
Origin of the Rare Hybrid Genus ×Trisetokoeleria Tzvelev (Poaceae) According to Molecular Phylogenetic Data
 , , , , , and
, , , , , and
Abstract
:1. Introduction
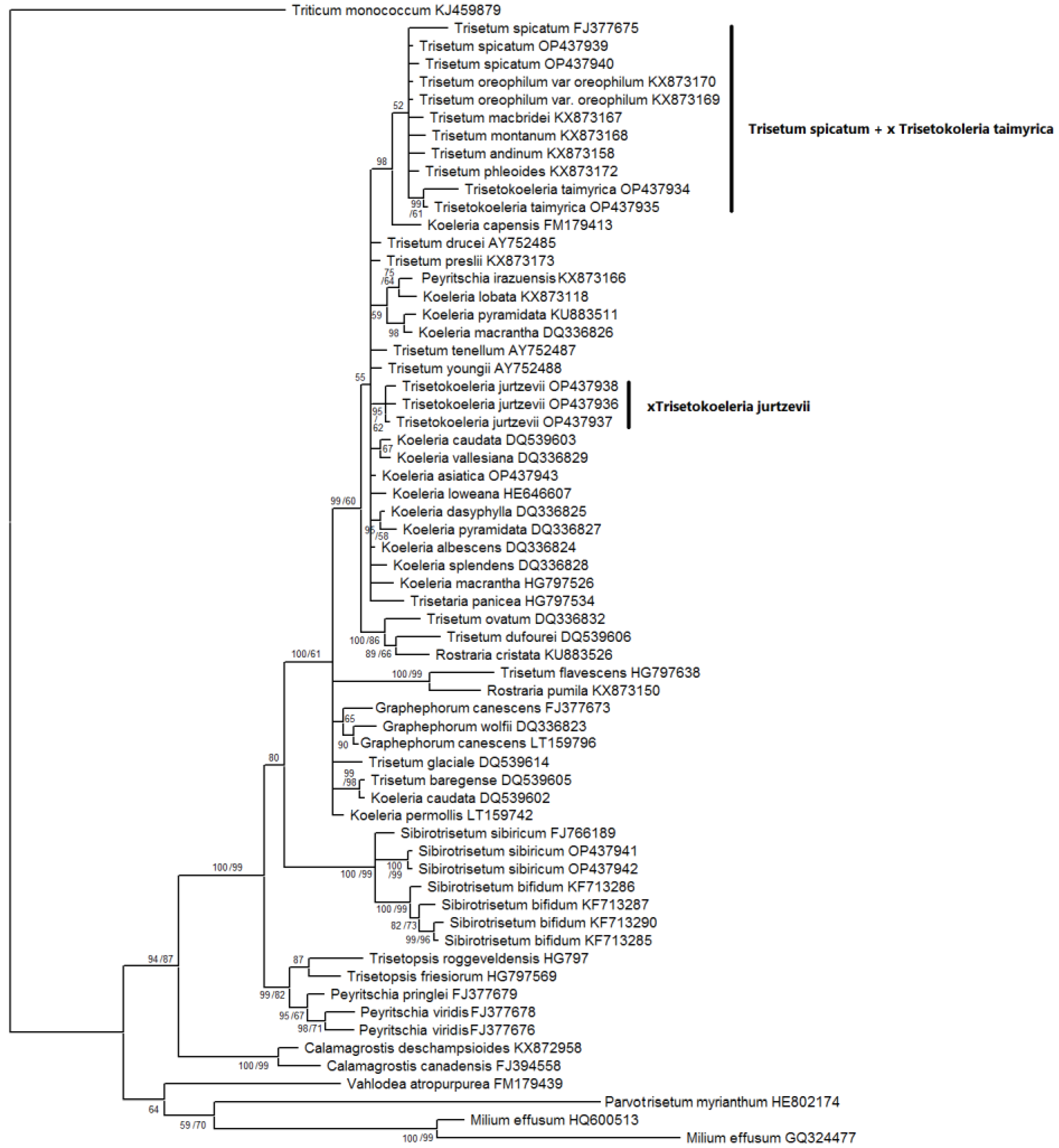
2. Results
Pollen Fertility Analysis
3. Discussion
3.1. Overview of the Genus
3.2. Origin of Each Species According to the Molecular Data
3.3. Allopolyploid Parental Taxa of ×Trisetokoeleria and Problem of Delimitation of the Genera
4. Conclusions
5. Materials and Methods
5.1. Molecular Phylogenetic Analysis
5.2. Pollen Fertility Detection
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Soreng, R.J.; Peterson, P.M.; Romaschenko, K.; Davidse, G.; Judziewicz, E.J.; Zuloaga, F.O.; Filgueiras, T.S.; Morrone, O. A worldwide phylogenetic classification of the Poaceae (Gramineae). J. Syst. Evol. 2015, 53, 117–137. [Google Scholar] [CrossRef]
- Saarela, J.M.; Bull, R.D.; Paradis, M.J.; Ebata, S.N.; Peterson, P.M.; Soreng, R.J.; Paszko, B. Molecular phylogenetics of cool-season grasses in the subtribes Agrostidinae, Anthoxanthinae, Aveninae, Brizinae, Calothecinae, Koeleriinae and Phalaridinae (Poaceae, Pooideae, Poeae, Poeae chloroplast group 1). PhytoKeys 2017, 87, 1–139. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tzvelev, N.N. Notes on some genera of grasses of the USSR flora. Nov. Sist. Vyssh. Rast. 1971, 7, 42–59. (In Russian) [Google Scholar]
- Probatova, N.S.; Yurtsev, B.A. New taxa of the family Poaceae from the north-east of the USSR. Bot. Zhurn. 1984, 69, 688–692. (In Russian) [Google Scholar]
- Tzvelev, N.N. New taxa of the grasses (Poaceae). Nov. Sist. Vyssh. Rast. 1974, 11, 70–72. (In Russian) [Google Scholar]
- Quintanar, A.; Castroviejo, S.; Catalán, P. Phylogeny of the tribe Aveneae (Pooideae, Poaceae) inferred from plastid trnT-F and nuclear ITS sequences. Am. J. Bot. 2007, 94, 1554–1569. [Google Scholar] [CrossRef]
- Wölk, A.; Röser, M. Hybridization and long-distance colonization in oat-like grasses of South and East Asia, including an amended circumscription of Helictotrichon and the description of the new genus Tzveleviochloa (Poaceae). Taxon 2017, 66, 20–43. [Google Scholar] [CrossRef]
- Barberá, P.; Quintanar, A.; Aedo, C. New combinations, new names, typifications, and a new section, sect. Hispanica, in Koeleria (Poeae, Poaceae). Phytoneuron 2019, 46, 1–13. [Google Scholar]
- Barberá, P.; Soreng, R.J.; Peterson, P.M.; Romaschenko, K.; Quintanar, A.; Aedo, C. Molecular phylogenetic analysis resolves Trisetum (Poaceae: Pooideae: Koeleriinae) polyphyletic: Evidence for a new genus, Sibirotrisetum and resurrection of Acrospelion. J. Syst. Evol. 2020, 58, 517–526. [Google Scholar] [CrossRef] [Green Version]
- Matyášek, R.; Renny-Byfield, S.; Fulneček, J.; Macas, J.; Grandbastien, M.A.; Nichols, R.; Leitch, A.; Kovařík, A. Next generation sequencing analysis reveals a relationship between rDNA unit diversity and locus number in Nicotiana diploids. BMC genomics 2012, 13, 722. [Google Scholar] [CrossRef] [Green Version]
- Grigoryan, M.Y.; Bobrov, A.A.; Brunton, D.F.; Volkova, P.A.; Logacheva, M.D.; Neretina, T.V. Next generation DNA sequencing reveals allopolyploid origin of decaploid Isoëtes lacustris (Isoëtaceae). Aquatic Botany 2021, 170, 103326. [Google Scholar] [CrossRef]
- Pellicer, J.; Balant, M.; Fernández, P.; Rodríguez González, R.; Hidalgo, O. Morphological and genome-wide evidence of homoploid hybridisation in Urospermum (Asteraceae). Plants 2022, 11, 182. [Google Scholar] [CrossRef] [PubMed]
- Tzvelev, N.N.; Probatova, N.S. Grasses of Russia; KMK Scientific Press: Moscow, Russia, 2019; p. 646. (In Russian) [Google Scholar]
- Álvarez, I.J.F.W.; Wendel, J.F. Ribosomal ITS sequences and plant phylogenetic inference. Mol. Phyl. Evol. 2003, 29, 417–434. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gurushidze, M.; Mashayekhi, S.; Blattner, F.R.; Friesen, N.; Fritsch, R.M. Phylogenetic relationships of wild and cultivated species of Allium section Cepa inferred by nuclear rDNA ITS sequence analysis. Plant Syst. Evol. 2007, 269, 259–269. [Google Scholar] [CrossRef]
- Soltis, P.S.; Soltis, D.E. The role of hybridization in plant speciation. Annu Rev. Plant Biol. 2009, 60, 561–588. [Google Scholar] [CrossRef] [Green Version]
- Kamelin, R.V. Flora Altaia; Azbuka: Barnaul, Russia, 2005; p. 338. (In Russian) [Google Scholar]
- Kamelin, R.V. The Peculiarities of Flowering Plants Speciation; Prilozhenie No. 1; Trudy Zoologicheskogo Instituta RAN: Saint Petersburg, Russia, 2009; pp. 141–149. (In Russian) [Google Scholar]
- Ebersbach, J.; Tkach, N.; Röser, M.; Favre, A. The role of hybridisation in the making of the species-rich arctic-alpine genus Saxifraga (Saxifragaceae). Diversity 2020, 12, 440. [Google Scholar] [CrossRef]
- Tkach, N.; Schneider, J.; Döring, E.; Wölk, A.; Hochbach, A.; Nissen, J.; Winterfeld, G.; Meyer, S.; Gabriel, J.; Hoffmann, M.H.; et al. Phylogenetic lineages and the role of hybridization as driving force of evolution in grass supertribe Poodae. Taxon 2020, 69, 234–277. [Google Scholar] [CrossRef] [Green Version]
- Consaul, L.L.; Gillespie, L.J.; Waterway, M.J. Evolution and polyploid origins in North American Arctic Puccinellia (Poaceae) based on nuclear ribosomal spacer and chloroplast DNA sequences. Am. J. Bot. 2010, 97, 324–336. [Google Scholar] [CrossRef] [Green Version]
- Rice, A.; Šmarda, P.; Novosolov, M.; Drori, M.; Glick, L.; Sabath, N.; Meiri, S.; Belmaker, J.; Mayrose, I. The global biogeography of polyploid plants. Nat. Ecol. Evol. 2019, 3, 265–273. [Google Scholar] [CrossRef]
- Favarger, C. Sur l’emploi des nombres chromo-somiques en géographie botanique historique. Ber. Geobot. Inst. Rübel 1961, 32, 119–146. [Google Scholar]
- Mandáková, T.; Joly, S.; Krzywinski, M.; Mummenhoff, K.; Lysak, M.A. Fast Diploidization in Close Mesopolyploid Relatives of Arabidopsis. Plant Cell 2010, 22, 2277–2290. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodionov, A.V.; Shneyer, V.S.; Gnutikov, A.A.; Nosov, N.N.; Punina, E.O.; Zhurbenko, P.M.; Loskutov, I.G.; Muravenko, O.V. Species dialectics: From initial uniformity, through the greatest possible diversity to ultimate uniformity. Bot. Zhurn. 2020, 105, 835–853. (In Russian) [Google Scholar] [CrossRef]
- Volkov, R.A.; Panchuk, I.I.; Borisjuk, N.V.; Hosiawa-Baranska, M.; Maluszynska, J.; Hemleben, V. Evolutional dynamics of 45S and 5S ribosomal DNA in ancient allohexaploid Atropa belladonna. BMC Plant Biol. 2017, 17, 21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Petrovsky, V.V.; Zhukova, P.G. Chromosome numbers and taxonomy of some plant species of the Wrangel Island. Bot. Zhurn. 1981, 66, 380–387. (In Russian) [Google Scholar]
- Zhukova, P.G. Chromosome numbers of some plant species of north-eastern Asia. Bot. Zhurn. 1982, 67, 360–365. (In Russian) [Google Scholar]
- Murray, B.G.; de Lange, P.J.; Ferguson, A.R. Nuclear DNA variation, chromosome numbers and polyploidy in the endemic and indigenous grass flora of New Zealand. Ann. Bot. (Oxford) 2005, 96, 1293–1305. [Google Scholar] [CrossRef] [Green Version]
- Seledets, V.P.; Probatova, N.S. Ecological area of species: Karyological aspects. In Karyology, Karyosystematics and Molecular Phylogeny. V International Symposium; Komarov Botanical Institute of the RAS: St.-Petersburg, Russia, 2005; pp. 95–97. (In Russian) [Google Scholar]
- Goncharov, N.F. Koeleria Pers. In Flora of the USSR; Rozhevitz, R.Y., Shishkin, B.K., Eds.; Izdatel’stvo Akademii Nauk SSSR: Leningrad, Russia, 1934; pp. 323–337. (In Russian) [Google Scholar]
- Soreng, R.J. Chloroplast-DNA phylogenetics and biogeography in a reticulating group: Study in Poa (Poaceae). Am. J. Bot. 1990, 77, 1383–1400. [Google Scholar] [CrossRef]
- Winkworth, R.C.; Wagstaff, S.J.; Glenny, D.; Lockhart, P.J. Evolution of the New Zealand mountain flora: Origins, diversification and dispersal. Org. Divers. Evol. 2005, 5, 237–247. [Google Scholar] [CrossRef] [Green Version]
- Rodionov, A.V.; Nosov, N.N.; Kim, E.S.; Machs, E.M.; Punina, E.O.; Probatova, N.S. Origin of high polyploid genomes of bluegrasses (Poa L.) and phenomenon of the gene flow between Northern Pacific and Sub-Antarctic islands. Rus. J. Gen. 2010, 46, 1407–1416. [Google Scholar] [CrossRef]
- Nie, Z.-L.; Sun, H.; Manchester, S.R.; Meng, Y.; Luke, Q.; Wen, J. Evolution of the intercontinental disjunctions in six continents in the Ampelopsis clade of the grape family (Vitaceae). BMC Evol. Biol. 2012, 12, 17. [Google Scholar] [CrossRef] [Green Version]
- Nosov, N.N.; Tikhomirov, V.N.; Machs, E.M.; Rodionov, A.V. On polyphyly of the former section Ochlopoa and the hybridogenic section Acroleucae (Poa, Poaceae): Insights from molecular phylogenetic analyses. Nord. J. Bot. 2019, 37, e02015. [Google Scholar] [CrossRef]
- Tzvelev, N.N. On the genus Koeleria Pers. (Poaceae) in Russia. Novosti Sist. Vyssh. Rast. 2011, 42, 63–89. (In Russian) [Google Scholar] [CrossRef]
- Mason-Gamer, R.J. Phylogeny of a genomically diverse group of Elymus (Poaceae) allopolyploids reveals multiple levels of reticulation. PLoS ONE 2013, 8, e78449. [Google Scholar] [CrossRef] [Green Version]
- Tan, L.; Huang, Q.-X.; Song, Y.; Wu, D.-D.; Cheng, Y.-R.; Zhang, C.-B.; Sha, L.-N.; Fan, X.; Kang, H.-Y.; Wang, Y.; et al. Biosystematics studies on Elymus breviaristatus and Elymus sinosubmuticus (Poaceae: Triticeae). BMC Plant Biol. 2022, 22, 57. [Google Scholar] [CrossRef] [PubMed]
- Müller, K. SeqState—primer design and sequence statistics for phylogenetic DNA data sets. App. Bioinf. 2005, 4, 65–69. [Google Scholar] [CrossRef]
- Ronquist, F.; Teslenko, M.; van der Mark, P.; Ayres, D.L.; Darling, A.; Höhna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MrBayes 3.2: Efficient Bayesian phylogenetic inference and model choice across a large model space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef] [Green Version]
- Ridgway, K.P.; Duck, J.M.; Young, J.P.W. Identification of roots from grass swards using PCR-RFLP and FFLP of the plastid trnL (UAA) intron. BMC Ecol. 2003, 3, 8. [Google Scholar] [CrossRef]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Innis, M.A., Gelfand, D.H., Sninsky, J.J., White, T.J., Eds.; Academic Press: New York, NY, USA, 1990; pp. 315–322. [Google Scholar] [CrossRef]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [Green Version]
- Okonechnikov, K.; Golosova, O.; Fursov, M.; the UGENE team. Unipro UGENE: A unified bioinformatics toolkit. Bioinformatics 2012, 28, 1166–1167. [Google Scholar] [CrossRef] [Green Version]
- Aronesty, E. Comparison of sequencing utility program. Open Bioinform. J. 2013, 7, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Rognes, T.; Flouri, T.; Nichols, B.; Quince, C.; Mahe, F. VSEARCH: A versatile open source tool for metagenomics. PeerJ. 2016, 4, e2584. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Clement, M.; Posada, D.; Crandall, K.A. TCS: A computer program to estimate gene genealogies. Mol. Ecol. 2000, 9, 1657–1660. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Múrias dos Santos, A.; Cabezas, M.P.; Tavares, A.I.; Xavier, R.; Branco, M. tcsBU: A tool to extend TCS network layout and visualization. Bioinformatics 2016, 32, 627–628. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huson, D.H.; Bryant, D. Application of Phylogenetic Networks in Evolutionary Studies. Mol. Biol. Evol. 2006, 23, 254–267. [Google Scholar] [CrossRef]
- Taberlet, P.; Gielly, L.; Pautou, G.; Bouve, J. Universal primers for amplification of three 430 non-coding regions 431 of chloroplast DNA. Plant Mol. Biol. 1991, 17, 1105–1109. [Google Scholar] [CrossRef]
- Kress, W.J.; Wurdack, K.J.; Zimmer, E.A.; Weigt, L.A.; Janzen, D.H. Use of DNA barcodes to identify flowering plants. Proc. Natl. Acad. Sci. USA 2005, 102, 8369–8374. [Google Scholar] [CrossRef] [Green Version]
- Johnson, L.A.; Soltis, D.E. Phylogenetic inference in Saxifragaceae sensu stricto and Gilia (Polemoniaceae) using matK sequences. Ann. Mo. Bot. Gard. 1995, 82, 149–175. [Google Scholar] [CrossRef]
- Edgar, R.C. MUSCLE: Multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Res. 2004, 32, 1792–1797. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Sample ID | 2n | Country of Origin | Collected by | Accession Number in Genbank Database | Number of Accessions | Total Number of Reads | Ribotype Number in Figure 1 | Ribotype Symbol | Number of Reads | % from the Total Number of the Reads |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Trisetum spicatum | L1 | 28 | Russian Federation: Russia: Yakutia, | S.V. Chinenko | OP557123–OP557200 | 78 | 21,799 | 1 | T1 | 16,839 | 77 |
| Anabarsky District | 3 | T2 | 50 | 0.2 | |||||||
| 7 | T5 | 18 | 0.08 | ||||||||
| Trisetum subalpestre | L13 | 28 | Russian Federation: Chukotka | A.A. Korobkov, B.A. Yurtzev | OP557201–OP557245 | 45 | 26,464 | 2 | Ts | 12,538 | 47 |
| 3 | T2 | 8685 | 33 | ||||||||
| Trisetum wrangelense | M3 | 28 | Russian Federation: Chukotka, | B. Yurtzev, T. Polozova | OP557389–OP557522 | 134 | 27,647 | 1 | T1 | 21,565 | 78 |
| Wrangel Island | 3 | T2 | 50 | 0.1 | |||||||
| 7 | T5 | 12 | 0.04 | ||||||||
| Trisetum ruprechtianum | M2 | 14 | Russian Federation: Chukotka | V.V. Petrovsky, T.V. Plieva | OP557327–OP557388 | 62 | 12,740 | 4 | T3 | 2548 | 20 |
| 5 | T4 | 1126 | 9 | ||||||||
| Koeleria asiatica | G18 | 14, 28 | Russian Federation: Yakutia, | T.M. Koroleva | OP557523–OP557563 | 41 | 22,341 | 3 | T2 | 12,064 | 54 |
| Anabarsky District | 6 | Ka | 1505 | 7 | |||||||
| 1 | T1 | 1085 | 5 | ||||||||
| ×Trisetokoeleria gorodkowii | G19 | N/A | Russian Federation: Yakutia | A.A. Korobkov, T.M. Koroleva | OP557281–OP557290 | 10 | 12,048 | 7 | T5 | 3492 | 29 |
| ×Trisetokoeleria jurtzevii | G16 | N/A | Russian Federation: Yakutia | P.A. Gogoleva, T.M. Koroleva | OP557246–OP557280 | 35 | 14,620 | 8 | Tj | 7090 | 48 |
| 3 | T2 | 2824 | 19 | ||||||||
| ×Trisetokoeleria taimyrica | G15 | N/A | Russian Federation: Yakutia | P.A. Gogoleva, T.M. Koroleva | OP557291–OP557326 | 36 | 19,245 | 1 | T1 | 14,574 | 76 |
| Species | Country of Origin | Collected by | Genbank Number, trnL-trnF | Genbank Number, trnK-rps16 | Genbank Number, ITS |
|---|---|---|---|---|---|
| ×Trisetokoeleria jurtzevii | Russia: Yakutia | P.A. Gogoleva, T.M. Koroleva | OP429186 | OP503155 | OP437936 |
| ×Trisetokoeleria jurtzevii | Russia: Yakutia | P.A. Gogoleva, T.M. Koroleva | OP429187 | OP503156 | OP437937 |
| ×Trisetokoeleria jurtzevii | Russia: Yakutia | T.M. Koroleva | OP429188 | OP503157 | OP437938 |
| ×Trisetokoeleria taimyrica | Russia: Yakutia | P.A. Gogoleva, T.M. Koroleva | OP429184 | OP503153 | OP437934 |
| ×Trisetokoeleria taimyrica | Russia: Yakutia | P.A. Gogoleva, T.M. Koroleva | OP429185 | OP503154 | OP437935 |
| Sibirotrisetum sibiricum | Russia: Yakutia | M. Telyatnikov | OP503160 | OP437941 | |
| Sibirotrisetum sibiricum | Russia: Yakutia | S.V. Chinenko | OP429191 | OP503161 | OP437942 |
| Trisetum spicatum | Russia: Yakutia | S.V. Chinenko | OP429189 | OP503158 | OP437939 |
| Trisetum spicatum | Russia: Yakutia, Anabarsky District | S.V. Chinenko | OP429190 | OP503159 | OP437940 |
| Koeleria asiatica | Russia: Yakutia, Anabarsky District | T.M. Koroleva | OP429192 | OP503162 | OP437943 |
| 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 3 | ||
| 9 | 0 | 1 | 1 | 4 | 4 | 4 | 5 | 5 | 6 | 7 | 7 | 2 | 2 | 4 | 4 | 5 | 5 | 6 | 8 | 3 | |
| 7 | 9 | 4 | 6 | 0 | 7 | 9 | 0 | 1 | 8 | 4 | 9 | 7 | 9 | 4 | 8 | 1 | 5 | 0 | 1 | 1 | |
| T1 | G | C | C | C | G | A | A | A | C | G | T | G | A | G | T | T | C | A | G | C | C |
| Ts | A | . | . | . | . | . | . | . | . | . | . | . | C | . | . | . | . | . | . | . | . |
| T2 | . | . | . | . | . | . | . | . | . | . | . | . | C | . | . | . | . | . | . | . | . |
| T3 | . | . | T | . | C | T | . | . | . | . | . | . | T | . | C | . | T | C | . | . | . |
| T4 | . | . | T | . | C | T | . | . | . | . | A | . | T | A | C | . | T | C | . | . | . |
| Ka | . | . | . | . | . | . | D | D | T | . | . | . | C | . | . | . | . | . | . | T | . |
| T5 | . | . | . | . | . | . | . | . | . | A | . | . | . | . | . | . | . | . | . | . | . |
| Tj | . | . | . | T | . | . | . | . | . | . | . | A | C | . | . | . | . | . | . | . | . |
| 55-G15 | . | . | . | . | . | . | . | . | . | . | . | . | T | . | . | C | . | . | C | . | . |
| 14-G15(3) | . | T | T | . | . | . | . | . | . | . | . | . | T | . | . | C | . | . | C | . | . |
| Peyritschia deyeuxioides FJ377668 | . | . | . | . | . | . | . | . | . | . | . | . | T | . | . | C | . | . | C | . | . |
| Cinnagrostis viridiflavescens KX873106 | . | . | . | . | . | . | . | . | . | . | . | . | T | . | . | C | . | . | C | . | T |
| Species | Total | Stained | Abortive | Abortive Pollen Percent |
|---|---|---|---|---|
| Trisetum spicatum | 1042 | 896 | 146 | 14.0% |
| Trisetum ruprechtianum | 1007 | 886 | 121 | 12.0% |
| Koeleria asiatica | 1005 | 932 | 74 | 7.4% |
| ×T. jurtzevii | 1000 | 0 | 1000 | 100% |
| ×T. gorodkowii | 1140 | 627 | 513 | 45.0% |
| ×T. taimyrica | 1080 | 885 | 195 | 18.1% |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gnutikov, A.A.; Nosov, N.N.; Koroleva, T.M.; Punina, E.O.; Probatova, N.S.; Shneyer, V.S.; Rodionov, A.V. Origin of the Rare Hybrid Genus ×Trisetokoeleria Tzvelev (Poaceae) According to Molecular Phylogenetic Data. Plants 2022, 11, 3533. https://doi.org/10.3390/plants11243533
Gnutikov AA, Nosov NN, Koroleva TM, Punina EO, Probatova NS, Shneyer VS, Rodionov AV. Origin of the Rare Hybrid Genus ×Trisetokoeleria Tzvelev (Poaceae) According to Molecular Phylogenetic Data. Plants. 2022; 11(24):3533. https://doi.org/10.3390/plants11243533
Chicago/Turabian StyleGnutikov, Alexander A., Nikolai N. Nosov, Tatiana M. Koroleva, Elizaveta O. Punina, Nina S. Probatova, Victoria S. Shneyer, and Alexander V. Rodionov. 2022. "Origin of the Rare Hybrid Genus ×Trisetokoeleria Tzvelev (Poaceae) According to Molecular Phylogenetic Data" Plants 11, no. 24: 3533. https://doi.org/10.3390/plants11243533