Phylogenetic Placement and Phylogeography of Large-Flowered Lotus Species (Leguminosae) Formerly Classified in Dorycnium: Evidence of Pre-Pleistocene Differentiation of Western and Eastern Intraspecific Groups

 ,
,  and
and
Abstract
:1. Introduction
2. Results
2.1. Analysis of nrDNA ITS
2.2. Analysis of Plastid DNA Dataset
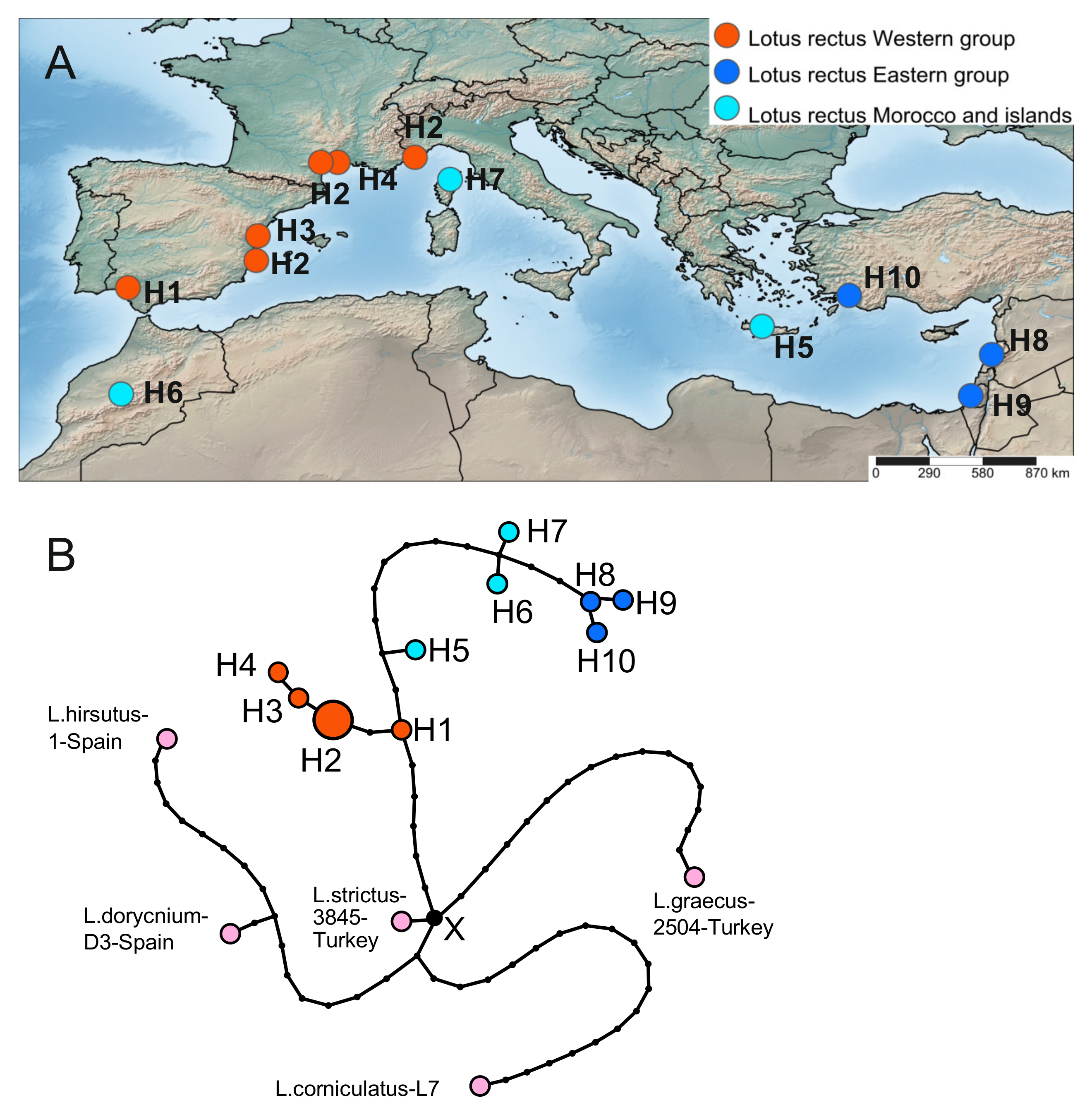
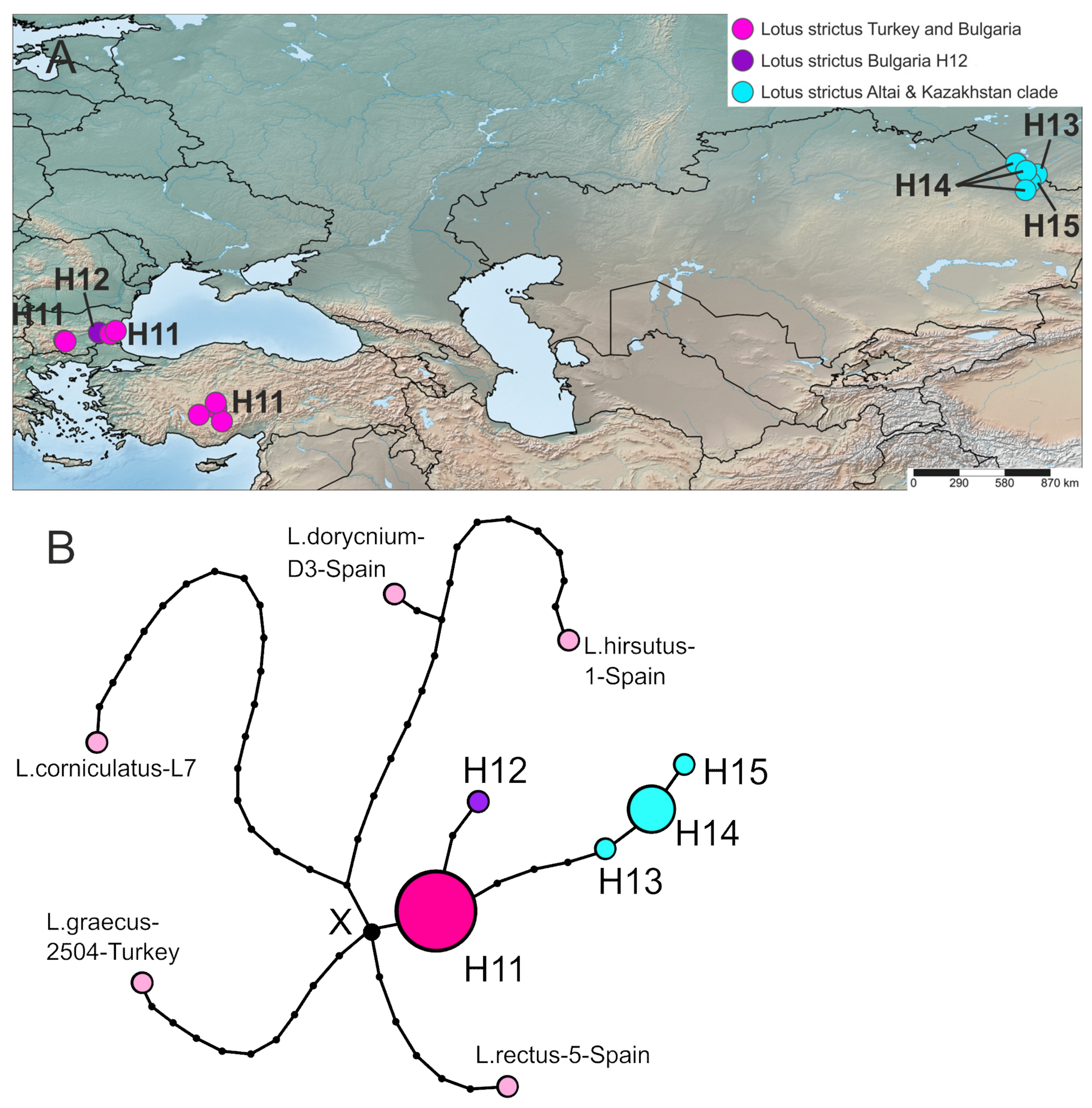
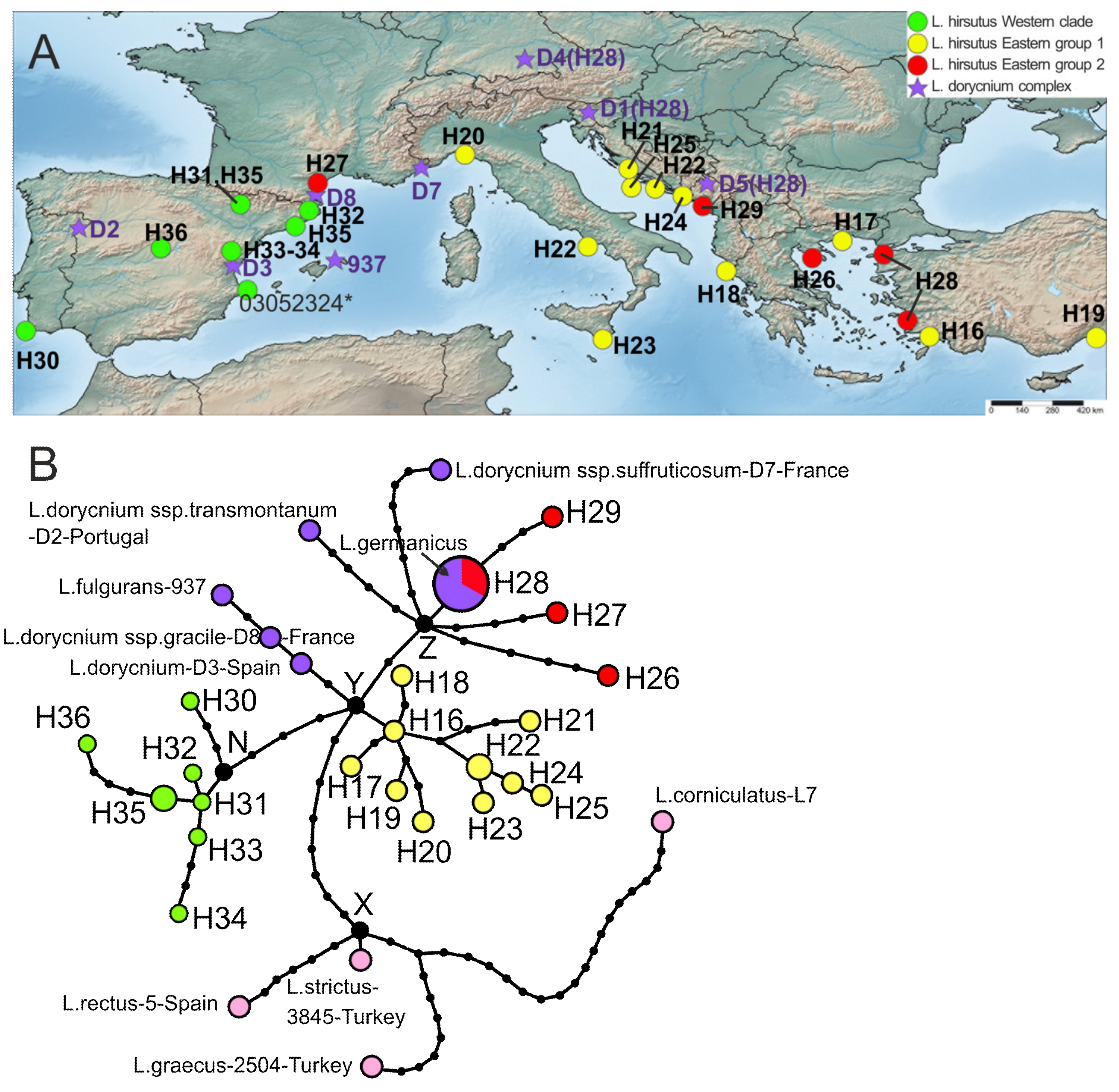
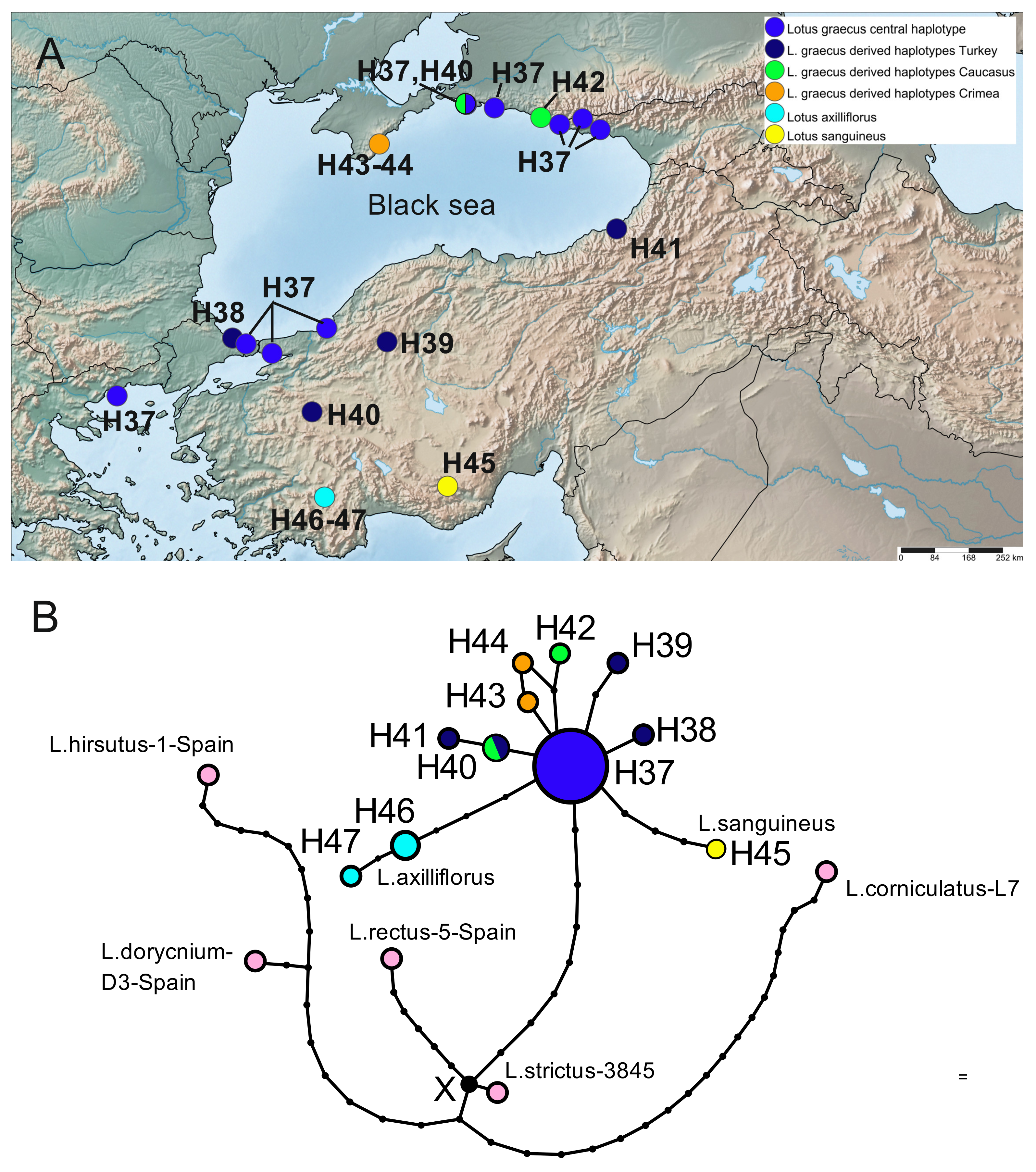
2.3. Phylogeography Based on the Plastid Dataset
2.3.1. Lotus rectus
2.3.2. Lotus strictus
2.3.3. Lotus hirsutus
2.3.4. Lotus graecus and Related Taxa
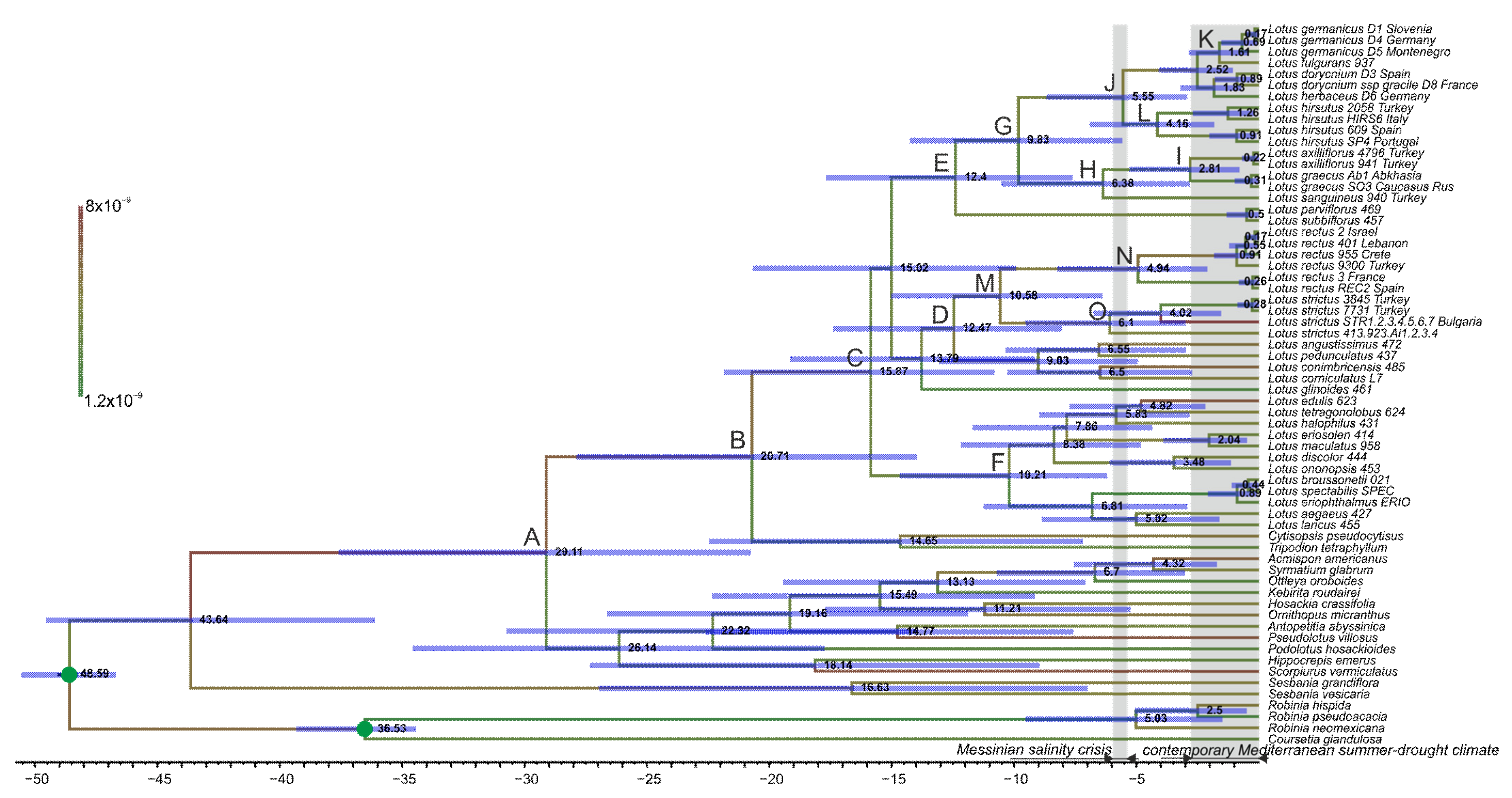
2.4. Dating Phylogeny of Lotus
3. Discussion
3.1. Diversification of the Genus Lotus Much Pre-Dates Formation of the Extant Mediterranean Biome
3.2. Evolutionary Histories of Individual Species Reveal Common Biogeographic Patterns
4. Materials and Methods
4.1. Plant Material
4.2. DNA Extraction, Amplification and Sequencing
4.3. Phylogenetic Analyses
4.4. Dating Analyses
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A
Appendix B
References
- Sokoloff, D.D.; Lock, J.M. Tribe Loteae. In Legumes of the World; Lewis, G., Schrire, B., Mackinder, B., Lock, M., Eds.; The Royal Botanic Gardens: Kew, UK, 2005; pp. 455–465. [Google Scholar]
- Kramina, T.E.; Sokoloff, D.D. On Lotus sect. Erythrolotus and related taxa (Leguminosae). Byull. Moskovsk. Obshch. Isp. Prir. Otd. Biol. 2003, 108, 59–62. (In Russian) [Google Scholar]
- Sokoloff, D.D. On taxonomy and phylogeny of the tribe Loteae DC. (Leguminosae). Byull. Moskovsk. Obshch. Isp. Prir. Otd. Biol. 2003, 108, 35–48. (In Russian) [Google Scholar]
- Degtjareva, G.V.; Kramina, T.E.; Sokoloff, D.D.; Samigullin, T.H.; Valiejo-Roman, C.M.; Antonov, A.S. Phylogeny of the genus Lotus (Leguminosae, Loteae): Evidence from nrITS sequences and morphology. Can. J. Bot. 2006, 84, 813–830. [Google Scholar] [CrossRef]
- Degtjareva, G.V.; Kramina, T.E.; Sokoloff, D.D.; Samigullin, T.H.; Sandral, G.; Valiejo-Roman, C.M. New data on nrITS phylogeny of Lotus (Leguminosae, Loteae). Wulfenia 2008, 15, 35–49. [Google Scholar]
- Kramina, T.E.; Degtjareva, G.V.; Samigullin, T.H.; Valiejo-Roman, C.M.; Kirkbride, J.H., Jr.; Volis, S.; Deng, T.; Sokoloff, D.D. Phylogeny of Lotus (Leguminosae: Loteae): Partial incongruence between nrITS, nrETS and plastid markers and biogeographic implications. Taxon 2016, 65, 997–1018. [Google Scholar] [CrossRef]
- Jaén-Molina, R.; Marrero-Rodríguez, Á.; Caujapé-Castells, J.; Ojeda, D.I. Molecular phylogenetics of Lotus (Leguminosae) with emphasis in the tempo and patterns of colonization in the Macaronesian region. Mol. Phylogenet. Evol. 2021, 154, 106970. [Google Scholar] [CrossRef] [PubMed]
- Allan, G.J.; Zimmer, E.A.; Wagner, W.L.; Sokoloff, D.D. Molecular phylogenetic analyses of tribe Loteae (Leguminosae): Implications for classification and biogeography. In Advances in Legume Systematics; Klitgaard, B.B., Bruneau, A., Eds.; Royal Botanic Gardens: Kew, UK, 2003; pp. 371–393, Part 10. [Google Scholar]
- Polhill, R.M. Loteae, Coronilleae. In Advances in Legume Systematics; Polhill, R.M., Raven, P.H., Eds.; Royal Botanic Gardens: Kew, UK, 1981; pp. 371–375. [Google Scholar]
- Ball, P.W. Dorycnium Miller. In Flora Europaea; Tutin, T.G., Heywood, V.H., Burges, N.A., Moore, D.M., Valentine, D.H., Walters, S.M., Webb, D.A., Eds.; Cambridge University Press: Cambridge, UK, 1968; Volume 2, pp. 172–173. [Google Scholar]
- Ball, P.W.; Chrtková-Žertová, A. Lotus L. In Flora Europaea; Tutin, T.G., Heywood, V.H., Burges, N.A., Moore, D.M., Valentine, D.H., Walters, S.M., Webb, D.A., Eds.; Cambridge University Press: Cambridge, UK, 1968; Volume 2, pp. 173–176. [Google Scholar]
- Greene, E.L. Enumeration of the North American Loti. Pittonia 1890, 2, 133–150. [Google Scholar]
- Reichenbach, L. Flora Germanica Excursoria; Carolum Cnobloch: Lipsiae, Saxonia, 1832; Volume 2, pp. 505–507, 515–517. [Google Scholar]
- Rikli, M. Die Gattung Dorycnium. Bot. Jahrb. Syst. 1901, 31, 314–404. [Google Scholar]
- Lassen, P. Acmispon sect. Simpeteria, Acmispon roudairei, Dorycnium strictum, Lotus benoistii. Willdenovia 1986, 16, 107–112. [Google Scholar]
- Gillett, J.B. Lotus in Africa south of the Sahara (excluding Cape Verde Islands and Socotra) and its distinction from Dorycnium. Kew Bull. 1958, 13, 361–381. [Google Scholar] [CrossRef]
- Sokoloff, D.D. Morphology and Classification of the Tribe Loteae DC. of the Family Leguminosae. Ph.D. Thesis, Moscow State University, Moscow, Russia, 2003. (In Russian). [Google Scholar]
- Legume Phylogeny Working Group (LPWG). A new subfamily classification of the Leguminosae based on a taxonomically comprehensive phylogeny. Taxon 2017, 66, 44–77. [Google Scholar] [CrossRef] [Green Version]
- Cardoso, D.; de Queiroz, L.P.; Pennigton, R.T.; de Lima, H.C.; Fonty, E.; Wojciechowski, M.F.; Lavin, M. Revisiting the phylogeny of papilionoid legumes: New insight from comprehensively sampled early-branching lineages. Am. J. Bot. 2012, 99, 1991–2013. [Google Scholar] [CrossRef] [PubMed]
- Lavin, M.; Herendeen, P.S.; Wojciechowski, M.F. Evolutionary rates analysis of Leguminosae implicates a rapid diversification of lineages during the Tertiary. Syst. Biol. 2005, 54, 530–549. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cardoso, D.; Pennington, R.T.; de Queiroz, L.P.; Boatwright, J.S.; Van Wyk, B.-E.; Wojciechowski, M.F.; Lavin, M. Reconstructing the deep-branching relationships of the papilionoid legumes. S. Arf. J. Bot. 2013, 89, 58–75. [Google Scholar] [CrossRef] [Green Version]
- Farruggia, F.T.; Lavin, M.; Wojciechowski, M.F. Phylogenetic systematics and biogeography of the pantropical genus Sesbania (Leguminosae). Syst. Bot. 2018, 43, 414–429. [Google Scholar] [CrossRef]
- Herendeen, P.S.; Crepet, W.L.; Dilcher, D.L. The fossil history of the Leguminosae: Phylogenetic and biogeographic implications. In Advances in Legume Systematics, Part 4, the Fossil Record; Herendeen, P.S., Dilcher, D.L., Eds.; Royal Botanic Gardens: Kew, UK, 1992; pp. 303–316. [Google Scholar]
- Matten, L.C.; Gastaldo, R.A.; Lee, M.R. Fossil Robinia wood from the western United States. Rev. Palaeobot. Palynol. 1977, 24, 195–208. [Google Scholar] [CrossRef]
- Berggren, W.A.; Kent, D.V.; Swisher, C.C., III; Aubry, M.-P. A revised Cenozoic geochronology and chronostratigraphy. In Geochronology, Time Scales and Global Stratigraphic Correlation; Berggren, W.A., Kent, D.V., Aubry, M.-P., Hardenbol, J., Eds.; SEPM Special Publication No 54; SEPM: Tulsa, OK, USA, 1995; pp. 129–212. [Google Scholar]
- Lavin, M.; Wojciechowski, M.F.; Gasson, P.; Hughes, C.; Wheeler, E. Phylogeny of robinioid legumes (Fabaceae) revisited: Coursetia and Gliricidia recircumscribed, and a biogeographical appraisal of the Caribbean endemics. Syst. Bot. 2003, 28, 387–409. [Google Scholar] [CrossRef]
- Médail, F.; Diadema, K. Glacial refugia influence plant diversity patterns in the Mediterranean Basin. J. Biogeogr. 2009, 36, 1333–1345. [Google Scholar] [CrossRef]
- Hewitt, G.M. Mediterranean peninsulas: The evolution of hotspots. In Biodiversity Hotspots; Zachos, F.E., Habel, J.C., Eds.; Springer: Berlin/Heidelberg, Germany, 2011; pp. 123–147. [Google Scholar] [CrossRef]
- Feliner, G.N. Patterns and processes in plant phylogeography in the Mediterranean Basin. A review. Perspect. Plant Ecol. 2014, 16, 265–278. [Google Scholar] [CrossRef]
- Myers, N.; Mittermeier, R.A.; Mittermeier, C.G.; da Fonseca, G.A.B.; Kent, J. Biodiversity hotspots for conservation priorities. Nature 2000, 403, 853–858. [Google Scholar] [CrossRef]
- Perrino, E.V.; Tomaselli, V.; Costa, R.; Pavone, P. Conservation status of habitats (Directive 92/43 EEC) of coastal and low hill belts in a mediterranean biodiversity hot spot (Gargano—Italy). Plant Biosyst. 2013, 147, 1006–1028. [Google Scholar] [CrossRef]
- Xofis, P.; Tsiourlis, G.; Konstantinidis, P. A Fire Danger Index for the early detection of areas vulnerable to wildfires in the Eastern Mediterranean region. Euro-Mediterr. J. Environ. Integr. 2020, 5, 1–13. [Google Scholar] [CrossRef]
- Vitelli, M.; Vessella, F.; Cardoni, S.; Pollegioni, P.; Denk, T.; Grimm, G.W.; Simeone, M.C. Phylogeographic structuring of plastome diversity in Mediterranean oaks (Quercus Group Ilex, Fagaceae). Tree Genet. Genomes 2017, 13, 1–17. [Google Scholar] [CrossRef]
- Greuter, W.; Burdet, H.M.; Long, G. Med-Checklist; Conservatoire et Jardin botaniques de la Ville de Genève: Geneva, Switzerland, 1989; Volume 4, Available online: http://ww2.bgbm.org/mcl/ (accessed on 27 January 2021).
- Duggen, S.; Hoernle, K.; van den Bogaard, P.; Rupke, L.; Morgan, J.P. Deep roots of the Messinian salinity crisis. Nature 2003, 422, 602–606. [Google Scholar] [CrossRef] [PubMed]
- Pardo, C.; Cubas, P.; Tahiri, H. Genetic variation and phylogeography of Stauracanthus (Fabaceae, Genisteae) from the Iberian Peninsula and northern Morocco assessed by chloroplast microsatellite (cpSSR) markers. Am. J. Bot. 2008, 95, 98–109. [Google Scholar] [CrossRef] [PubMed]
- Jabbour, F.; Renner, S.S. Consolida and Aconitella are an annual clade of Delphinium (Ranunculaceae) that diversified in the Mediterranean basin and the Irano-Turanian region. Taxon 2011, 60, 1029–1040. [Google Scholar] [CrossRef]
- Lledó, M.D.; Crespo, M.B.; Fay, M.F.; Chase, M.W. Molecular phylogenetics of Limonium and related genera (Plumbaginaceae): Biogeographical and systematic implications. Am. J. Bot. 2005, 92, 1189–1198. [Google Scholar] [CrossRef] [Green Version]
- Suc, J.P.; Popescu, S.M. Pollen records and climatic cycles in the North Mediterranean region since 2.7 Ma. Geol. Soc. Lond. Spec. Publ. 2005, 247, 147–158. [Google Scholar] [CrossRef]
- Rosselló, J.A.; Lázaro, A.; Cosín, R.; Molins, A. A phylogeographic split in Buxus balearica (Buxaceae) as evidenced by nuclear ribosomal markers: When ITS paralogues are welcome. J. Mol. Evol. 2007, 64, 143–157. [Google Scholar] [CrossRef]
- Rosselló, J.A.; Cosín, R.; Bacchetta, G.; Brullo, S.; Mayol, M. Nuclear and chloroplast DNA variation in Cephalaria squamiflora (Dipsacaceae), a disjunct Mediterranean species. Taxon 2009, 58, 1242–1253. [Google Scholar] [CrossRef]
- Kadereit, G.; Yaprak, A.E. Microcnemum coralloides (Chenopodiaceae-Salicornioideae): An example of intraspecific East–West disjunctions in the Mediterranean region. An. Jard. Bot. Madr. 2008, 65, 415–426. [Google Scholar] [CrossRef] [Green Version]
- Casimiro-Soriguer, R.; Talavera, M.; Balao, F.; Terrab, A.; Herrera, J.; Talavera, S. Phylogeny and genetic structure of Erophaca (Leguminosae), a East–West Mediterranean disjunct genus from the Tertiary. Mol. Phylogenet. Evol. 2010, 56, 441–450. [Google Scholar] [CrossRef] [PubMed]
- Kropf, M.; Kadereit, J.W.; Comes, H.P. Late Quaternary distributional stasis in the submediterranean mountain plant Anthyllis montana L. (Fabaceae) inferred from ITS sequences and amplified fragment length polymorphism markers. Mol. Ecol. 2002, 11, 447–463. [Google Scholar] [CrossRef] [PubMed]
- Kropf, M. Intraspecific patterns of European mountain plants: A morphometric analysis confirms molecular results in the submediterranean oreophyte Anthyllis montana L. (Fabaceae). Taxon 2008, 57, 511–524. [Google Scholar] [CrossRef]
- García-Castaño, J.L.; Terrab, A.; Ortiz, M.A.; Stuessy, T.F.; Talavera, S. Patterns of phylogeography and vicariance of Chamaerops humilis L. (Palmae). Turk. J. Bot. 2014, 38, 1132–1146. [Google Scholar] [CrossRef]
- Hardion, L.; Dumas, P.J.; Abdel-Samad, F.; Kharrat, M.B.D.; Surina, B.; Affre, L.; Médail, F.; Bacchetta, G.; Baumel, A. Geographical isolation caused the diversification of the Mediterranean thorny cushion-like Astragalus L. sect. Tragacantha DC. (Fabaceae). Mol. Phylogenet. Evol. 2016, 97, 187–195. [Google Scholar] [CrossRef]
- Ackerly, D.D. Evolution, origin and age of lineages in the Californian and Mediterranean floras. J. Biogeogr. 2009, 36, 1221–1233. [Google Scholar] [CrossRef]
- Cardona, M.A.; Llorens, L.; Sierra, E. Étude biosystématique de Dorycnium pentaphyllum Scop. subsp. fulgurans (Porta) comb. nova, endémique des Baléares Orientales. Collect. Bot. 1983, 14, 133–150. [Google Scholar]
- Cardona, M.A.; Contandriopoulos, J.; Ràfols, E.S. Étude biosystématique d’Anthyllis hystrix de Minorque et d’A. hermanniae de la Méditerranée orientale et centrale. Orsis 1986, 2, 5–25. [Google Scholar]
- Heyn, C.C. Lotus L. In Flora of Turkey and the East Aegean Islands; Davis, P.H., Ed.; Cambridge University Press: Cambridge, UK, 1970; Volume 3, pp. 518–531. [Google Scholar]
- Kuzmanov, B. Dorycnium Mill., Lotus L. In Flora Reipublicae Popularis Bulgaricae; Jordanov, D., Ed.; Aedibus Academiae Scientiarum Bulgaricae: Serdicae, Bulgaria, 1976; Volume 6, pp. 197–214. (In Bulgarian) [Google Scholar]
- Kurbatskij, V.I. Lotus L. In Flora Sibiriae: Fabaceae (Leguminosae); Polozhij, A.V., Malyschev, L.I., Eds.; Nauka: Novosibirsk, Russia, 1994; Volume 9, pp. 168–170. (In Russian) [Google Scholar]
- Grossheim, A.A. Dicotyledons (Saururaceae—Leguminosae). In Flora of the Caucasus; USSR: Tiflis, Georgia, 1930; Volume 2, p. 438. (In Russian) [Google Scholar]
- Avetisyan, Z.E. Lotus L. In Flora of Armenia; Takhtajian, A.A., Ed.; Acad. Sci. USSR: Erevan, Armenia, 1962; Volume 4, pp. 90–93. (In Russian) [Google Scholar]
- Hayek von, A. Prodromus florae peninsulae Balcanicae 1. Repert. Spec. Nov. Regn. Veg. 1926, 30, 673–960. [Google Scholar]
- Greuter, W.; Raus, T. Med-Checklist Notulae, 13. Willdenowia 1986, 16, 103–116. [Google Scholar]
- Chrtková-Žertová, A. Lotus. In Flora Iranica; Rechinger, K.H., Ed.; Akademische Druck und Verlagsanstalt: Graz, Austria, 1984; pp. 327–342. [Google Scholar]
- Goloskokov, V.P. Lotus. In Flora Kazakhstana; Pavlov, N.V., Ed.; Izdatel’stvo Akademii Nauk Kazakhskoi SSR: Alma-Ata, Kazakhstan, 1961; Volume 5, pp. 62–65. (In Russian) [Google Scholar]
- Harzhauser, M.; Piller, W.E. Benchmark data of a changing sea—palaeogeography, palaeobiogeography and events in the Central Paratethys during the Miocene. Palaeogeogr. Palaeocl. 2007, 253, 8–31. [Google Scholar] [CrossRef]
- Demiriz, H. Dorycnium Miller. In Flora of Turkey and the East Aegean Islands; Davis, P.H., Ed.; Cambridge University Press: Cambridge, UK, 1970; Volume 3, pp. 512–518. [Google Scholar]
- Díaz Lifante, Z. Dorycnium Mill. In Flora Iberica; Real Jardín Botaníco: Madrid, Spain, 2000; Volume 7, pp. 812–823. [Google Scholar]
- Galushko, A.I. Flora Severnogo Kavkaza. Opredelitel; Izdatelstvo Rostovskogo Universiteta: Rostov-na-Donu, Russia, 1980; Volume 2, pp. 1–352. (In Russian) [Google Scholar]
- Subaşı, Ü.; Şekerciler, F.; Vural, M. Conservation assessment and action on Lotus sanguineus (Fabaceae), a critically endangered endemic species to Turkey. Hacquetia 2020, 19, 307–315. [Google Scholar] [CrossRef]
- Shorthouse, D.P. SimpleMappr, an Online Tool to Produce Publication-Quality Point Maps. 2010. Available online: http://www.simplemappr.net (accessed on 12 January 2021).
- Doyle, J.J.; Doyle, J.L. A rapid DNA isolation procedure for small quantities of fresh leaf material. Phytochem. Bull. Bot. Soc. Am. 1987, 19, 11–15. [Google Scholar]
- Wen, J.; Zimmer, E. Phylogeny and biogeography of Panax L. (the ginseng genus, Araliaceae): Inferences from ITS sequences of nuclear ribosomal DNA. Mol. Phylogenet. Evol. 1996, 6, 167–177. [Google Scholar] [CrossRef] [PubMed]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Innis, M.A., Gelfand, D.H., Sninsky, J.J., White, T.J., Eds.; Academic Press: San Diego, CA, USA, 1990; pp. 315–322. [Google Scholar]
- Taberlet, P.; Gielly, L.; Pautou, G.; Bouvet, J. Universal primers for amplification of three non-coding regions of chloroplast DNA. Plant Mol. Biol. 1991, 17, 1105–1109. [Google Scholar] [CrossRef]
- Oxelman, B.; Lidén, M.; Berglund, D. Chloroplast rps16 intron phylogeny of the tribe Sileneae (Caryophyllaceae). Plant Syst. Evol. 1997, 206, 393–410. [Google Scholar] [CrossRef] [Green Version]
- Bohley, K.; Joos, O.; Hartmann, H.; Sage, R.; Liede-Schumann, S.; Kadereit, G. Phylogeny of Sesuvioideae (Aizoaceae)—Biogeography, leaf anatomy and the evolution of C4 photosynthesis. Perspect. Plant Ecol. 2015, 17, 116–130. [Google Scholar] [CrossRef]
- Ye, J.; Coulouris, G.; Zaretskaya, I.; Cutcutache, I.; Rozen, S.; Madden, T.L. Primer-BLAST: A tool to design target-specific primers for polymerase chain reaction. BMC Bioinf. 2012, 13, 134. [Google Scholar] [CrossRef] [Green Version]
- Katoh, K.; Misawa, K.; Kuma, K.; Miyata, T. MAFFT: A novel method for rapid multiple sequence alignment based on fast Fourier transform. Nucleic Acids Res. 2002, 30, 3059–3066. [Google Scholar] [CrossRef] [Green Version]
- Katoh, K.; Standley, D.M. MAFFT Multiple Sequence Alignment Software Version 7: Improvements in Performance and Usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hall, T.A. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef] [PubMed]
- Ronquist, F.; Teslenko, M.; van der Mark, M.; Ayres, D.; Darling, A.; Höhna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MrBayes 3.2: Efficient Bayesian phylogenetic inference and model choice across a large model space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Swofford, D.L. PAUP*. Phylogenetic Analysis Using Parsimony (*and Other Methods); Sinauer Associates: Sunderland, UK, 2003; Version 4. [Google Scholar]
- Ali, R.H.; Bark, M.; Miró, J.; Muhammad, S.A.; Sjöstrand, J.; Zubair, S.M.; Abbas, R.M.; Arvestad, L. VMCMC: A graphical and statistical analysis tool for Markov chain Monte Carlo traces. BMC Bioinf. 2017, 18, 97. [Google Scholar] [CrossRef] [Green Version]
- Clement, M.; Posada, D.; Crandall, K.A. TCS: A computer program to estimate gene genealogies. Mol. Ecol. 2000, 9, 1657–1659. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Santos dos, A.M.; Cabezas, M.P.; Tavares, A.I.; Xavier, R.; Branco, M. tcsBU: A tool to extend TCS network layout and visualization. Bioinformatics 2016, 32, 627–628. [Google Scholar] [CrossRef] [Green Version]
- Rozas, J.; Ferrer-Mata, A.; Sánchez-DelBarrio, J.C.; Guirao-Rico, S.; Librado, P.; Ramos-Onsins, S.E.; Sánchez-Gracia, A. DnaSP 6: DNA sequence polymorphism analysis of large datasets. Mol. Biol. Evol. 2017, 34, 3299–3302. [Google Scholar] [CrossRef]
- Bouckaert, R.; Vaughan, T.G.; Barido-Sottani, J.; Duchêne, S.; Fourment, M.; Gavryushkina, A.; Heled, J.; Jones, G.; Kühnert, D.; De Maio, N.; et al. BEAST 2.5: An advanced software platform for Bayesian evolutionary analysis. PLoS Comput. Biol. 2019, 15, e1006650. [Google Scholar] [CrossRef] [Green Version]
- Drummond, A.J.; Ho, S.Y.W.; Phillips, M.J.; Rambaut, A. Relaxed phylogenetics and dating with confidence. PLoS Biol. 2006, 4, e88. [Google Scholar] [CrossRef]
- Gernhard, T. The conditioned reconstructed process. J. Theor. Biol. 2008, 253, 769–778. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maturana, P.M.; Brewer, B.J.; Klaere, S.; Bouckaert, R.R. Model selection and parameter inference in phylogenetics using Nested Sampling. Syst. Biol. 2019, 68, 219–233. [Google Scholar] [CrossRef]
- Rambaut, A.; Drummond, A.J.; Xie, D.; Baele, G.; Suchard, M.A. Posterior summarisation in Bayesian phylogenetics using Tracer 1.7. Syst. Biol. 2018, 67, 901–904. [Google Scholar] [CrossRef] [PubMed] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Taxa | Lotus dorycnium, L. herbaceus, L. germanicus, L. fulgurans, L. graecus, L. axilliflorus, L. sanguineus (Species Formerly Considered as Typical Members of Dorycnium) | L. rectus | L. hirsutus | L. strictus | Other Species of Lotus | Outgroups |
|---|---|---|---|---|---|---|
| Taxonomic placement in Sokoloff (2003b) and Degtjareva et al. (2006) | Lotus section Dorycnium | Lotus section Bonjeanea | Lotus sections Benedictella, Chamaelotus, Erythrolotus, Heinekenia, Krokeria, Lotea, Lotus, Ononidium, Pedrosia, Rhyncholotus, Tetragonolobus | Tripodion, Hammatolobium, Cytisopsis | ||
| Leaf rachis | Almost always absent | Present | Present or absent | Present or absent | Usually present, but absent in some species of the Southern Clade | Present or absent |
| Position of sterile bract (see Sokoloff et al., 2007) | Often separated from partial inflorescence by a stalk | Often separated from partial inflorescence by a stalk | Often separated from partial inflorescence by a stalk | At the base of partial inflorescence | At the base of partial inflorescence | At the base of partial inflorescence (Hammatolobium, Tripodion) or absent (Cytisopsis) |
| Flowers and partial inflorescences | Partial inflorescences more commonly many-flowered (6–30 flowers often arranged on inflorescence axis in more than one whorl) | Partial inflorescences commonly few-flowered (a whorl of 1–8 flowers). | ||||
| Flower size | <7 mm | <7 mm | >7 mm | >7 mm | > or <7 mm | >7 mm |
| Petal colour | Never yellow | Never yellow | Never yellow | Yellow to white | Yellow or not yellow | Yellow or not yellow |
| Keel shape | Obtuse | Obtuse | Rostrate | Rotrate | Rostrate | Rostrate or obtuse |
| Distal parts of wing petals | Adhering together | Free | Free | Free | Free or (in some members of the Southern Clade) adhering together | Free |
| Outgrowths in distal parts of wing petals | Present | Absent | Absent | Absent | Absent | Absent |
| Style surface | Smooth | Smooth | Smooth | Smooth | Papillose | Smooth |
| Fruits | 1-seeded (few-seeded in L. graecus) | Many-seeded | Many-seeded | Many-seeded | Usually many-seeded | Two- to many-seeded |
| Brown cells in endocarp | Usually present | Present | Present | Present | Absent | Present or absent |
| Node | nrITS Node Defined as MRCA or Crown Clade of | Mean Age (Ma) | CI Values (95% HPD) |
|---|---|---|---|
| A | Loteae | 29.11 | 18.286–34.82 |
| B | Lotus + Cytisopsis + Tripodion | 20.71 | 12.907–25.643 |
| C | Lotus | 15.87 | 9.995–19.813 |
| D | Lotus Northern clade 1 | 12.47 | 7.526–15.784 |
| E | Lotus Northern clade 2 | 12.4 | 7.228–16.036 |
| F | Lotus Southern clade | 10.21 | 5.827–13.356 |
| G | Lotus section Dorycnium (incl. L. hirsutus) | 9.83 | 5.447–13.151 |
| H | L. sanguineus + L. graecus + L. axilliflorus | 6.38 | 2.715–9.641 |
| I | L. graecus + L. axilliflorus | 2.81 | 0.776–5.042 |
| J | L. hirsutus + L. dorycnium | 5.55 | 2.916–8.143 |
| K | L. fulgugans + L. germanicus | 1.61 | 0.489–2.831 |
| L | L. hirsutus | 4.16 | 1.731–6.412 |
| M | L. rectus + L. strictus | 10.58 | 6.124–13.964 |
| N | L. rectus | 4.94 | 2.078–7.603 |
| O | L. strictus | 6.1 | 2.815–8.768 |
| BD | Yule | |
|---|---|---|
| relaxed clock | −7391.169 ± 6.251 | −7386.553 ± 6.170 |
| strict clock | −7415.723 ± 8.205 | −7422.717 ± 8.211 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kramina, T.E.; Lysova, M.V.; Samigullin, T.H.; Schanzer, I.A.; Özbek, M.U.; Sokoloff, D.D. Phylogenetic Placement and Phylogeography of Large-Flowered Lotus Species (Leguminosae) Formerly Classified in Dorycnium: Evidence of Pre-Pleistocene Differentiation of Western and Eastern Intraspecific Groups. Plants 2021, 10, 260. https://doi.org/10.3390/plants10020260
Kramina TE, Lysova MV, Samigullin TH, Schanzer IA, Özbek MU, Sokoloff DD. Phylogenetic Placement and Phylogeography of Large-Flowered Lotus Species (Leguminosae) Formerly Classified in Dorycnium: Evidence of Pre-Pleistocene Differentiation of Western and Eastern Intraspecific Groups. Plants. 2021; 10(2):260. https://doi.org/10.3390/plants10020260
Chicago/Turabian StyleKramina, Tatiana E., Maya V. Lysova, Tahir H. Samigullin, Ivan A. Schanzer, Mehmet U. Özbek, and Dmitry D. Sokoloff. 2021. "Phylogenetic Placement and Phylogeography of Large-Flowered Lotus Species (Leguminosae) Formerly Classified in Dorycnium: Evidence of Pre-Pleistocene Differentiation of Western and Eastern Intraspecific Groups" Plants 10, no. 2: 260. https://doi.org/10.3390/plants10020260