
Development and Biomechanics of Grewia lasiocarpa E. Mey. Ex Harv. Trichomes Exudate
 ,
,  ,
,  ,
, 

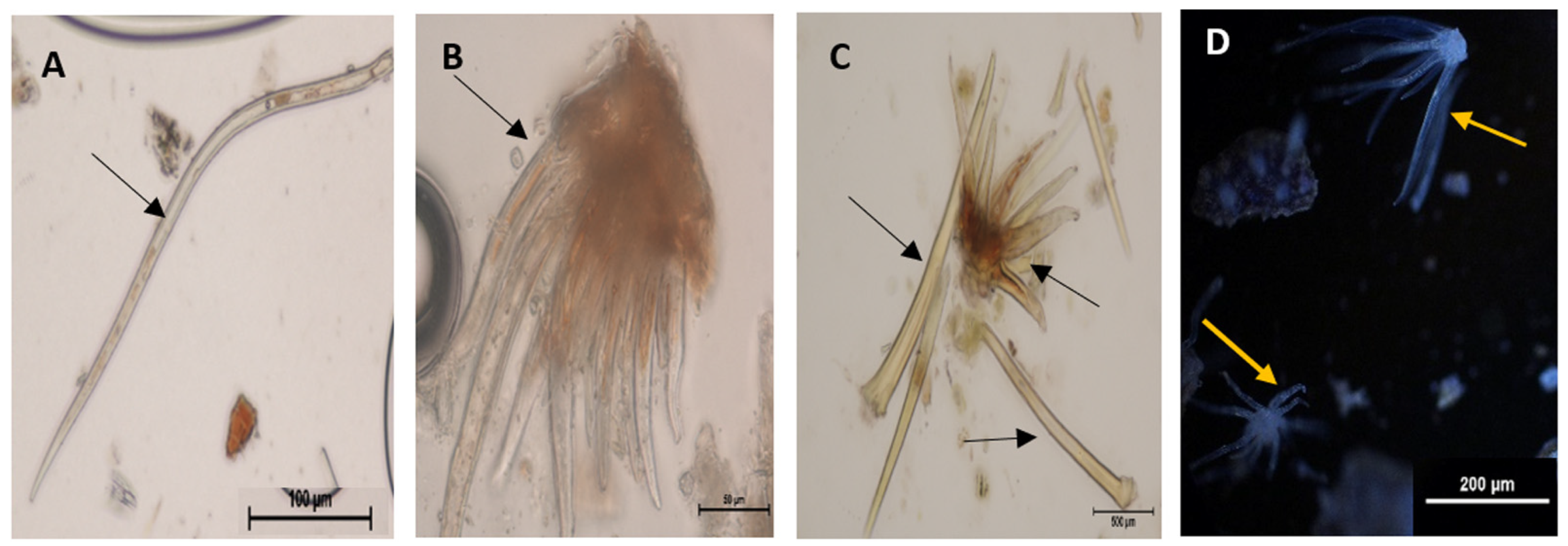{kind=link}
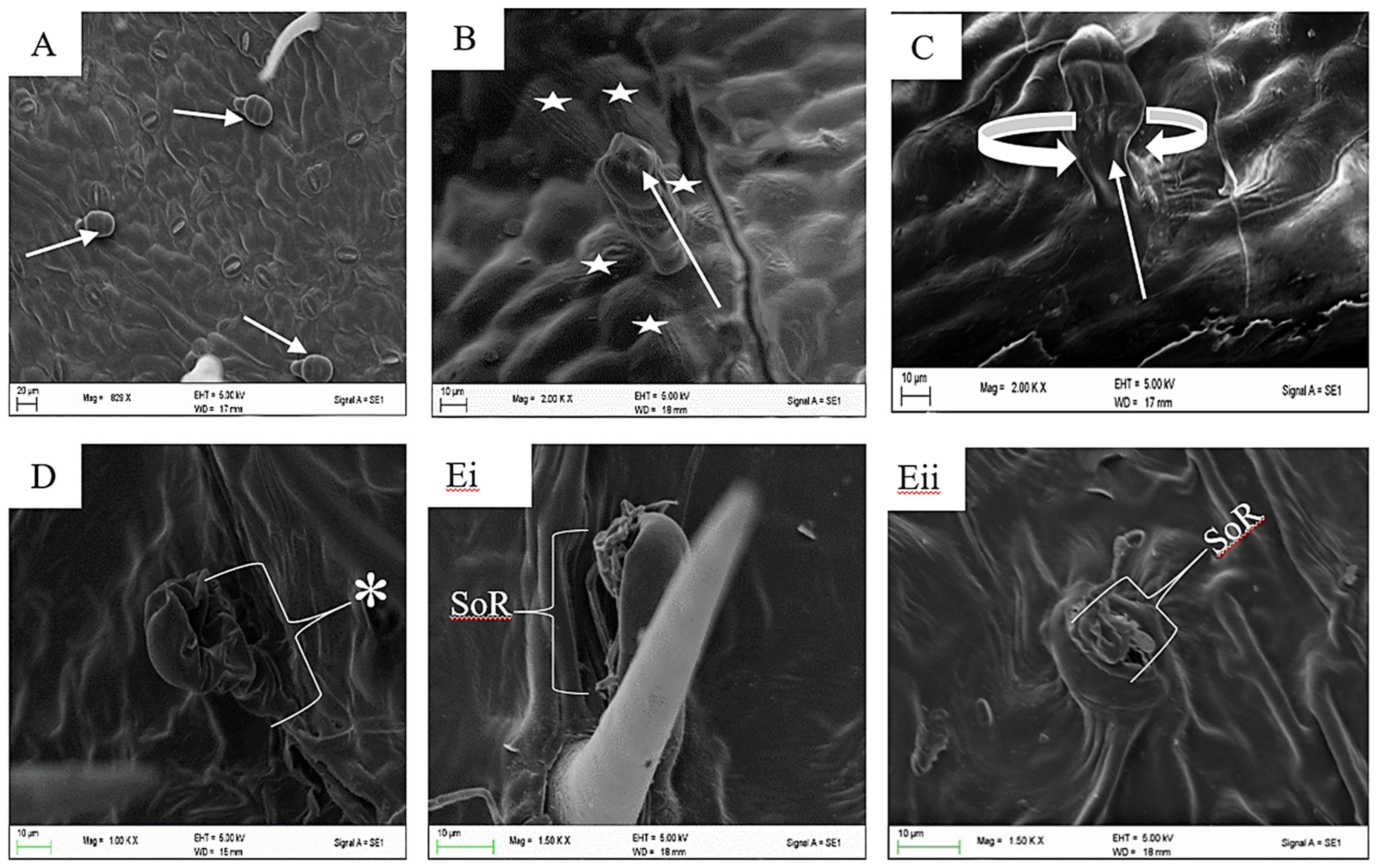{kind=link}
{kind=link}
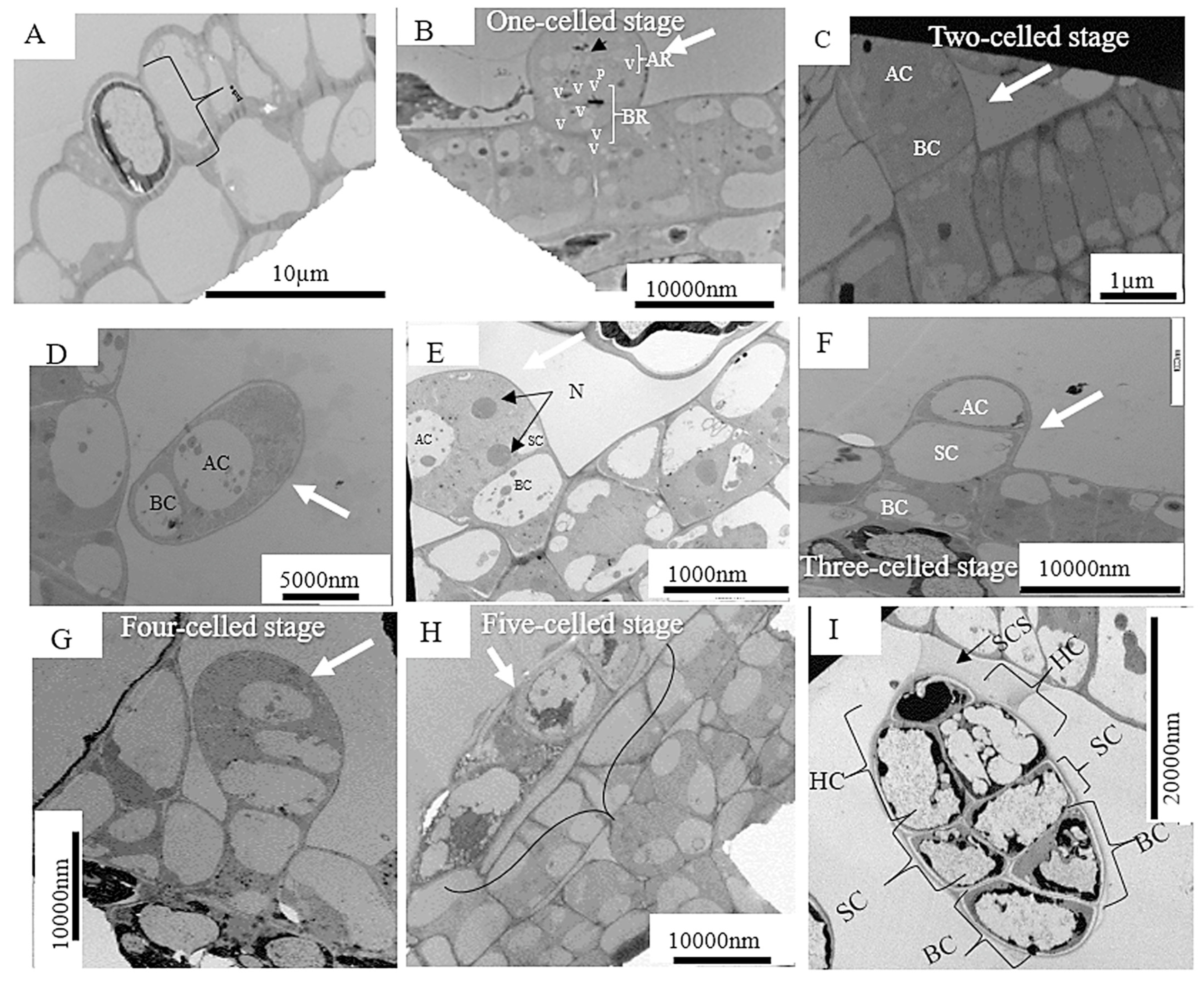{kind=link}
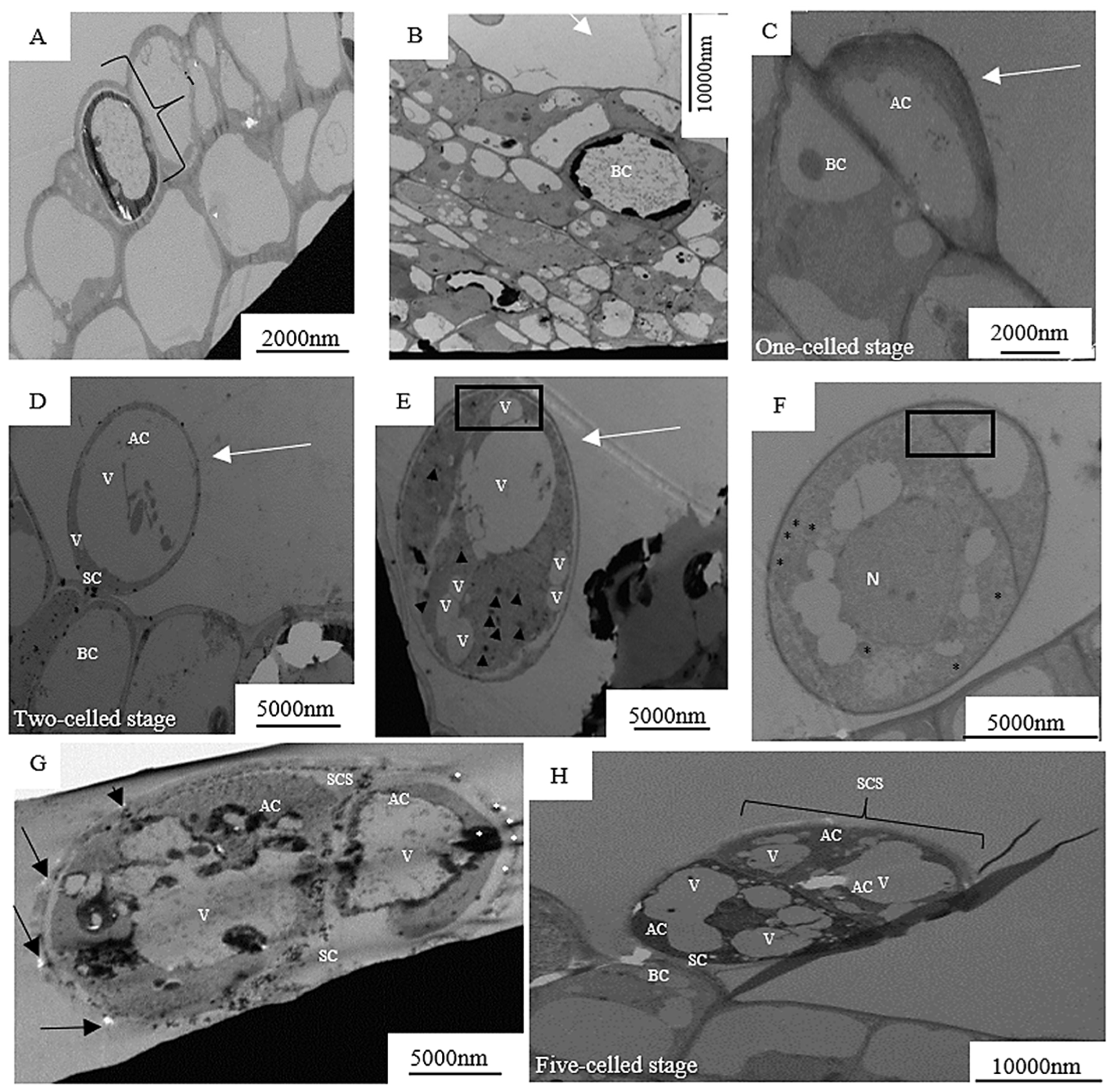{kind=link}
Abstract
:1. Introduction
2. Results
3. Discussion
4. Materials and Methods
4.1. Plant Material
4.2. Sample Preparation for Light Microscopy (LM)
4.3. Sample Preparation for Scanning Electron Microscopy (SEM)
4.4. Sample Preparation for Transmission Electron Microscopy (TEM)
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Chung, R. Revision of Grewia (Malvaceae–Grewioideae) in Peninsular Malaysia and Borneo. Edinb. J. Bot. 2005, 62, 1–27. [Google Scholar] [CrossRef]
- Boon, R. Pooley’s Trees of Eastern South Africa: A Complete Guide; Flora and Fauna Publications Trust: Durban, South Africa, 2010. [Google Scholar]
- Bayer, C.; Kubitzki, K. Malvaceae. In The Families and Genera of Vascular Plants, Malvales, Capparales and Non-Betalain Caryophyllales; Kubitzki, K., Ed.; Springer: Berlin/Heidelberg, Germany, 2003; Volume 5, pp. 225–311. [Google Scholar]
- Wahlert, G.A.; Nusbaumer, L.; Ranirison, P. Grewia gautieri Wahlert & Nusb.(Malvaceae, Grewioideae): A new species from Madagascar. Candollea 2014, 69, 149–155. [Google Scholar]
- Czarnecka, E.; Wiland-Szymanska, J.; Gawronska, K. Phytogeography of the genus Microcos L.(Malvaceae, Grewioidae) in Africa. Biodivers. Res. Conserv. 2006, 34, 269–271. [Google Scholar]
- Akwu, N.; Naidoo, Y.; Singh, M.; Nundkumar, N.; Lin, J. Phytochemical screening, in vitro evaluation of the antimicrobial, antioxidant and cytotoxicity potentials of Grewia lasiocarpa E. Mey. ex Harv. S. Afr. J. Bot. 2019, 123, 180–192. [Google Scholar] [CrossRef]
- Akwu, N.; Naidoo, Y.; Singh, M. A comparative study of the proximate, FTIR analysis and mineral elements of the leaves and stem bark of Grewia lasiocarpa E. Mey. ex Harv.: An indigenous southern African plant. S. Afr. J. Bot. 2019, 123, 9–19. [Google Scholar] [CrossRef]
- Qamar, M.; Akhtar, S.; Barnard, R.T.; Sestili, P.; Ziora, Z.M.; Lazarte, C.E.; Ismail, T. Antiinflammatory and anticancer properties of Grewia asiatica crude extracts and fractions: A bioassay-guided approach. BioMed Res. Int. 2022, 2022, 1–14. [Google Scholar] [CrossRef]
- Bari, W.U.; Zahoor, M.; Zeb, A.; Khan, I.; Nazir, Y.; Khan, A.; Rehman, N.U.; Ullah, R.; Shahat, A.A.; Mahmood, H.M. Anticholinesterase, antioxidant potentials, and molecular docking studies of isolated bioactive compounds from Grewia optiva. Int. J. Food Prop. 2019, 22, 1386–1396. [Google Scholar] [CrossRef]
- Haridas, R.; Sumathi, P.; Thomas, B. Phytochemical investigation by using tender leaf part of Grewia tilifolia VAHL. Kong. Res. J. 2017, 4, 68–70. [Google Scholar] [CrossRef]
- Mongalo, N.; McGaw, L.J.; Finnie, J.F.; Van Staden, J. Isolation and characterization of antimicrobial and anti-inflammatory triterpenoids from the acetone extract of Grewia flava DC.(Malvaceae) roots. S. Afr. J. Bot. 2022, 149, 87–95. [Google Scholar] [CrossRef]
- Nyalo, P.; Omwenga, G.; Ngugi, M. Quantitative Phytochemical Profile and In Vitro Antioxidant Properties of Ethyl Acetate Extracts of Xerophyta spekei (Baker) and Grewia tembensis (Fresen). J. Evid.-Based Integr. Med. 2023, 28, 2515690x231165096. [Google Scholar] [CrossRef]
- Kumar, S.; Singh, B. Traditional uses, phytochemical and pharmacological properties of Grewia sps (Grewia asiatica and Grewia serrulata). In Himalayan Fruits and Berries; Elsevier: Amsterdam, The Netherlands, 2023; pp. 203–219. [Google Scholar]
- Birudu, R.B.; Padmavathi, P. Secondary metabolites of ethanolic leaf extract of Grewia hirsuta. Eur. J. Biomed. 2018, 5, 868–870. [Google Scholar]
- Levin, D. The role of trichomes in plant defense Qrtly. Rev. Biol. 1973, 48, 15. [Google Scholar] [CrossRef]
- Werker, E. Trichome Diversity and Development. In Advances in Botanical Research; Academic Press: NewYork, NY, USA, 2000; Volume 31, pp. 1–35. [Google Scholar]
- Chalvin, C.; Drevensek, S.; Dron, M.; Bendahmane, A.; Boualem, A. Genetic control of glandular trichome development. Trends Plant Sci. 2020, 25, 477–487. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Peled-Zehavi, H.; Galili, G. The m6A reader ECT2 post-transcriptionally regulates proteasome activity in Arabidopsis. New Phytol. 2020, 228, 151–162. [Google Scholar] [CrossRef]
- Akwu, N.A.; Naidoo, Y.; Singh, M. Die anatomie en histochemie van Grewia lasiocarpa E. Mey. ex Harv.(Malvaceae). Suid-Afrik. Tydskr. Nat. Tegnol. 2020, 39, 91–107. [Google Scholar] [CrossRef]
- Xue, S.; Dong, M.; Liu, X.; Xu, S.; Pang, J.; Zhang, W.; Weng, Y.; Ren, H. Classification of fruit trichomes in cucumber and effects of plant hormones on type II fruit trichome development. Planta 2019, 249, 407–416. [Google Scholar] [CrossRef]
- Farag, R.S.; Abdel-Latif, M.S.; Emam, S.; Tawfeek, L.S. Phytochemical screening and polyphenol constituents of pomegranate peels and leave juices. Agric. Soil Sci. 2014, 1, 86–93. [Google Scholar]
- Mohanta, B.; Chakraborty, A.; Sudarshan, M.; Dutta, R.K.; Baruah, M. Elemental profile in some common medicinal plants of India. Its correlation with traditional therapeutic usage. J. Radioanal. Nucl. Chem. 2003, 258, 175–179. [Google Scholar] [CrossRef]
- Randrianasolo, A.; Lowry, P.P.; Schatz, G.E.; Phillipson, P.B.; Wahlert, G.A. The lianescent species of Grewia L.(Malvaceae sl, formerly Tiliaceae) in Madagascar. Adansonia 2013, 35, 73–85. [Google Scholar] [CrossRef]
- Schuurink, R.; Tissier, A. Glandular trichomes: Micro-organs with model status? New Phytol. 2020, 225, 2251–2266. [Google Scholar] [CrossRef]
- Ilham, M.; Mukarromah, S.R.; Rakashiwi, G.A.; Indriati, D.T.; Yoku, B.F.; Purnama, P.R.; Junairiah, J.; Sehanat, P.; Purnobasuki, H.; Wahyuni, D. Morpho-anatomical characterization and DNA barcoding of Achillea millefolium L. Biodivers. J. Biol. Divers. 2022, 23, 1958–1969. [Google Scholar] [CrossRef]
- Tissier, A. Glandular trichomes: What comes after expressed sequence tags? Plant J. 2012, 70, 51–68. [Google Scholar] [CrossRef] [PubMed]
- Figueiredo, A.C.; Barroso, J.G.; Pedro, L.G.; Scheffer, J.J. Factors affecting secondary metabolite production in plants:volatile components and essential oils. Flavour Fragr. J. 2008, 23, 213–226. [Google Scholar] [CrossRef]
- Meer, S.D.; Naidoo, Y.; Dewir, Y.H.; Akwu, N.A.; Fuller, M.P. Foliar microstructure and histochemical analysis of the lavender tree (Heteropyxis natalensis Harv.). Micron 2022, 153, 103184. [Google Scholar] [CrossRef] [PubMed]
- Parusnath, M.; Naidoo, Y.; Akwu, N. The foliar trichomes of Combretum apiculatum subsp. apiculatum (Combretaceae): Morphoanatomy and histochemistry. S. Afr. J. Bot. 2022, 150, 528–547. [Google Scholar] [CrossRef]
- Serna, L.; Martin, C. Trichomes: Different regulatory networks lead to convergent structures. Trends Plant Sci. 2006, 11, 274–280. [Google Scholar] [CrossRef]
- Machado, A.; Wu, Y.; Yang, Y.; Llewellyn, D.J.; Dennis, E.S. The MYB transcription factor GhMYB25 regulates early fibre and trichome development. Plant J. 2009, 59, 52–62. [Google Scholar] [CrossRef]
- Hülskamp, M. Plant trichomes: A model for cell differentiation. Nat. Rev. Mol. Cell Biol. 2004, 5, 471–480. [Google Scholar] [CrossRef]
- Song, S.; Liu, B.; Song, J.; Pang, S.; Song, T.; Gao, S.; Zhang, Y.; Huang, H.; Qi, T. A molecular framework for signaling crosstalk between jasmonate and ethylene in anthocyanin biosynthesis, trichome development, and defenses against insect herbivores in Arabidopsis. J. Integr. Plant Biol. 2022, 64, 1770–1788. [Google Scholar] [CrossRef]
- Wei, Z.; Cheng, Y.; Zhou, C.; Li, D.; Gao, X.; Zhang, S.; Chen, M. Genome-wide identification of direct targets of the TTG1–bHLH–MYB complex in regulating trichome formation and flavonoid accumulation in Arabidopsis thaliana. Int. J. Mol. Sci. 2019, 20, 5014. [Google Scholar] [CrossRef]
- Zeng, J.; Yang, L.; Tian, M.; Xie, X.; Liu, C.; Ruan, Y. SDG26 is involved in trichome control in Arabidopsis thaliana: Affecting phytohormones and adjusting accumulation of H3K27me3 on genes related to trichome growth and Development. Plants 2023, 12, 1651. [Google Scholar] [CrossRef] [PubMed]
- Wagner, G.; Wang, E.; Shepherd, R. New approaches for studying and exploiting an old protuberance, the plant trichome. Ann. Bot. 2004, 93, 3. [Google Scholar] [CrossRef] [PubMed]
- Punja, Z.K.; Sutton, D.B.; Kim, T. Glandular trichome development, morphology, and maturation are influenced by plant age and genotype in high THC-containing cannabis (Cannabis sativa L.) inflorescences. J. Cannabis Res. 2023, 5, 12. [Google Scholar] [CrossRef] [PubMed]
- Nandi, D.; Chakraborty, A.; Biswas, T.; Meher, D.; Singh, A.P. Role of trichomes in plant defence-A crop specific review. Crop Res. 2022, 57, 460–475. [Google Scholar] [CrossRef]
- de Souza Marinke, L.; de Resende, J.T.V.; Hata, F.T.; Dias, D.M.; de Oliveira, L.V.B.; Ventura, M.U.; Zanin, D.S.; de Lima Filho, R.B. Selection of tomato genotypes with high resistance to Tetranychus evansi mediated by glandular trichomes. Phytoparasitica 2022, 50, 629–643. [Google Scholar] [CrossRef]
- Berhin, A.; de Bellis, D.; Franke, R.; Buono, R.; Nowack, M.; Nawrath, C. The root cap cuticle: A cell wall structure for seedling establishment and lateral root formation. Cell 2019, 176, 1367–1378. [Google Scholar] [CrossRef]
- Jia, W.; Wang, Y.; Qi, Q.; He, S.; Mi, Z.; Zhu, X. Leaf epidermal morphology of ten wild tree Peonies in China and its taxonomic significance. Horticulturae 2022, 8, 502. [Google Scholar] [CrossRef]
- Schwab, B.; Folkers, U.; Ilgenfritz, H.; Hülskamp, M. Trichome morphogenesis in Arabidopsis. Philos. Trans. R. Soc. Lond. Ser. B Biol. Sci. 2000, 355, 879–883. [Google Scholar] [CrossRef]
- de Andrade Wagner, M.; Loeuille, B.F.P.; Siniscalchi, C.M.; Melo-de-Pinna, G.F.; Pirani, J.R. Diversity of non-glandular trichomes in subtribe Lychnophorinae (Asteraceae: Vernonieae) and taxonomic implications. Plant Syst. Evol. 2014, 300, 1219–1233. [Google Scholar] [CrossRef]
- Fahn, A. Structure and function of secretory cells. In Advances in Botanical Research. Plant Trichomes; Hallahan, D.L., Gray, J.C., Eds.; Academic Press: Cambridge, MA, USA, 2000; Volume 31, pp. 37–75. [Google Scholar]
- Yang, C.; Ye, Z. Trichomes as models for studying plant cell differentiation. Cell. Mol. Life Sci. 2013, 70, 1937–1948. [Google Scholar] [CrossRef]
- Konarska, A.; Łotocka, B. Glandular trichomes of Robinia viscosa Vent. var. hartwigii (Koehne) Ashe (Faboideae, Fabaceae)—Morphology, histochemistry and ultrastructure. Planta 2020, 252, 102. [Google Scholar] [CrossRef] [PubMed]
- Koudounas, K.; Manioudaki, M.E.; Kourti, A.; Banilas, G.; Hatzopoulos, P. Transcriptional profiling unravels potential metabolic activities of the olive leaf non-glandular trichome. Front. Plant Sci. 2015, 6, 633. [Google Scholar] [CrossRef] [PubMed]
- Stratmann, J.W.; Bequette, C.J. Hairless but no longer clueless: Understanding glandular trichome development. J. Exp. Bot. 2016, 67, 5285–5287. [Google Scholar] [CrossRef] [PubMed]
- Chandra-kuntal, K. Ethylene and ROS Crosstalk in Plant Developmental Processes. In Ethylene in Plant Biology, 1st ed.; Singh, S., Husain, T., Singh, V.P., Tripathi, D.K., Prasad, S.M., Dubey, N.K., Eds.; John Wiley & Sons Ltd., Wiley: Hoboken, NJ, USA, 2023; pp. 125–177. [Google Scholar]
- Autran, D.; Bassel, G.W.; Chae, E.; Ezer, D.; Ferjani, A.; Fleck, C.; Hamant, O.; Hartmann, F.P.; Jiao, Y.; Johnston, I.G. What is quantitative plant biology? Quant. Plant Biol. 2021, 2, e10. [Google Scholar] [CrossRef]
- Yu, D.; Li, X.; Li, Y.; Ali, F.; Li, F.; Wang, Z. Dynamic roles and intricate mechanisms of ethylene in epidermal hair development in Arabidopsis and cotton. New Phytol. 2022, 234, 375–391. [Google Scholar] [CrossRef]
- Abu-Asab, M.S.; Cantino, P.D. Phylogenetic implications of leaf anatomy in subtribe Melittidinae (Labiatae) and related taxa. J. Arnold Arbor. 1987, 68, 1–34. [Google Scholar] [CrossRef]
- Hunsche, M.; Bürling, K.; Saied, A.S.; Schmitz-Eiberger, M.; Sohail, M.; Gebauer, J.; Noga, G.; Buerkert, A. Effects of NaCl on surface properties, chlorophyll fluorescence and light remission, and cellular compounds of Grewia tenax (Forssk.) Fiori and Tamarindus indica L. leaves. Plant Growth Regul. 2010, 61, 253–263. [Google Scholar] [CrossRef]
- Akram, Z.; Perveen, R.; Noreen, A.; Inayat, M.; Hussain, M.; Ibrahim, S.; Razzak, M.; Qadeer, K. Morphological, phytochemical, and pharmacological studies of Grewia asiatica: A review. RADS J. Pharm. Pharm. Sci. 2019, 7, 53–62. [Google Scholar]
- Rajagopal, P.; Vijayan, S.K.; Sreejith, K.; Aneeshia, S.; Premaletha, K. Anti-arthritic and anti-microbial effects of the fruits of Grewia subinaequalis DC. Int. J. Pharma. Res. Health Sci. 2016, 4, 1389–1393. [Google Scholar] [CrossRef]
- Chung, R. Leaf epidermal micromorphology of Grewia L. and Microcos L.(Tiliaceae) in Peninsular Malaysia and Borneo. Gard. Bull. Singap. 2002, 54, 263–286. [Google Scholar]
- Sinha, J.; Purwar, S.; Chuhan, S.K.; Rai, G. Nutritional and medicinal potential of Grewia subinaequalis DC.(syn. G. asiatica.) (Phalsa). J. Med. Plants Res. 2015, 9, 594–612. [Google Scholar]
- Lange, B.M.; Turner, G.W. Terpenoid biosynthesis in trichomes—Current status and future opportunities. Plant Biotechnol. J. 2013, 11, 2–22. [Google Scholar] [CrossRef] [PubMed]
- Werker, E.; Putievsky, E.; Ravid, U.; Dudai, N.; Katzir, I. Glandular hairs, secretory cavities, and the essential oil in the leaves of tarragon (Artemisia dracunculus L.). J. Herbs Spices Med. Plants 1994, 2, 19–32. [Google Scholar] [CrossRef]
- Paetzold, H.; Garms, S.; Bartram, S.; Wieczorek, J.; Urós-Gracia, E.-M.; Rodríguez-Concepción, M.; Boland, W.; Strack, D.; Hause, B.; Walter, M.H. The isogene 1-deoxy-D-xylulose 5-phosphate synthase 2 controls isoprenoid profiles, precursor pathway allocation, and density of tomato trichomes. Mol. Plant 2010, 3, 904–916. [Google Scholar] [CrossRef] [PubMed]
- Kang, J.-H.; McRoberts, J.; Shi, F.; Moreno, J.E.; Jones, A.D.; Howe, G.A. The flavonoid biosynthetic enzyme chalcone isomerase modulates terpenoid production in glandular trichomes of tomato. Plant Physiol. 2014, 164, 1161–1174. [Google Scholar] [CrossRef]
- Xiao, L.; Tan, H.; Zhang, L. Artemisia annua glandular secretory trichomes: The biofactory of antimalarial agent artemisinin. Sci. Bull. 2016, 61, 26–36. [Google Scholar] [CrossRef]
- Raghu, K.; Naidoo, Y.; Dewir, Y. Secretory structures in the leaves of Hibiscus sabdariffa L. S. Afr. J. Bot. 2019, 121, 16–25. [Google Scholar] [CrossRef]
- Lattar, E.C.; Galati, B.G.; Carrera, C.S.; Ferrucci, M.S. Floral nectaries of Heliocarpus popayanensis and Luehea divaricata (Malvaceae-Grewioideae): Structure and ultrastructure. Aust. J. Bot. 2018, 66, 59–73. [Google Scholar] [CrossRef]
- Kumar, V.; Uthappa, A.; Srivastava, M.; Vijay, D.; Kumaranag, K.; Manjunatha, N.; Rana, M.; Newaj, R.; Handa, A.; Chaturvedi, O. Floral biology of Grewia flavescens Juss.: An underutilized crop. Genet. Resour. Crop Evol. 2017, 64, 1789–1795. [Google Scholar] [CrossRef]
- Tominaga-Wada, R.; Ishida, T.; Wada, T. New insights into the mechanism of development of Arabidopsis root hairs and trichomes. Int. Rev. Cell Mol. Biol. 2011, 286, 67–106. [Google Scholar]
- Ishida, T.; Kurata, T.; Okada, K.; Wada, T. A genetic regulatory network in the development of trichomes and root hairs. Ann. Rev. Plant Biol. 2008, 59, 365–386. [Google Scholar] [CrossRef] [PubMed]
- Roland, J.; Vian, B. General preparation and staining of thin sections. In Electron Microscopy of Plant Cells; Hall, J.L., Hawes, C., Eds.; Academic Press: London, UK, 1991; pp. 1–66. [Google Scholar]
- Weber, M. The formation of pollenkitt in Apium nodiflorum (Apiaceae). Ann. Bot. 1992, 70, 573–577. [Google Scholar] [CrossRef]
- Spurr, A.R. A low-viscosity epoxy resin embedding medium for electron microscopy. J. Ultrastruct. Res. 1969, 26, 31–43. [Google Scholar] [CrossRef] [PubMed]
- Bozzola, J.J.; Russell, L.D. Autoradiography and Radioautography. In Electron Microscopy: Principles and Techniques for Biologists; Jones & Bartlett Learning: Sudbury, MA, USA, 1999; pp. 293–308. [Google Scholar]
- Hanaichi, T.; Sato, T.; Iwamoto, T.; Malavasi-Yamashiro, J.; Hoshino, M.; Mizuno, N. A stable lead by modification of Sato’s method. Microscopy 1986, 35, 304–306. [Google Scholar]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Akwu, N.A.; Naidoo, Y.; Singh, M.; Dewir, Y.H.; Magyar-Tábori, K.; Lekhooa, M.; Aremu, A.O. Development and Biomechanics of Grewia lasiocarpa E. Mey. Ex Harv. Trichomes Exudate. Plants 2023, 12, 2198. https://doi.org/10.3390/plants12112198
Akwu NA, Naidoo Y, Singh M, Dewir YH, Magyar-Tábori K, Lekhooa M, Aremu AO. Development and Biomechanics of Grewia lasiocarpa E. Mey. Ex Harv. Trichomes Exudate. Plants. 2023; 12(11):2198. https://doi.org/10.3390/plants12112198
Chicago/Turabian StyleAkwu, Nneka Augustina, Yougasphree Naidoo, Moganavelli Singh, Yaser Hassan Dewir, Katalin Magyar-Tábori, Makhotso Lekhooa, and Adeyemi Oladapo Aremu. 2023. "Development and Biomechanics of Grewia lasiocarpa E. Mey. Ex Harv. Trichomes Exudate" Plants 12, no. 11: 2198. https://doi.org/10.3390/plants12112198