
New Aspects of Secretory Structures in Five Alismataceae Species: Laticifers or Ducts?

 , ,
, ,
Abstract
:1. Introduction
2. Results
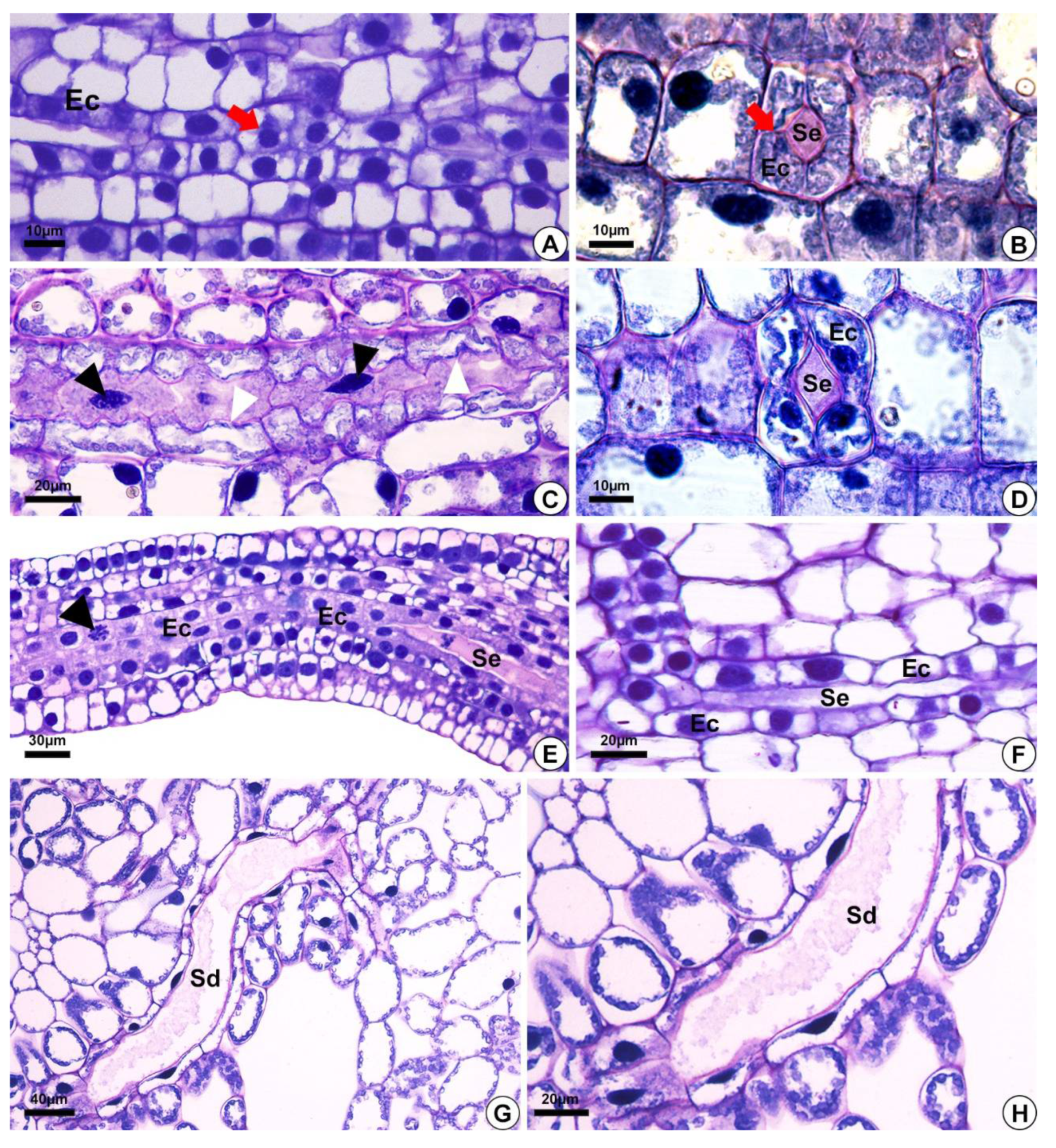
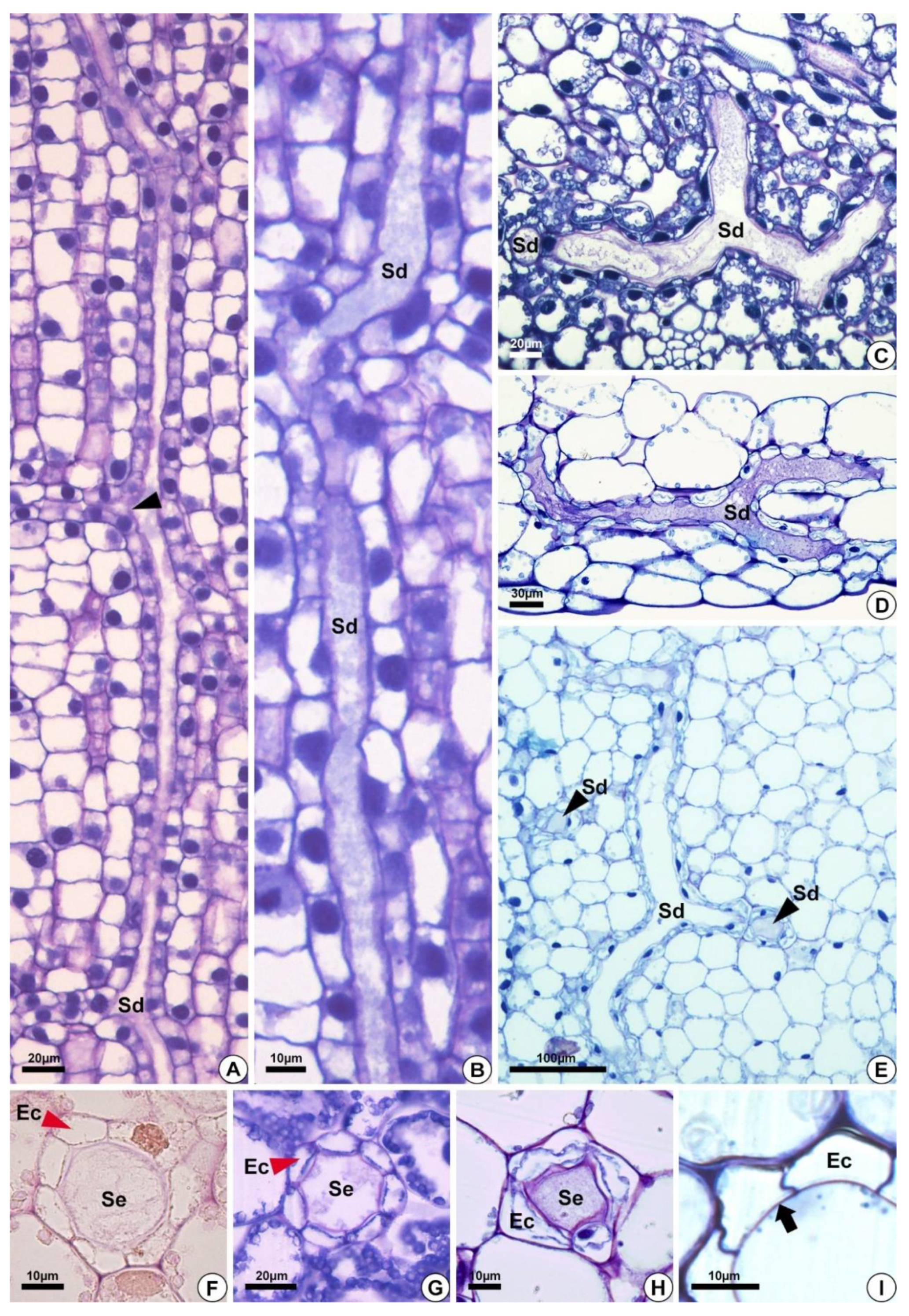
2.1. Origin and Morphology of the Secretory Structures
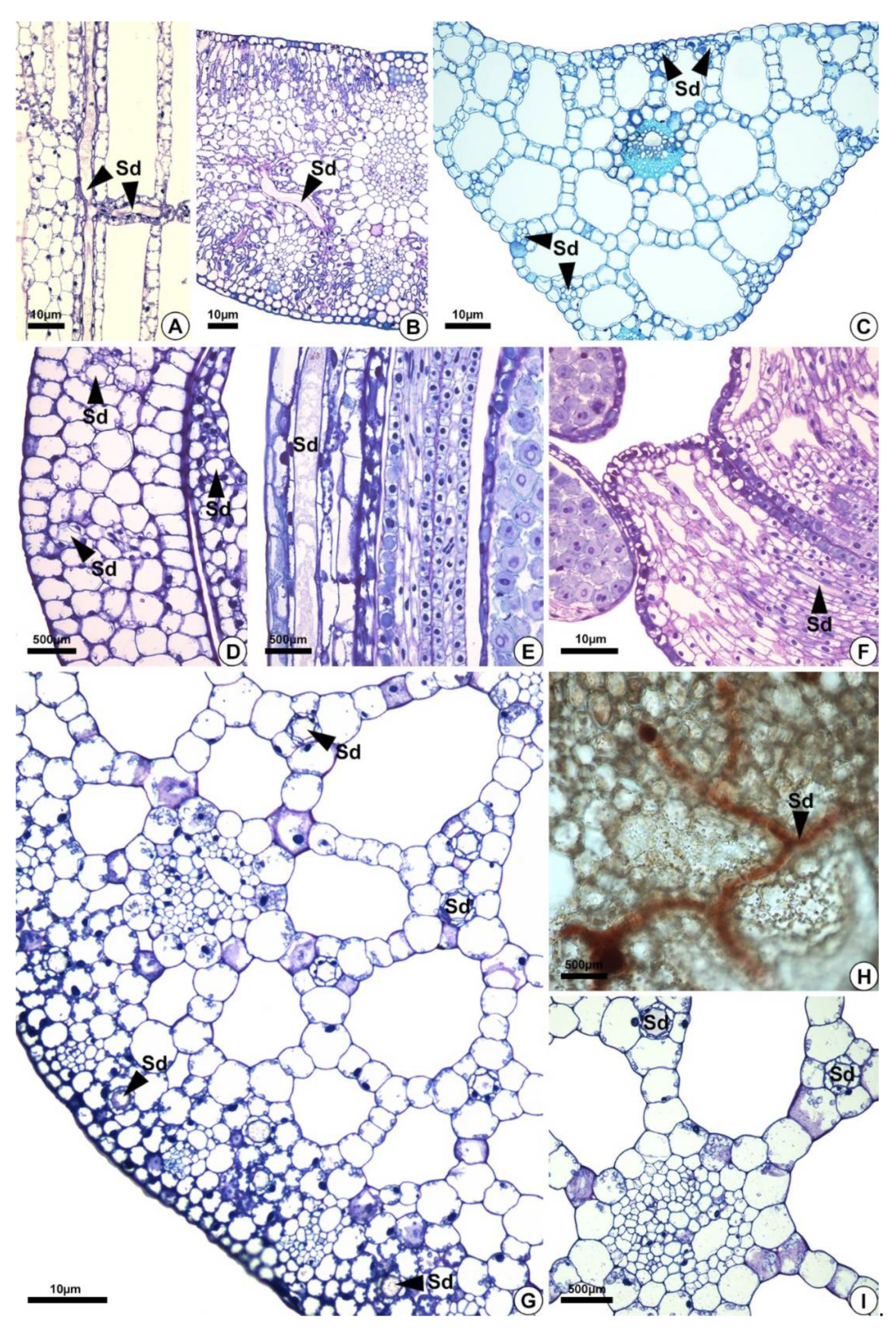
2.2. Distribution of the Secretory Ducts in the Plant
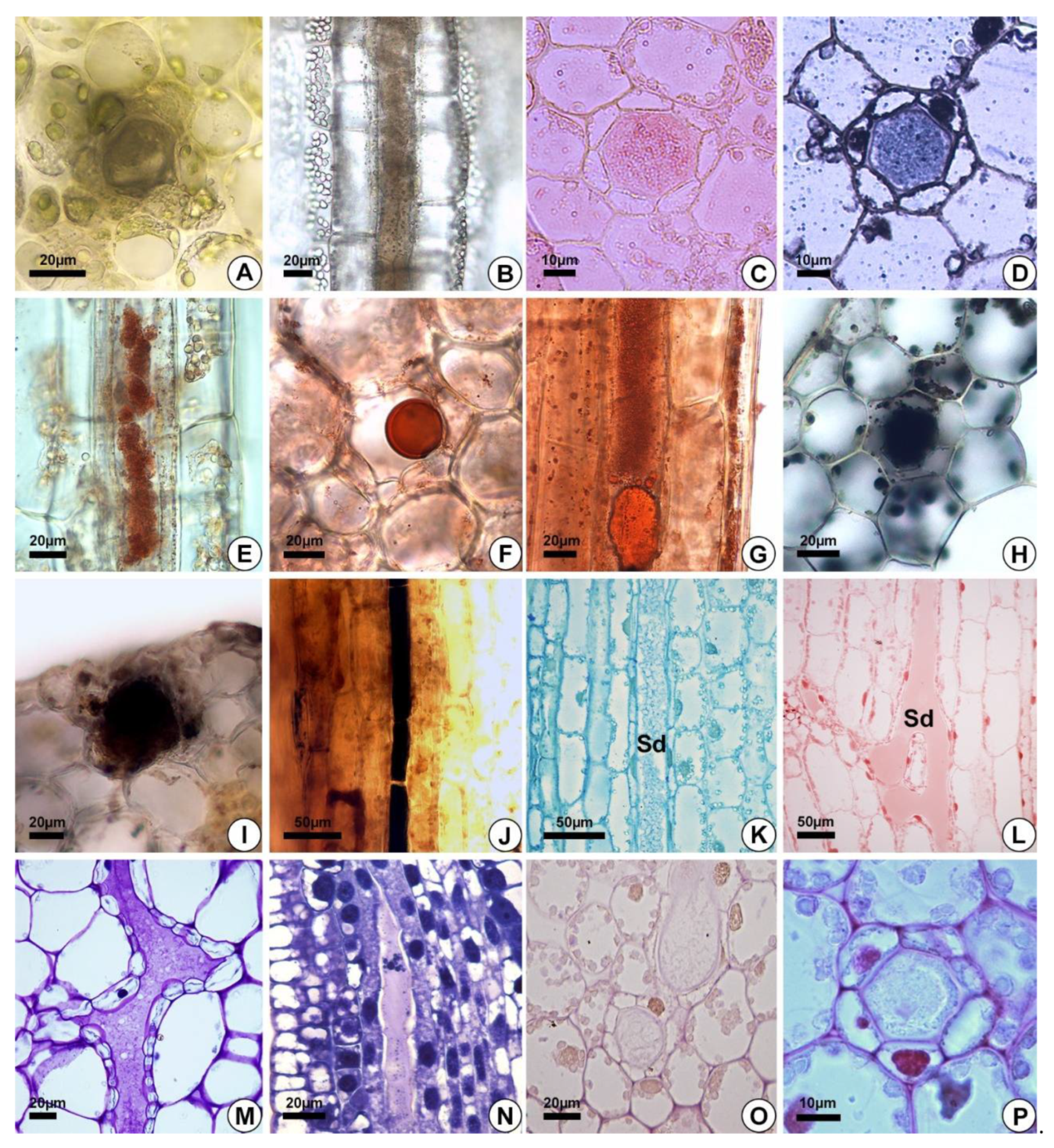
2.3. Secretion Composition
3. Discussion
3.1. Resin Duct Structure
3.2. Secretion Composition and Functions
3.3. Taxonomic Implications
4. Materials and Methods
4.1. Plant Material
4.2. Histological Analysis
4.3. Histochemical Analysis
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Haynes, R.R.; Les, D.H.; Holm-Nielsen, L.B. Alismataceae. In The Families and Genera of Vascular Plants; Kubitzki, K., Ed.; Springer: Heidelberg, Germany, 1998; Volume 4, pp. 11–18. [Google Scholar]
- APG (Angiosperm Phylogeny Group) III. An update of the Angiosperm Phylogeny Group classification for the orders and families of flowering plants: APG III. Bot. J. Linn. Soc. 2009, 161, 105–121. [Google Scholar] [CrossRef] [Green Version]
- Chen, L.Y.; Chen, J.M.; Gituru, R.W.; Temam, T.D.; Wang, Q.F. Generic phylogeny and historical biogeography of Alismataceae, inferred from multiple DNA sequences. Mol. Phylogenet. Evol. 2012, 63, 407–416. [Google Scholar] [CrossRef] [PubMed]
- The Plant List. Available online: http://www.theplantlist.org/ (accessed on 14 October 2021).
- Lehtonen, S. Systematics of the Alismataceae—A morphological evaluation. Aquat. Bot. 2009, 91, 279–290. [Google Scholar] [CrossRef]
- Pott, V.J.; Pott, A. Plantas Aquáticas do Pantanal; Embrapa: Brasília, Brazil, 2000; pp. 80–92. [Google Scholar]
- Bercu, R. Histoanatomical features of the aquatic plant Helanthium tenellum (Mart.) Britt. (Alismataceae). Ann. West Univ. Timiş. Ser. Biol. 2015, 18, 67–72. [Google Scholar]
- ANVISA (Agência Nacional de Vigilância Sanitária). Farmacopéia Brasileira, 6th ed.; 2019; Volume 1. Available online: https://www.gov.br/anvisa/pt-br/assuntos/farmacopeia/farmacopeia-brasileira/arquivos/7989json-file-1 (accessed on 30 January 2021).
- Dias, E.G.E.; Valenzuela, V.C.T.; Alves, M.R.; Duarte, M.G.R.; Garcia, E.F. Qualidade e autenticidade de folhas de chapéu-de-couro (Echinodorus grandiflorus) oriundas de fornecedores de São Paulo. Rev. Bras. Plantas Med. 2013, 15, 250–256. [Google Scholar] [CrossRef] [Green Version]
- Marques, A.; Provance, D.W., Jr.; Kaplan, M.A.C.; Figueiredo, M.R. Echinodorus grandiflorus: Ethnobotanical, phytochemical and pharmacological overview of a medicinal plant used in Brazil. Food Chem. Toxicol. 2017, 109, 1032–1047. [Google Scholar] [CrossRef]
- Oliveira, D.P.; Braga, F.C.; Teixeira, M.M. Medicinal plants and their potential use in the treatment of rheumatic diseases. In Inflammation and Natural Products, 1st ed.; Gopi, S., Amalraj, A., Kunnumakkara, A., Thomas, S., Eds.; Academic Press: Cambridge, MA, USA, 2021; pp. 205–234. [Google Scholar]
- Dutra, R.C.; Tavares, C.Z.; Ferraz, S.O.; Sousa, O.V.; Pimenta, D.S. Investigação das atividades analgésica e antiinflamatória do extrato metanólico dos rizomas de Echinodorus grandiflorus. Braz. J. Pharm. 2006, 16, 469–474. [Google Scholar] [CrossRef]
- Govindarajalu, E. Further contribution to the anatomy of the alismataceae: Sagittaria guayanensis H.B.K. ssp. lappula (D.Don) bogin. Proc. Indian Acad. Sci. 1967, 65, 142–152. [Google Scholar] [CrossRef]
- Prado, E.; Demarco, D. Laticifers and Secretory Ducts: Similarities and Differences. In Ecosystem Services and Global Ecology; Hufnagel, L., Ed.; IntechOpen: London, UK, 2018; pp. 103–123. [Google Scholar]
- Gunawardena, A.; Greenwood, J.S.; Dengler, N.G. Cell wall degradation and modification during programmed cell death in lace plant, Aponogeton madagascariensis (Aponogetonaceae). Am. J. Bot. 2007, 94, 1116–1128. [Google Scholar] [CrossRef] [Green Version]
- French, J.C. Systematic occurrence of anastomosing laticifers in Araceae. Bot. Gaz. 1988, 149, 71–81. [Google Scholar] [CrossRef]
- Keating, R.C. Leaf Anatomical Characters and Their Value in Understanding Morphoclines in the Araceae. Bot. Rev. 2003, 68, 510–523. [Google Scholar] [CrossRef]
- Tomlinson, P.B. VII. Helobiae (Alismatidae). In Anatomy of the Monocotyledons; Metcalfe, C.R., Ed.; Clarendon Press: Oxford, UK, 1982; p. 559. [Google Scholar]
- Stant, M.Y. Anatomy of the Alismataceae. J. Linn. Soc. 1963, 59, 1–42. [Google Scholar] [CrossRef]
- Judd, W.S.; Campbell, C.S.; Kellogg, E.A.; Stevens, P.F.; Donoghue, M.J. Sistemática Vegetal: Um Enfoque Filogenético, 3rd ed.; Artmed: Porto Alegre, Brazil, 2009; p. 632. [Google Scholar]
- Dahlgren, R.M.T.; Clifford, H.T.; Yeo, P.F. Evolution within the monocotyledons. In Flowering Plants. Monocotyledon. The Families and Genera of Vascular Plants; Kubitzki, K., Ed.; Springer: Heidelberg, Germany, 1998; Volume 4, pp. 91–106. [Google Scholar]
- Colares, M.N.; Delucchi, G.; Nova, M.C.; Vizcaíno, C.E. Anatomia y etnobotanica de las especies medicinales de monocotiledoneas de la estepa pampeana: Alismataceae, Araceae y Arecaceae. Acta Farm. Bonaer. 1997, 16, 137–143. [Google Scholar]
- Matias, L.Q.; Soares, A.; Scatena, V.L. Systematic consideration of petiole anatomy of species of Echinodorus Richard (Alismataceae) from north-eastern Brazil. Flora 2007, 202, 395–402. [Google Scholar] [CrossRef]
- Matias, L.Q.; Soares, A.; Scatena, V.L. Anatomy of Echinodorus (Alismataceae) scapes from Northeastern Brazil as applied to taxonomy. Edinb. J. Bot. 2008, 65, 11–21. [Google Scholar] [CrossRef] [Green Version]
- Fahn, A. Secretory Tissues in Plants; Academic Press: London, UK, 1979. [Google Scholar]
- Fahn, A. Plant Anatomy, 4th ed.; Pergamon Press: Oxford, UK, 1990. [Google Scholar]
- Evert, R.F.; Eichhorn, S.E. Esau’s Plant Anatomy: Meristems, Cells, and Tissues of the Plant Body: Their Structure, Function, and Development, 3rd ed.; John Wiley & Sons Inc.: Hoboken, NJ, USA, 2006. [Google Scholar]
- Pickard, W.F. Research review. Laticifers and secretory ducts: Two other tube systems in plants. New Phytol. 2008, 177, 877–888. [Google Scholar] [CrossRef] [PubMed]
- Alencar, A.C.; Tölke, E.D.; Mayer, J.L.S. New perspectives on secretory structures in Clusia (Clusiaceae—Clusiod clade): Production of latex or resins? Botany 2020, 98, 161–172. [Google Scholar] [CrossRef]
- Teixeira, S.P.; Marinho, C.R.; Leme, F.M. Structural diversity and distribution of laticifers. In Advances in Botanical Research: Latex, Laticifers and Their Molecular Components: From Functions to Possible Applications; Nawrot, R., Ed.; Academic Press; Elsevier: London, UK, 2020; Volume 93, pp. 27–53. [Google Scholar]
- Ramos, M.V.; Freitas, C.D.T.; Morais, F.S.; Prado, E.; Medina, M.C.; Demarco, D. Plant latex and latex-borne defense. In Advances in Botanical Research: Latex, Laticifers and Their Molecular Components: From Functions to Possible Applications; Nawrot, R., Ed.; Academic Press: London, UK, 2020; Volume 93, pp. 1–25. [Google Scholar]
- Demarco, D.; Castro, M.M.; Ascensão, L. Two laticifer systems in Sapium haematospermum—New records for Euphorbiaceae. Botany 2013, 91, 545–554. [Google Scholar] [CrossRef]
- Demarco, D.; Kinoshita, L.S.; Castro, M.M. Laticíferos articulados anastomosados—Novos registros para Apocynaceae. Rev. Bras. Bot. 2006, 29, 133–144. [Google Scholar] [CrossRef] [Green Version]
- Leme, F.M.; Borella, P.H.; Marinho, C.R.; Teixeira, S.P. Expanding the laticifer knowledge in Cannabaceae: Distribution, morphology, origin, and latex composition. Protoplasma 2020, 257, 1183–1199. [Google Scholar] [CrossRef] [PubMed]
- Jacob, J.L.; D’Auzac, J.; Prevôt, J.C. The Composition of Natural Latex from Hevea brasiliensis. Clin. Rev. Allergy Immunol. 1993, 11, 225–337. [Google Scholar]
- Endress, M.E.; Bruyns, P.V. A Revised Classification of Apocynaceae s.l. Bot. Rev. 2000, 66, 1–56. [Google Scholar] [CrossRef] [Green Version]
- Ramos, M.V.; Demarco, D.; Costa-Souza, I.C.; Freitas, C.D.T. Laticifers, latex, and their role in plant defense. Trends Plant Sci. 2019, 24, 553–567. [Google Scholar] [CrossRef] [PubMed]
- Costa, E.R.; Tangerina, M.M.; Ferreira, M.J.; Demarco, D. Two Origins, Two Functions: The Discovery of Distinct Secretory Ducts Formed during the Primary and Secondary Growth in Kielmeyera. Plants 2021, 10, 877. [Google Scholar] [CrossRef] [PubMed]
- Hagel, J.M.; Yeung, E.C.; Facchini, P.J. Got milk? The secret life of laticifers. Trends Plant Sci. 2008, 13, 631–639. [Google Scholar] [CrossRef]
- Bercu, R. Structural aspects of Alisma plantago—Aquatica L. (Alismataceae). Ann. West Univ. Timiş. Ser. Biol. 2017, 20, 179–184. [Google Scholar]
- Leite, J.P.V.; Pimenta, D.S.; Gomes, R.S.; Dantas-Barros, A.M. Contribuição ao estudo farmacobotânico da Echinodorus macrophyllus (Kunth) Micheli (chapéu-de-couro)—Alismataceae. Rev. Bras. Farmacogn. 2007, 17, 242–248. [Google Scholar] [CrossRef] [Green Version]
- Metcalfe, C.R. Recent work on the systematic anatomy of the monocotyledons (with special reference to investigations at the Jodrell Laboratory at Kew). Kew Bull. 1954, 9, 523–532. [Google Scholar] [CrossRef]
- Souza, L.R.; Trindade, F.G.; Oliveira, R.A.; Costa, L.C.D.B.; Moreira, V.G.; Da Cunha, M. Histochemical characterization of secretory ducts and essential oil analysis of Protium species (Burseraceae). J. Essent. Oil Res. 2015, 28, 166–171. [Google Scholar] [CrossRef]
- Silva, K.M.M.; Luna, B.N.; Joffily, A.; Paiva, S.R.; Barros, C.F. Revealing the development of secretory structures in the leaves of Clusia fluminensis and Clusia lanceolata (Clusiaceae). Flora 2019, 256, 69–78. [Google Scholar] [CrossRef]
- Tolke, E.D.; Lacchia, A.P.S.; Lima, E.A.; Demarco, D.; Ascensão, L.; Carmello-Guerreiro, S.M. Secretory ducts in Anacardiaceae revisited: Updated concepts and new findings based on histochemical evidence. S. Afr. J. Bot. 2021, 138, 394–405. [Google Scholar] [CrossRef]
- Ciccarelli, D.; Andreucci, A.C.; Pagni, A.M. Translucent glands and secretory canals in Hypericum perforatum L. (Hypericaceae): Morphological, anatomical and histochemical studies during the course of ontogenesis. Ann. Bot. 2001, 88, 637–644. [Google Scholar] [CrossRef] [Green Version]
- Gonçalves-Souza, P.; Schlindwein, C.; Paiva, E.A.S. Floral resins of Philodendron adamantinum (Araceae): Secretion, release and synchrony with pollinators. Acta Bot. Bras. 2018, 32, 392–401. [Google Scholar] [CrossRef]
- Pessoa, M.J.G.; Pireda, S.; Simioni, P.; Bautz, N.; Da Cunha, M. Structural and histochemical attributes of secretory ducts and cavities in leaves of four species of Calophyllaceae J. Agardh in Amazonian savannas. Plant Biol. 2021, 23, 1128–1140. [Google Scholar] [CrossRef]
- Dorly, D.; Tjitrosemito, S.; Poerwanto, R.; Juliarni, J. Secretory Duct Structure and Phytochemistry Compounds of Yellow Latex in Mangosteen Fruit. HAYATI J. Biosci. 2008, 15, 99–104. [Google Scholar] [CrossRef] [Green Version]
- French, J.C. Systematic survey of resin canals in roots of Araceae. Bot. Gaz. 1987, 148, 360–371. [Google Scholar] [CrossRef]
- Garcia, T.B.; Costa, E.R.; Kikuchi, T.Y.S.; Aguiar-Dias, A.C.A.; Demarco, D. Coalescent cavities: A novel process of secretory duct formation in Theobroma L. (Malvaceae). In Plant Ontogeny: Studies, Analyses and Evolutionary Implications; Demarco, D., Ed.; Nova: New York, NY, USA, 2020; pp. 1–20. [Google Scholar]
- Venning, F.D. The ontogeny of the laticiferous canals in the Anacardiaceae. Am. J. Bot. 1948, 35, 637–644. [Google Scholar] [CrossRef]
- Luo, D.; Xu, Y.Y. Preliminary study on the blocked laticiferous canals in the bark of Rhus verniciflua Stokes. Sci. Sin. 1984, 20, 418–422. [Google Scholar]
- Gleason, F.; Chollet, R. Plant Biochemistry, 1st ed.; Jones & Bartlett Learning: Sudbury, MA, USA, 2012. [Google Scholar]
- Esau, K. Plant Anatomy; John Wiley and Sons Inc.: New York, NY, USA, 1965. [Google Scholar]
- Agrawal, A.A.; Konno, K. Latex: A model for understanding mechanisms, ecology, and evolution of plant defense against herbivory. Annu. Rev. Ecol. Evol. Syst. 2009, 40, 311–331. [Google Scholar] [CrossRef] [Green Version]
- Cusimano, N.; Bogner, J.; Mayo, S.J.; Boyce, P.C.; Wong, S.Y.; Hesse, M.; Hetterscheid, W.L.A.; Keating, R.C.; French, J.C. Relationships within the Araceae: Comparison of morphological patterns with molecular phylogenies. Am. J. Bot. 2011, 98, 654–668. [Google Scholar] [CrossRef] [PubMed]
- Farrell, B.D.; Dussourd, D.E.; Mitter, C. Escalation of plant defense: Do latex and resin canals spur plant diversification? Am. Nat. 1991, 138, 881–900. [Google Scholar] [CrossRef]
- Ekchaweng, K.; Evangelisti, E.; Schornack, S.; Tian, M.; Churngchow, N. The plant defense and pathogen counter defense mediated by Hevea brasiliensis serine protease HbSPA and Phytophthora palmivora extracellular protease inhibitor PpEPI10. PLoS ONE 2017, 12, e0175795. [Google Scholar] [CrossRef]
- Amorin, A.; Borba, H.R.; Carauta, J.P.P.; Lopes, D.; Kaplan, M.A.C. Anthelmintic activity of the latex of Ficus species. J. Ethnopharmacol. 1999, 64, 255–258. [Google Scholar] [CrossRef]
- Lewinsohn, T.M. The geographical distribution of plant latex. Chemoecology 1991, 2, 64–68. [Google Scholar] [CrossRef]
- Kuo, J.; McComb, A.J. Posidoniaceae. In Flowering Plants·Monocotyledons. The Families and Genera of Vascular Plants; Kubitzki, K., Ed.; Springer: Heidelberg, Germany, 1998; pp. 404–408. [Google Scholar]
- Li, X.; Zhou, Z. Phylogenetic studies of the core Alismatales inferred from morphology and rbcL sequences. Prog. Nat. Sci. 2009, 19, 931–945. [Google Scholar] [CrossRef]
- Van Bruggen, H.W.E. Aponogetonaceae. In Flowering Plants·Monocotyledons. The Families and Genera of Vascular Plants; Kubitzki, K., Ed.; Springer: Heidelberg, Germany, 1998; pp. 21–25. [Google Scholar]
- CooK, C.D.K. Hydrocharitaceae. In Flowering Plants·Monocotyledons. The Families and Genera of Vascular Plants; Kubitzki, K., Ed.; Springer: Heidelberg, Germany, 1998; Volume 4, pp. 234–248. [Google Scholar]
- Lillie, R.D. Histopathologic Technic and Practical Histochemistry; McGraw-Hill Book Company: New York, NY, USA, 1965. [Google Scholar]
- Johansen, D.A. Plant Microtechnique; McGraw-Hill Book Company: New York, NY, USA, 1940. [Google Scholar]
- Marinho, C.R.; Teixeira, S.P. Novel reports of laticifers in Moraceae and Urticaceae: Revisiting synapomorphies. Plant Syst. Evol. 2019, 305, 13–31. [Google Scholar] [CrossRef]
- O’Brien, T.P.; Feder, N.; McCully, M.E. Polychromatic staining of plant cell walls by toluidine blue O. Protoplasma 1964, 59, 367–373. [Google Scholar] [CrossRef]
- Canaveze, Y.; Machado, S.R. The occurrence of intrusive growth associated with articulated laticifers in Tabernaemontana catharinensis A. DC., a new record for Apocynaceae. Int. J. Plant Sci. 2016, 177, 458–467. [Google Scholar] [CrossRef] [Green Version]
- Canaveze, Y.; Mastroberti, A.A.; Mariath, J.E.; Machado, S.R. Cytological differentiation and cell wall involvement in the growth mechanisms of articulated laticifers in Tabernaemontana catharinensis A. DC. (Apocynaceae). Protoplasma 2019, 256, 131–146. [Google Scholar] [CrossRef] [Green Version]
- Pearse, A.G.E. Histochemistry: Theoretical and Applied, 4th ed.; C. Livingstone: Edinburgh, UK, 1985; Volume 2. [Google Scholar]
- Ventrella, M.C.; Almeida, A.L.; Nery, L.A.; Coelho, V.P.M. Métodos Histoquímicos Aplicados às Sementes; Editora UFV: Viçosa, Brazil, 2013. [Google Scholar]
- Furr, M.; Mahlberg, P.G. Histochemical analyses of laticifers and glandular trichomes in Cannabis sativa. J. Nat. Prod. 1981, 44, 153–159. [Google Scholar] [CrossRef]
- Jensen, W.E. Botanical Histochemistry: Principles and Practice; W. H. Freeman: San Francisco, CA, USA, 1962. [Google Scholar]
- Gregory, M.; Baas, P. A survey of mucilage cells in vegetative organs of the dicotyledons. Isr. J. Bot. 1989, 38, 125–174. [Google Scholar]
- Fisher, D.B. Protein staining of ribboned epon sections for light microscopy. Histochemie 1968, 16, 92–96. [Google Scholar] [CrossRef] [PubMed]
- Vidal, B.C. Dichroism in collagen bundles stained with xylidine-Ponceau 2R. Ann. Histochim. 1970, 15, 289–296. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Organ | Echinodorus grandiflorus | Helanthium tenellum | Hydrocleys nymphoides | Limnocharis flava | Sagittaria rhombifolia | |
|---|---|---|---|---|---|---|
| Leaf | Petiole | + | + | + | + | + |
| Blade | + | + | + | + | + | |
| Flower | Floral scape | NA | NA | + | + | + |
| Floral organs | NA | NA | + | + | + |
| Reagent | Target Compound | Color | Echinodorus grandiflorus | Helanthium tenellum | Hydrocleys nymphoides | Limnocharis flava | Sagittaria rhombifolia |
|---|---|---|---|---|---|---|---|
| Schiff (PAS) | Neutral polysaccharides | pink | + | − | + | + | + |
| Lugol | Starch grains | blue | − | − | − | − | − |
| Sudan Black B | Total lipids | black | + | + | + | + | + |
| Sudan IV | Total lipids | light orange | + | + | + | + | + |
| Oil Red | Lipids | orange | + | + | + | + | + |
| Comassie Blue | Protein | blue | + | + | + | + | + |
| Wagner’s reagent | Alkaloids | blue-black to reddish | + | + | + | + | + |
| Toluidine Blue | Phenolic compounds | green | − | − | − | − | − |
| Ferric chloride | Phenolic compounds | brownish | − | − | − | − | − |
| Family/Species | Laticifers | Secretory Canal | Secretory Structure | References |
|---|---|---|---|---|
| Alismataceae Echinodorus grandiflorus, Helanthium tenellum, Hydrocleys nymphoides, Limnocharis flava, Sagittaria rhombifolia | Absent | Present | Secretory ducts anastomosed | This study |
| Alismataceae | Present | Absent | Not specified | [14] |
| Alismataceae (Limnocharitaceae) | Not specified | Not specified | Secretory ducts (“laticifers”) | [1,18,19] |
| Alismataceae | Not specified | Not specified | Secretory ducts or latex canal | [42] |
| Echinodorus macrophyllus | Not specified | Present | Secretory canal | [41] |
| E. glandulosus, E. lanceolatus, E. palaefolius, E. paniculatus, E. pubescens, E. subalatus subsp. subalatus. | Not specified | Not specified | Laticiferous ducts | [24] |
| Sagittaria guayanensis ssp. lappula | Not specified | Not specified | Laticifers or laticifer canal (articulated) | [11] |
| Sagittaria montevidensis | Not specified | Not specified | Schizogenous duct | [22] |
| Sagittaria acutifolia, Alisma plantago, Baldellia ranunculoides, Damasonium alisma, Echinodorus, Liminophyton obtusifolium, Ranalisma humile, Wisneria schweinfurthii | Not specified | Not specified | Secretory ducts | [19] |
| Sagittarialatifolia, Luronium natans and Wisneria | Not specified | Not specified | Tannin cells | [19] |
| Aponogetonaceae | Present | Absent | [14] | |
| Present | Absent | [15] | ||
| Present | Absent | Articulated laticifers and tannin cells | [64] | |
| Araceae | Present | Present | Not specified | [14,17] |
| Present | Present | Not specified | [16,50] | |
| Butomaceae | Absent | Absent | Not specified | [18] |
| Cymodoceaceae | Absent | Absent | Tannin cells | [18] |
| Hydrocharitaceae | Absent | Absent | Absence of schizogenous secretory ducts | [19,65] |
| Juncaginaceae Triglochin | Present | Absent | Not specified | [1,20] |
| Maundiaceae | Absent | Absent | Not specified | [1] |
| Posidoniaceae | Absent | Absent | Tannin cells | [62] |
| Ruppiaceae | Absent | Absent | Tannin cells | [1] |
| Scheuchzeriaceae | Absent | Absent | Tannin cells | [18] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Leme, F.M.; Bento, J.P.S.P.; Fabiano, V.S.; González, J.D.V.; Pott, V.J.; Arruda, R.d.C.d.O. New Aspects of Secretory Structures in Five Alismataceae Species: Laticifers or Ducts? Plants 2021, 10, 2694. https://doi.org/10.3390/plants10122694
Leme FM, Bento JPSP, Fabiano VS, González JDV, Pott VJ, Arruda RdCdO. New Aspects of Secretory Structures in Five Alismataceae Species: Laticifers or Ducts? Plants. 2021; 10(12):2694. https://doi.org/10.3390/plants10122694
Chicago/Turabian StyleLeme, Flávia Maria, João Pedro Silvério Pena Bento, Vitoria Silva Fabiano, Jean David Varilla González, Vali Joana Pott, and Rosani do Carmo de Oliveira Arruda. 2021. "New Aspects of Secretory Structures in Five Alismataceae Species: Laticifers or Ducts?" Plants 10, no. 12: 2694. https://doi.org/10.3390/plants10122694