | ✔ | Sorghum bicolor, Zea mays, Phaseolus vulgaris, Vigna radiata, Centrosema pubescens | [128,129,130] | |||
| ✔ | ✔ | Brassica chinensis | [131] | |||
| ✔ | ✔ | Glycine max, Gossypium hirsutum | [132] | |||
| ✔ | Lycopersicum esculentum | [123] | ||||
| ✔ | ✔ | Amaranthus spinosus, Amaranthus spinosus | [134] | |||
| ✔ | Cicer arietinum, Cajanus cajan | [135] | ||||
| ✔ | Ageratum conyzoides | [136] | ||||
| ✔ | Allium cepa | [137] | ||||
| Leaf, root | ✔ | ✔ | Bidens pilosa, Ageratum conyzoides, Amaranthus spinosus, Conyza sumatrensis, Chenopodium ambrosioides, Rottboellia exaltata, Digitaria sanguinalis, Hemisteptia lyrata, Youngia japonica, Dicliptera chinensis | [125] | ||
| Aerial part | ✔ | ✔ | Sesame indicum, Brassica juncea, Raphenus raphanistrm | [133] | ||
| Whole plant | ✔ | ✔ | ✔ | Echinochloa crus-galli, Amaranthus viridis | [127] | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Kato-Noguchi, H.; Kato, M. Evolution of the Secondary Metabolites in Invasive Plant Species Chromolaena odorata for the Defense and Allelopathic Functions. Plants 2023, 12, 521. https://doi.org/10.3390/plants12030521
AMA Style
Kato-Noguchi H, Kato M. Evolution of the Secondary Metabolites in Invasive Plant Species Chromolaena odorata for the Defense and Allelopathic Functions. Plants. 2023; 12(3):521. https://doi.org/10.3390/plants12030521
Chicago/Turabian StyleKato-Noguchi, Hisashi, and Midori Kato. 2023. "Evolution of the Secondary Metabolites in Invasive Plant Species Chromolaena odorata for the Defense and Allelopathic Functions" Plants 12, no. 3: 521. https://doi.org/10.3390/plants12030521
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.