Phytochemicals and Biological Activity of Desert Date (Balanites aegyptiaca (L.) Delile)
by
 , and
, and
Hosakatte Niranjana Murthy
1 ,
,
Guggalada Govardhana Yadav
1,
Yaser Hassan Dewir
2,3,* and
and
Abdullah Ibrahim
2 1
Department of Botany, Karnatak University, Dharwad 580003, India
2
Plant Production Department, P.O. Box 2460, College of Food and Agriculture Sciences, King Saud University, Riyadh 11451, Saudi Arabia
3
Faculty of Agriculture, Kafrelsheikh University, Kafr El-Sheikh 33516, Egypt
*
Author to whom correspondence should be addressed.
Plants 2021, 10(1), 32; https://doi.org/10.3390/plants10010032
Submission received: 28 November 2020
/
Revised: 19 December 2020
/
Accepted: 22 December 2020
/
Published: 25 December 2020
(This article belongs to the Special Issue Structural and Functional Analysis of Extracts in Plants)
Abstract
:Many underutilized tree species are good sources of food, fodder and possible therapeutic agents. Balanites aegyptiaca (L.) Delile belongs to the Zygophyllaceae family and is popularly known as “desert date”, reflecting its edible fruits. This tree grows naturally in Africa, the Middle East and the Indian subcontinent. Local inhabitants use fruits, leaves, roots, stem and root bark of the species for the treatment of various ailments. Several research studies demonstrate that extracts and phytochemicals isolated from desert date display antioxidant, anticancer, antidiabetic, anti-inflammatory, antimicrobial, hepatoprotective and molluscicidal activities. Mesocarp of fruits, seeds, leaves, stem and root bark are rich sources of saponins. These tissues are also rich in phenolic acids, flavonoids, coumarins, alkaloids and polysterols. Some constituents show antioxidant, anticancer and antidiabetic properties. The objective of this review is to summarize studies on diverse bioactive compounds and the beneficial properties of B. aegyptiaca.
1. Introduction
Balanites aegyptiaca (L.) Delile (Family: Zygophyllaceae) is an underutilized fruit-yielding tree (Figure 1A) native to Africa and distributed in tropical and subtropical regions of Africa, from Senegal in the west (16 °W) to Somali in the East (49 °E) and Jordan in the north (35 °N) to Zimbabwe in the south (19 °S). B. aegyptiaca is also distributed in India, Myanmar, Iran, Jordan, Oman, Palestine, Saudi Arabia, Syria and Yemen [1]. Young leaves (Figure 1B) and tender shoots are used as vegetables. Leaves and fruits are used as fodder for livestock [1]. Fiber obtained from tender bark and older dried bark is used for the preparation of medicines (Figure 1C). Unripe and ripe fruits (Figure 1D,E) are edible and popularly known as “desert date”. The fruits are processed into beverages and liquor. Timber is suitable for the construction of furniture, domestic items and musical instruments. The wood produces high-quality charcoal fuel and industrial activated charcoal. Gum or resin produced from stems are used as glue. Seeds contain about 49% edible oil (Figure 1F,G), which is also used in the production of biodiesel fuel [1].
B. aegyptiaca is used in African and Indian traditional medicine. Roots and bark are purgative and anthelmintic. A decoction of roots is used to treat malaria. The bark is used to deworm cattle, and the roots are boiled into a soup and used to treat edema and stomach pains. Roots are also used as an emetic [1]. The fruit is used to treat jaundice in Sudan [2]. Seed oil is used as a laxative and for the treatment of hemorrhoids, stomach aches, jaundice, yellow fever, syphilis and epilepsy [3]. Bark extracts are used to kill freshwater snails and copepods. A decoction of bark is also used as an abortifacient and antidote in West African traditional medicine [4].
2. Nutritional Composition of Fruits, Seeds and Leaves
Ripe fruits display a thin brownish epicarp (Figure 1E), dark brown and fleshy mesocarp (Figure 1E) and thick endocarp nut (Figure 1F). The edible parts of the pulp and kernel yield oil. The pulp is rich in carbohydrates (62.63%) and protein (9.19%; Table 1) [5]. Fruit pulp shows lesser amounts of fat (2.58%) and dietary fiber (2.93%). The overall energy value is 346.82 kcal/100 g. Fruits are also rich in minerals, including calcium, magnesium, phosphorus, potassium and sodium (Table 1) [6]. Iron, copper, manganese, lead, chromium, cobalt, cadmium and selenium are reported in lower concentrations (Table 1). Major fatty acids in fruit pulp are oleic (37.17%), linoleic (27.73%) and palmitic (22.02%; Table 2) [7]. The fruit pulp also exhibits amino acids (Table 3) [8] and vitamins (Table 3). Antinutritional factors are comparatively less (Table 4) [5].
Seeds are rich in fixed oil content (49.00%) with a significant content of proteins (32.40%) and carbohydrates (8.70%; Table 1) [9,10]. Seed oil is used for edible purposes; major fatty acids are linoleic (47.84%), oleic (22.80%), palmitic (16.68%) and stearic (11.67%) (Table 2) [11]. It has been demonstrated that biodiesel from seed oil meets all international biodiesel standards [11]. Seeds contain minerals, such as potassium, phosphorus and calcium in higher concentrations (Table 1) and amino acids (Table 3) [9]; seed cake is used for animal feed. However, seeds also contain oxalate (8.51 mg/g DW), antinutrient and possibly toxic constituents (Table 4).

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Nutritional and mineral composition of desert date pulp, seeds and leaves.
| Proximate (%) | |||
|---|---|---|---|
| Components | Pulp [5,6] | Seeds [9,10] | Leaves [12,13] |
| Moisture | 18.27 | 5.20 | 13.11 |
| Protein | 9.19 | 32.40 | 15.86 |
| Fat | 2.58 | 49.00 | 2.90 |
| Ash | 4.40 | 3.30 | 9.26 |
| Carbohydrate | 62.63 | 8.70 | 28.12 |
| Dietary fiber | 2.93 | 1.40 | 30.75 |
| Energy (kcal/100g) | 346.82 | 605.40 | 202.02 |
| Mineral composition (mg/g DW) | |||
| Calcium (Ca) | 1.41 | 1.51 | 0–2.65 |
| Iron (Fe) | 0.0494 | 0.0484 | NR * |
| Magnesium (Mg) | 0.73 | 0.887 | 0.23–0.77 |
| Phosphorus (P) | 0.48 | 3.60 | 1.51–5.32 |
| Potassium (K) | 22.20 | 6.36 | 1.76–4.81 |
| Sodium (Na) | 0.48 | 0.053 | NR |
| Zinc (Zn) | 0.0065 | 0.0286 | NR |
| Copper (Cu) | 0.0039 | 0.0118 | 0.05–0.65 |
| Manganese (Mn) | 0.0033 | 0.0192 | 0.02–0.08 |
| Lead (Pb) | 0.0030 | 0.0050 | NR |
| Chromium (Cr) | 0.0040 | 0.0060 | NR |
| Cobalt (Co) | 0.0107 | 0.0120 | NR |
| Cadmium (Cd) | 0.0347 | 0.0163 | NR |
| Selenium (Se) | 0.0005 | NR | NR |
* NR = not reported.
Young leaves and shoots are used as vegetables in African countries. Leaves and shoots are also popular livestock fodder [1]. Leaves are a good source of carbohydrates (28.12%) and proteins (15.86%) and contain ash (9.26%) and dietary fiber (30.75%; Table 1) [12]. Leaves also provide minerals (Table 1), fatty acids (Table 2), amino acids (Table 3) and vitamins (Table 3) [12,13]. Leaves contain antinutrients in meager concentrations (Table 4).
Table 2.
Fatty acid composition of desert date pulp, seeds and leaves.
| Fatty Acid | Pulp (% in oil) [7] | Seeds (% in oil) [11] | Leaves (µg/g DW) [13] |
|---|---|---|---|
| Saturated fatty acids (SFA) | |||
| Lauric (C12:0) | ND * | ND | 0.17–0.28 |
| Myristic (C14:0) | 0.1 | 0.05 | 0.043–0.074 |
| Pentadecylic (C15:0) | ND | 0.046 | ND |
| Palmitic (C16:0) | 22.02 | 16.683 | 0.29–1.98 |
| Margaric (17:0) | ND | 0.106 | ND |
| Stearic (C18:0) | ND | 11.67 | 0.008–0.049 |
| Nonadecylic(19:0) | ND | 0.032 | ND |
| Arachidic (C20:0) | ND | 0.34 | 0.274–0.439 |
| Behenic (C22:0) | ND | 0.059 | 0.026–0.038 |
| Tricosylic (C23:0) | ND | 0.012 | 0–0.003 |
| Lignoceric (C24:0) | ND | 0.042 | 0.028–0.298 |
| Hyenic (C25:0) | ND | ND | 0.078–0.121 |
| Monounsaturated fatty acids (MUSFA) | |||
| Pentadecenoic (C15:1) | ND | 0.003 | 0.08–0.31 |
| Palmitoleic (C16:1) | ND | 0.027 | ND |
| Oleic (18:1) | 37.17 | 22.807 | 0.03–0.061 |
| Nonadecenoic (19:1) | ND | 0.175 | ND |
| Gadoleic (20:1) | ND | 0.061 | ND |
| Tetracosenoic (C24:1) | ND | ND | 0.077–0.266 |
| Polyunsaturated fatty acids (PUSFA) | |||
| Hexadecadienoic (C16:2) | ND | ND | 0.406–1.835 |
| Linoleic (C18:2) | 27.73 | 47.847 | 0.025–0.642 |
| Eicosadienoic (C20:2) | ND | ND | 0.116–0.296 |
| Hexadecatrienoic (C16:3) | ND | ND | 0.761–2.142 |
| Octadecatrienoic (C18:3) | ND | ND | 0.20–0.525 |
| Total SFA (%) | 22.16 | 29.04 | 1.84–3.38 |
| Total MUSFA (%) | 37.17 | 23.073 | 0.179–0.637 |
| Total PUSFA (%) | 27.73 | 47.847 | 1.727–5.174 |
* ND = not detected.
Table 3.
Amino acid and vitamin composition of desert date pulp, seeds and leaves.
| Amino Acid | Pulp (mg/g DW) [5,8,14] | Seeds (g/100 g of Protein) [9] | Leaves (g/100 g of Protein) [12,13] |
|---|---|---|---|
| Alanine | 2.90 | 3.50 | 1.80 |
| Aspartic acid | 4.43 | 8.29 | 7.86 |
| Arginine | 3.00 | 2.70 | 4.20 |
| Cystine | 1.65 | 2.52 | 0.79 |
| Glutamic acid | 7.10 | 8.91 | 10.80 |
| Glycine | 2.52 | 4.10 | 9.65 |
| Histidine | 0.80 | 1.99 | 2.83 |
| Isoleucine | 1.87 | 3.47 | 3.50 |
| Leucine | 3.04 | 6.47 | 6.23 |
| Lysine | 1.64 | 5.00 | 4.51 |
| Methionine | 0.60 | 0.75 | 0.73 |
| Phenylalanine | 1.90 | 4.61 | 4.80 |
| Proline | 30.80 | 2.78 | 1.85 |
| Serine | 1.80 | 4.29 | 2.01 |
| Threonine | 2.17 | 4.25 | 2.88 |
| Tyrosine | 1.84 | 2.75 | 3.16 |
| Valine | 2.23 | 3.29 | 4.07 |
| Tryptophan | 0.70 | NR * | NR |
| Vitamin (mg/g DW) | |||
| Vitamin A | |||
| α-carotene | NR | NR | 0.33–0.54 |
| β-carotene | 0.6484 | NR | 0.25–0.81 |
| β-cryptoxanthin | NR | NR | 0.02–1.14 |
| Vitamin B | |||
| Thiamine | 0.0027 | NR | 0.24–0.51 |
| Riboflavin | 0.0007 | NR | NR |
| Niacin (B3) | 0.0174 | NR | NR |
| Vitamin B6 | 0.0021 | NR | NR |
| Vitamin C | |||
| Ascorbic acid | 1.05 | NR | 0.57–2.05 |
| Vitamin E | |||
| α-tocopherol | NR | NR | 0.08–0.57 |
| β-tocopherol | NR | NR | 0.01–0.04 |
| γ-tocopherol | NR | NR | 0.01–0.063 |
| δ-tocopherol | NR | NR | 0.13–0.96 |
| Vitamin K | |||
| Phylloquinone | NR | NR | 0.21–1.37 |
* NR = not reported.
3. Phytochemicals Isolated from Desert Date
B. aegyptiaca produces a variety of secondary metabolites, such as polyphenols (phenolic acids, flavonoids and coumarins), alkaloids, steroids, saponins (spirostanol saponins, furostanol saponins and open-chain steroidal saponins) and pregnane glycosides, isolated from plant tissues, such as fruit, seeds, leaves, stem bark, roots and galls (Table 5).
3.1. Polyphenols
Polyphenols exhibit phenolic structural features with one or more aromatic rings, each with one or more hydroxyl groups [15]. Polyphenols are grouped into phenolic acids, flavonoids, stilbenes, lignans and tannins. These compounds are important as natural therapeutic agents involved in the prevention of degenerative diseases, particularly cancers, cardiovascular diseases and neurodegenerative diseases [16].
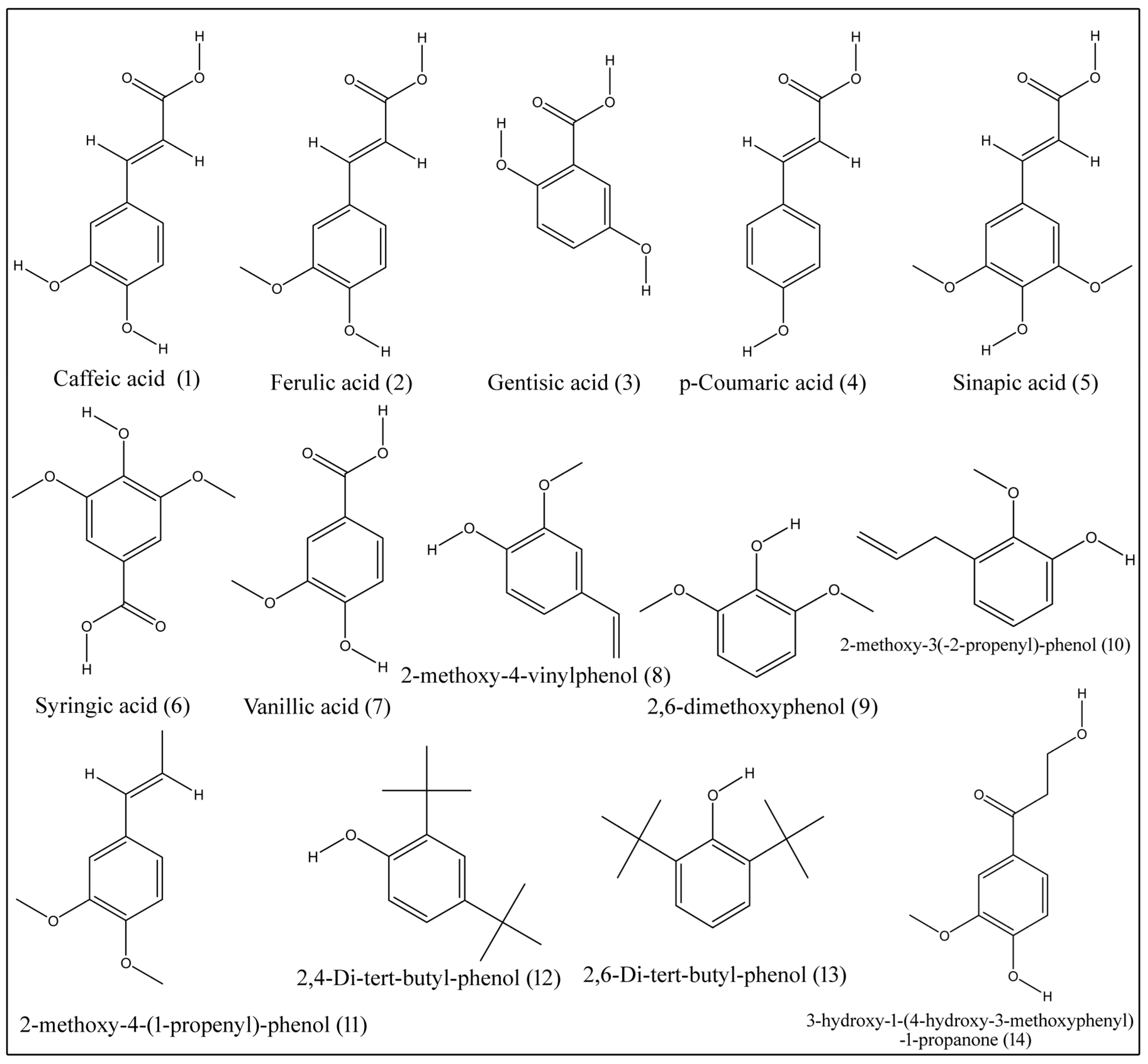
Phenolic acids are nonflavonoid polyphenolic compounds of benzoic acid and cinnamic acid. Major phenolic acids, which are isolated from tissues of B. aegyptiaca, include caffeic acid (1), ferulic acid (2), gentisic acid (3), p-coumaric acid (4), sinapic acid (5), syringic acid (6), vanillic acid (7), 2-methoxy-4-vinylphenol (8), 2,6-dimethoxyphenol (9), 2-methoxy-3(-2-propenyl)-phenol (10), 2-methoxy-4-(1-propenyl)-phenol (11), 2,4-di-tert-butyl-phenol (12), 2,6-di-tert-butyl-phenol (13) and 3-hydroxy-1-(4-hydroxy-3-methoxyphenyl)-1-propanone (14) (Table 5; Figure 2) [2,17,18,19].
3.2. Flavonoids
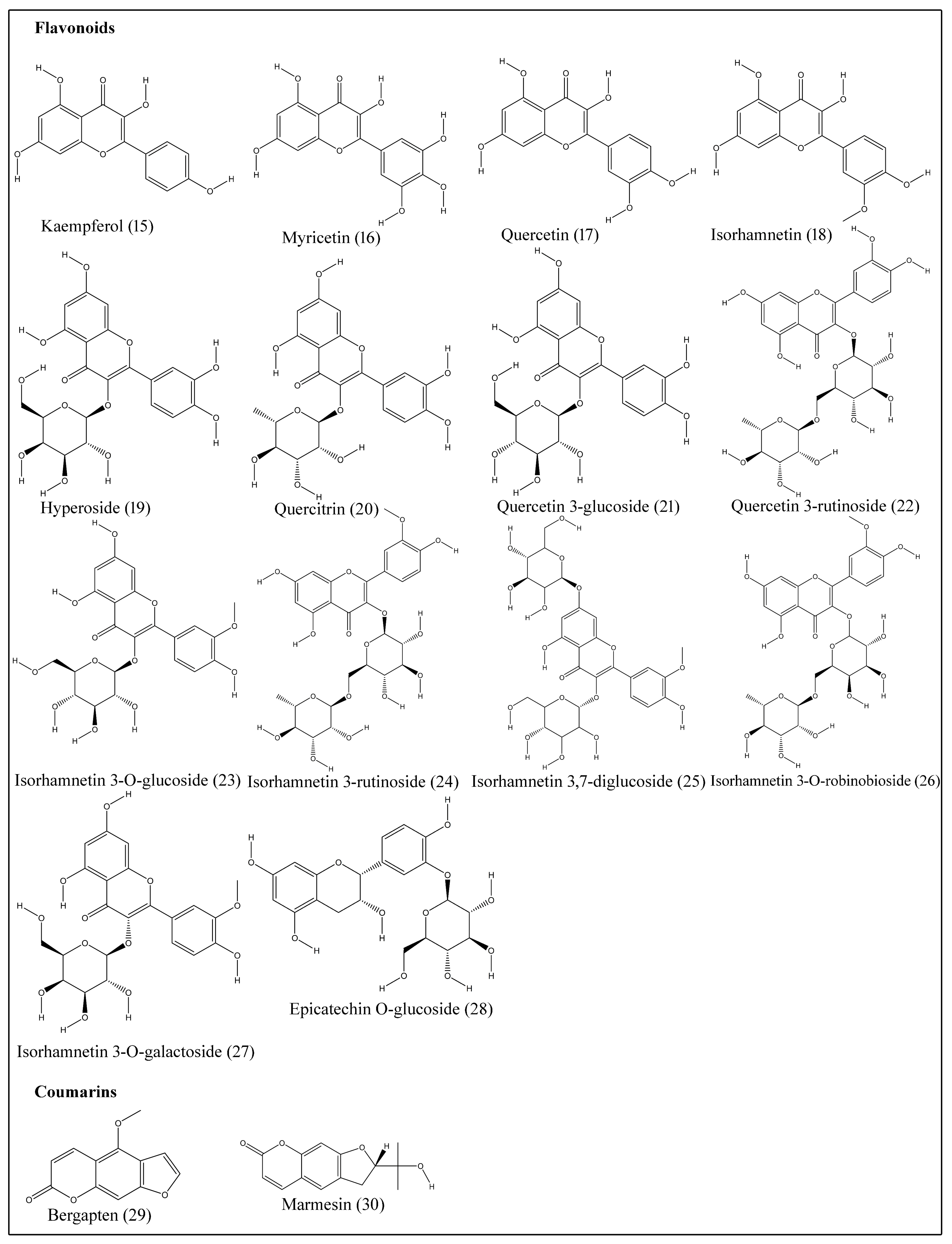
Flavonoids exhibit a diphenyl propane–flavone skeleton with a three-carbon bridge between phenyl groups and commonly cyclized with oxygen. Epicatechin O-glucoside (28), hyperoside (19), isorhamnetin (18), isorhamnetin-3-O-glucoside (23), isorhamnetin 3,7-diglucoside (25), isorhamnetin 3-O-galactoside (27), isorhamnetin 3-O-robinobioside (26), isorhamnetin 3-rutinoside (24), kaempferol (15), myricetin (16), quercetin (17), quercetin 3-glucoside (21), quercetin 3-rutinoside (22) and quercitrin (20) are isolated from different tissues of B. aegyptiaca (Table 5; Figure 3) [19,20,21,22].
Table 5.
Phytochemicals isolated from various parts of desert date.
| No. | Compound Name | Plant Parts | Reference |
|---|---|---|---|
| PHENOLICS | |||
| 1 | Caffeic acid | Gall, leaf | [19] |
| 2 | Ferulic acid | Gall, leaf | [19] |
| 3 | Gentisic acid | Gall, leaf | [19] |
| 4 | p-Coumaric acid | Gall, leaf | [19] |
| 5 | Sinapic acid | Gall, leaf | [19] |
| 6 | Syringic acid | Stem bark | [2] |
| 7 | Vanillic acid | Stem bark | [2] |
| 8 | 2-methoxy-4-vinylphenol | Fruit, leaf | [18] |
| 9 | 2,6-dimethoxyphenol | Leaf | [18] |
| 10 | 2-methoxy-3(-2-propenyl)-phenol | Leaf | [18] |
| 11 | 2-methoxy-4-(1-propenyl)-phenol (Isoeugenol) | Leaf | [18] |
| 12 | 2,4-di-tert-butyl-phenol | Seed | [17] |
| 13 | 2,6-di-tert-butyl-phenol | Seed | [17] |
| 14 | 3-hydroxy-1-(4-hydroxy-3-methoxyphenyl)-1-propanone | Stem bark | [2] |
| FLAVONOIDS | |||
| 15 | Kaempferol | Leaf | [19] |
| 16 | Myricetin | Leaf | [19] |
| 17 | Quercetin | Fruit, leaf, seed | [19,20,22] |
| 18 | Isorhamnetin | Fruit, seed | [20,22] |
| 19 | Hyperoside | Gall, leaf | [19] |
| 20 | Quercitrin | Leaf | [19] |
| 21 | Quercetin 3-glucoside (isoquercetin) | Leaf, seed | [19,21,22] |
| 22 | Quercetin 3-rutinoside (Rutin) | Fruit, gall, leaf, seed | [19,20,21,22] |
| 23 | Isorhamnetin-3-O-glucoside | Fruit, leaf, seed | [20,21,22] |
| 24 | Isorhamnetin 3-rutinoside | Fruit, leaf, seed | [20,21,22] |
| 25 | Isorhamnetin 3,7-diglucoside | Leaf, seed | [21,22] |
| 26 | Isorhamnetin 3-O-robinobioside | Seed | [22] |
| 27 | Isorhamnetin 3-O-galactoside | Seed | [22] |
| 28 | Epicatechin O-glucoside | Fruit | [20] |
| COUMARINS | |||
| 29 | Bergapten | Stem bark | [23] |
| 30 | Marmesin | Stem bark | [23] |
| ALKALOIDS | |||
| 31 | N-trans-Feruloyltyramine | Stem bark | [2] |
| 32 | N-cis-feruloyltyramine | Stem bark | [2] |
| 33 | Trigonelline | Fruit | [20] |
| STEROIDS | |||
| 34 | Diosgenin | Fruit | [24] |
| 35 | Yamogenin | Fruit, root, stem bark | [25] |
| 36 | 6-Methyldiosgenin | Fruit | [24] |
| 37 | Rotenone | Root | [26] |
| 38 | β-Sitosterol | Seed (oil) | [27] |
| 39 | Cholesterol | Seed (oil) | [27] |
| 40 | Campesterol | Seed (oil) | [27] |
| 41 | Stigmasterol | Seed (oil) | [27] |
| PREGNANE GLYCOSIDES | |||
| 42 | Pregn-5-ene-3β,16β,20(R)-triol 3-O-(2,6-di-O-α-L-rhamnopyranosyl)-β-D-glucopyranoside | Fruit | [28] |
| 43 | Pregn-5-ene-3β,16β,20(R)-triol 3-O-β-D-glucopyranoside | Fruit | [28] |
| SAPONINS | |||
| SPIROSTANOL SAPONINS | |||
| 44 | Balanitin 1 | Root, stem bark | [29] |
| 45 | Balanitin 2 | Root, stem bark | [29] |
| 46 | Balanitin 3 | Root, stem bark | [29] |
| 47 | Balanitin 4 | Seed | [30] |
| 48 | Balanitin 5 | Seed | [30] |
| 49 | Balanitin 6 | Seed | [30,31] |
| 50 | Balanitin 7 | Fruit, root, seed | [30,31,32] |
| 51 | Deltonin | Seed | [33] |
| 52 | (3β,20S,22R,25R)-spirost-5-en-3-yl β-D-xylopyranosyl-(1→3)-β-D-glucopyranosyl-(1→4)[α-L-rhamnopyranosyl-(1→2)]-β-D-glucopyranoside | Root | [34] |
| 53 | (3β,20S,22R,25S)-spirost-5-en-3-yl β-D-xylopyranosyl-(1→3)-β-D-glucopyranosyl-(1→4)[α-L-rhamnopyranosyl-(1→2)]-β-D-glucopyranoside | Root | [34] |
| FUROSTANOL SAPONINS | |||
| 54 | Balanitesin | Fruit | [35] |
| 55 | Balanitoside | Fruit | [24,36] |
| 56 | 22R and 22S epimers of 26-(O-β-D-glucopyranosyl)-3-β-[4-O-(β-D-glucopyranosyl)-2-O-(α-L-rhamnopyranosyl)-β-D-glucopyranosyloxy]-22,26-dihydroxyfurost-5-ene | Fruit | [37] |
| 57 | Xylopyranosyl derivative of 26-(O-β-D-glucopyranosyl)-3-β-[4-O-(β-D-glucopyranosyl)-2-O-(α-L-rhamnopyranosyl)-β-D-glucopyranosyloxy]-22,26-dihydroxyfurost-5-ene | Fruit | [37] |
| 58 | (3β,20S,22R,25R)-26-(β-D-glucopyranosyloxy)-22-methoxyfurost-5-en-3-yl β-D-xylopyranosyl-(1→3)-β-D-glucopyranosyl-(1→4) [α-L-rhamnopyranosyl-(1→2)]-β-D-glucopyranoside | Root | [34] |
| 59 | (3β,20S,22R,25S)-26-(β-D-glucopyranosyloxy)-22-methoxyfurost-5-en-3-yl β-D-xylopyranosyl-(1→3)-β-D-glucopyranosyl-(1→4) [α-L-rhamnopyranosyl-(1→2)]-β-D-glucopyranoside | Root | [34] |
| 60 | 26-O-β-D-glucopyranosyl-(25R)-furost-5-ene-3β,22,26-triol 3-O-(2,4-di-O-α-L-rhamnopyranosyl)-β-D-glucopyranoside | Fruit | [38] |
| 61 | 22-methyl ether of 26-O-β-D-glucopyranosyl-(25R)-furost-5-ene-3β,22,26-triol 3-O-(2,4-di-O-α-L-rhamnopyranosyl)-β-D-glucopyranoside | Fruit | [38] |
| 62 | 26-O-β-D-glucopyranosyl-(25R)-furost-5-ene-3β,22,26-triol 3-O-[α-L-rhamnopyranosyl-(1→2)]-[β-D-xylopyranosyl (1→3)]-[α-L-rhamnopyranosyl-(1→4)]-β-D-glucopyranoside | Fruit | [38] |
| 63 | 22-methyl ether of 26-O-β-D-glucopyranosyl-(25R)-furost-5-ene-3β,22,26-triol 3-O-[α-L-rhamnopyranosyl-(1→2)]-[β-D-xylopyranosyl (1→3)]-[α-L-rhamnopyranosyl-(1→4)]-β-D-glucopyranoside | Fruit | [38] |
| 64 | Balanin B2 | Stem bark | [39] |
| 65 | 26-(O-β-D-glucopyranosyl)-22-O-methylfurost- 5-ene-3β,26-diol-3-O-β-D-glucopyranosyl-(1→4)- [α-L-rhamnopyranosyl-(1→2)]-β-D-glucopyranoside | Fruit | [40] |
| 66 | 25R and 25S epimers of 26-O-β-D-glucopyranosyl-furost-5-ene-3,22,26- triol 3-O-[α-L-rhamnopyranosyl-(1→3)- β-D-glucopyranosyl-(1→2)]- α-L-rhamnopyranosyl-(1→4)-β- D-glucopyranoside | Fruit | [41] |
| 67 | 26-O-β-D-glucopyranosyl-(25R)-furost-5-ene-3,22,26-triol 3-O-[ β-D-glucopyranosyl-( 1→2)]- α-L-rhamnopyranosyl-(1→4)-β-D-glucopyranoside | Fruit | [41] |
| 68 | 26-O-β-D-glucopyranosyl-(25R)-furost-5,20-diene-3,26-diol 3-O-[α-L-rhamnopyranosyl-(1→3)- β-D-glucopyranosyl-(1→2)]- α-L-rhamnopyranosyl-(1→4)-β-D-glucopyranoside | Fruit | [41] |
| 69 | 25R and 25S epimers of 26-O-β-D-glucopyranosyl-furost-5,20-diene-3,26-diol 3-O-[ β-D-glucopyranosyl-(1→2)]- α-L-rhamnopyranosyl-(1→4)-β-D-glucopyranoside | Fruit | [41] |
| OPEN-CHAIN STEROIDAL SAPONINS | |||
| 70 | (3β,12α,14β,16β)-12-hydroxycholest-5-ene-3,16-diyl bis (β-D-glucopyranoside) | Root | [34] |
| 71 | Balanin B1 | Stem bark | [39] |
| 72 | β-Sitosterol glucoside | Stem bark | [42] |
| 73 | Stigmasterol-3-O-β-D-glucopyranoside | Stem bark | [40] |
3.3. Coumarins
Coumarins are phenolic compounds displaying fused benzene and α-pyrone rings and are known for anti-inflammatory, anticoagulant, antimicrobial, anticancer, antioxidant and neuroprotective properties [43]. Bergapten (29) and marmesin (30) are coumarins extracted from stem bark (Table 5; Figure 3) [23].
3.4. Alkaloids
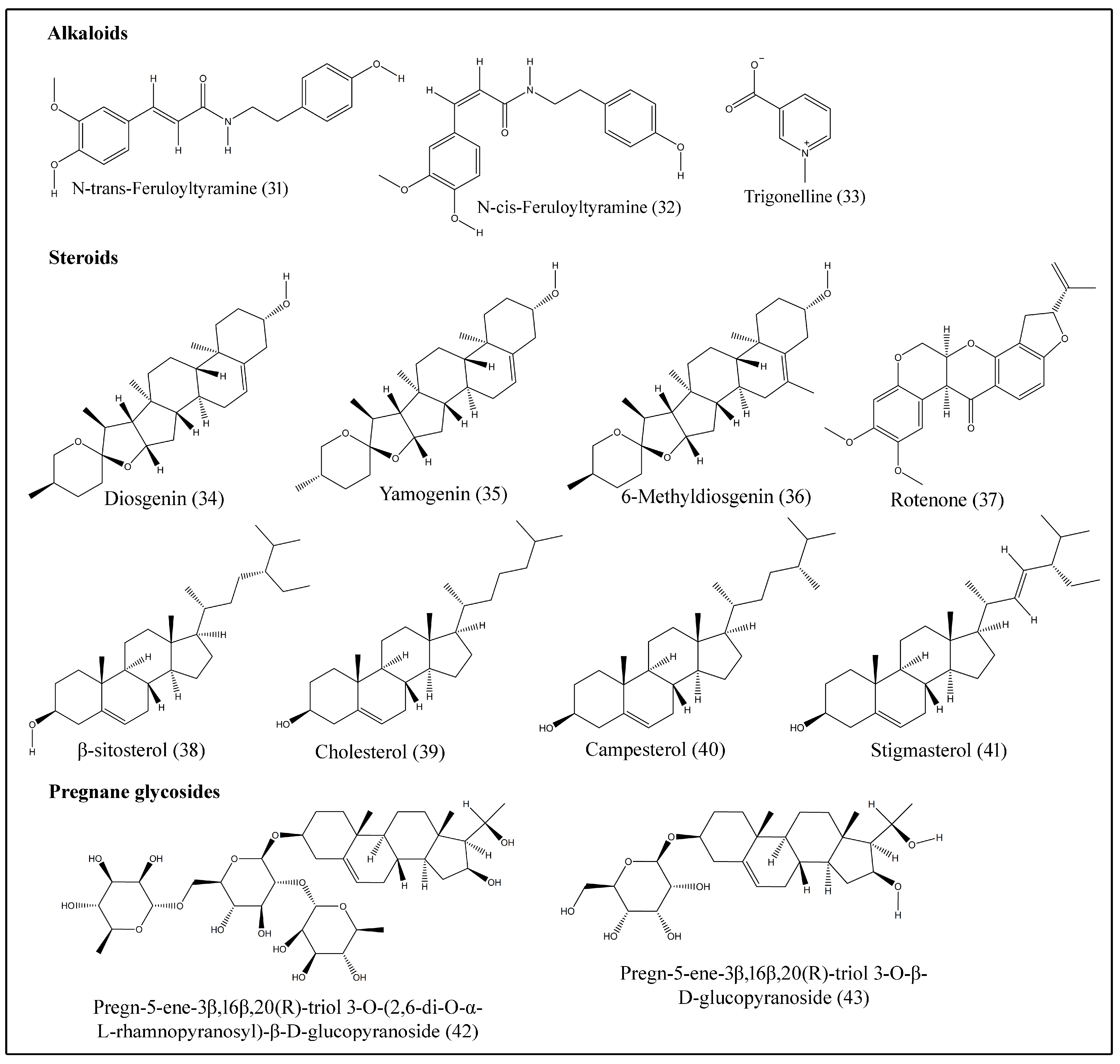
Alkaloids are compounds that contain basic nitrogen atoms [44] and show varied biological activities. They are especially useful for cancer treatment. N-cis-feruloyltyramine (32), N-trans-feruloyltyramine (31) and trigonelline (33) are some of the alkaloids isolated from stem bark and fruit (Table 5; Figure 4) [2,20].
3.5. Phytosterols
Phytosterols are bioactive compounds found naturally in food with chemical structures similar to cholesterol. Various clinical studies consistently show that intake of phytosterols, such as beta-sitosterol, campesterol and stigmasterol, is associated with a significant reduction in levels of low-density lipoprotein in humans. B. aegyptiaca produces several steroids, such as campesterol (40), cholesterol (39), diosgenin (34), 6-methyldiosgenin (36), rotenone (37), β-sitosterol (38), stigmasterol (41) and yamogenin (35) (Table 5; Figure 4) [24,25,26,27].
Pregnane glycosides are naturally occurring sugar conjugates of C21 steroidal compounds, isolated from various plants and many show anticarcinogenic properties [45]. Pregn-5-ene-3β,16β,20(R)-triol 3-O-(2,6-di-O-α-L-rhamnopyranosyl)-β-D-glucopyranoside (42) and pregn-5-ene-3β,16β,20(R)-triol 3-O-β-D-glucopyranoside (43) were extracted from the fruits of desert date (Table 5; Figure 4) [28].
3.6. Saponins
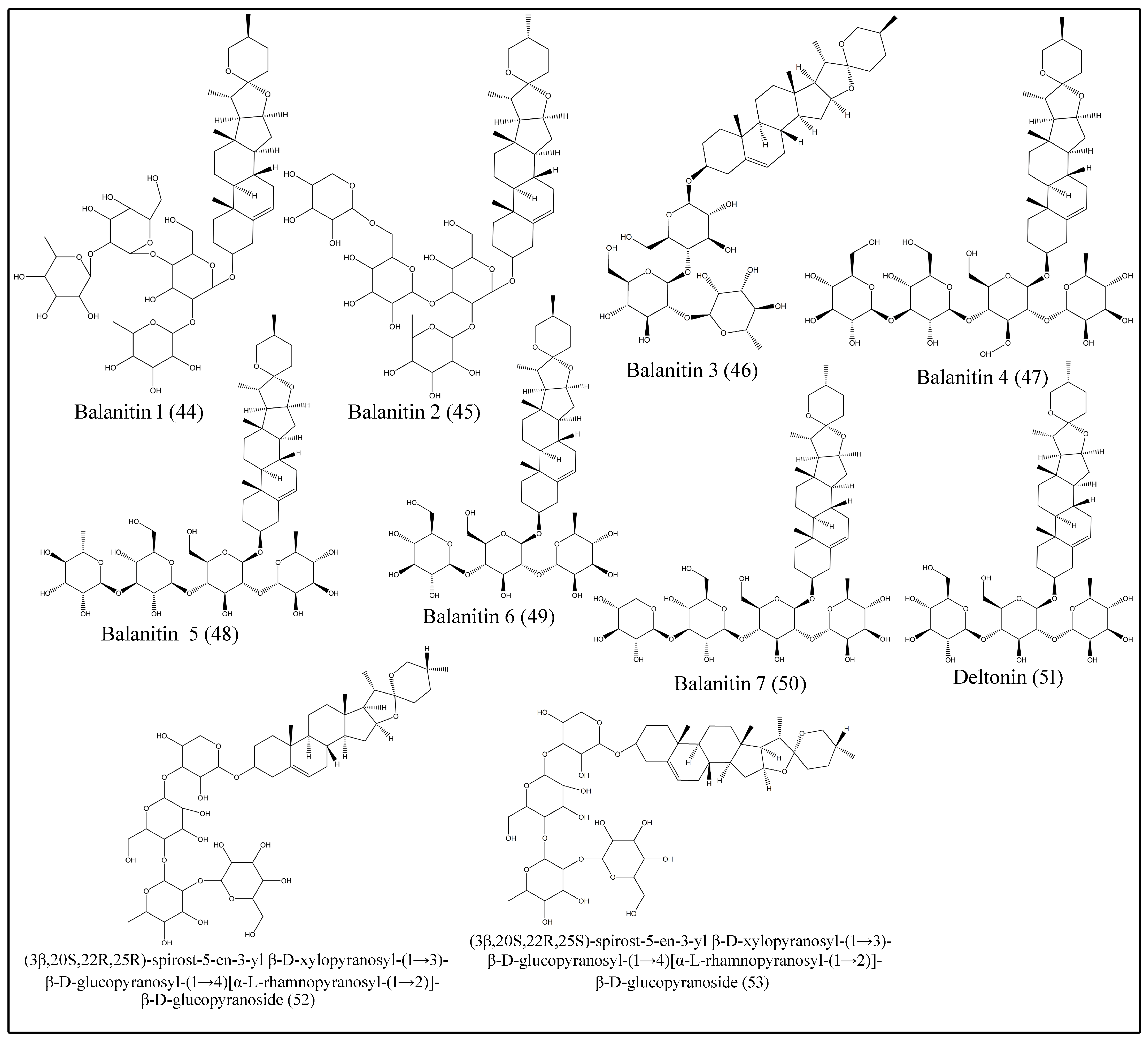
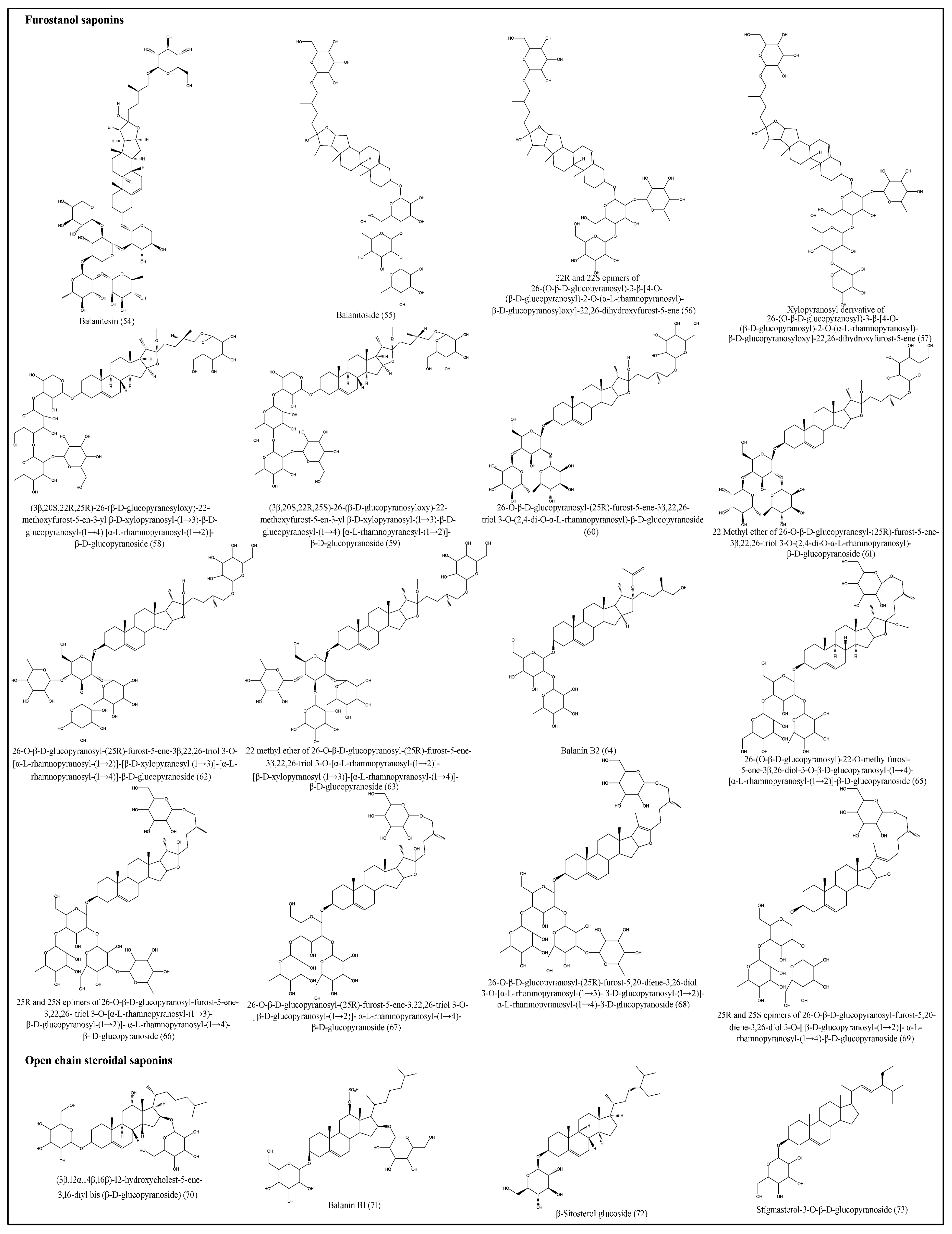
Saponins are bioorganic compounds that exhibit triterpenoid or steroidal skeletons that are glycosylated by varying numbers of sugar moieties attached at different positions. Steroidal saponins are further classified into spirostanol, furostanol and open-chain steroidal saponins [46]. Saponins exhibit a wide range of biological properties, including hemolytic factor sand anti-inflammatory, antimicrobial, insecticidal, anticancer and molluscicidal activities [47]. Various spirostanol, furostanol and open-chain steroidal saponins, which are isolated from fruits, seeds, roots and stem bark are presented in Table 5 and Figure 5 and Figure 6 [24,29,30,31,32,33,34,35,36,37,38,39,40,41,42].
4. Biological Activity
Extracts and compounds from extractions of B. aegyptiaca todate exhibit a wide range of biological activity (Table 6).
4.1. Antioxidant Properties
Various kinds of physical and physiological stresses lead to the overproduction of oxidants in the human body, which can cause oxidative damage of DNA, proteins and lipids. Furthermore, this damage is responsible for several disorders in the human body such as cardiovascular diseases, cancer and aging. It was reported that minor fruits and nuts possess abundant antioxidant phytochemicals, and the consumption of minor fruits and nuts is beneficial to the human body [48]. The antioxidant effects of methanol extracts of stem bark on 1,1-diphenyl-2-picrylhydrazyl (DPPH) and 2,2’-azino-bis(3-ethylbenzothiazoline-6-sulfonic acid (ABTS) scavenging is demonstrated and accounted for in the total soluble phenolic and flavonoid contents [49]. Furthermore, Hassan et al.’s [49] results show that methanol extracts display the highest phenolic content (35.17 mg gallic acid equivalents/g) and considerable flavonoid content (112.83 mg quercetin equivalents/g). Methanol extract showed the highest free radical scavenging activity at IC50 = 40 µg/mL and IC50 = 125.85 µg/mL in DPPH and ABTS assays, respectively. The antioxidant properties of aqueous fruit extracts were assessed in streptozotocin-induced diabetic rats [50]. Oral administration produced a significant (p < 0.01) increase in mean plasma total antioxidant levels and a significant (p < 0.01) decrease in malondialdehyde levels. The antioxidant properties of leaf and root extracts were also demonstrated [51]. Balanitin 1 and balanitin 2 (saponins) were isolated from bark extracts and demonstrated antioxidant properties in vitro, using a method based on the Briggs–Rauscher oscillating reaction [39]. Polyphenols such as quercetin and kaempferol are the major components responsible for antioxidant activities [52]. In addition, phytosterols including ß-sitoterol, stigmasterol and campesterol have been reported to exert antioxidant activity [53]. Polyphenols, phytosterols and saponins together might be responsible for the antioxidant activity of desert date.
4.2. Antimicrobial Properties
Plants synthesize several antimicrobial compounds, including phenolics such as simple phenols, phenolic acids, quinones, flavonoids, flavones, flavonols, tannins, coumarins, terpenoids, essential oils and alkaloids [54]. The mechanism of action of these compounds ranges from membrane disruption, substrate deprivation, intercalation into the cell wall/or DNA and enzyme inhibition. Desert date is rich in all these phytochemicals and demonstrates potent antimicrobial activity. The bark of B. aegyptiaca is widely used in African folk medicine for the treatment of wounds and skin diseases. The effects of aqueous ethanolic extracts of bark on bacteria isolated from wounds have been reported [55]. These extracts inhibited the growth of Pseudomonas aeruginosa and Staphylococcus aureus in vitro. The in vitro antifungal activity of saponin-rich extracts of fruit mesocarp was explored against phytopathogenic fungi [56]. These extracts were moderately active (34.7%) against Alternaria solani and highly active (89.01%) against Pythium ultimum, and activity was significantly higher compared to the fungicide, metalaxyl (15 μg/mL). The antifungal activity of ethanolic and methanolic extracts of root bark and fruit have been demonstrated against Aspergillus niger, Candida albicans, Penicillium crustosum and Saccharomyces cerevisiae [57].
4.3. Hepatoprotective Properties
A methanolic extract of leaves was evaluated for hepatoprotective activity against carbon tetrachloride (CCl4)-induced hepatic damage in rats [58]. Administration of the extract (200 and 400 mg/kg per os) markedly reduced the CCl4-induced elevation of serum marker enzymes, such as glutamate pyruvate transaminase, glutamate oxaloacetate transaminase, alkaline phosphatase and bilirubin. Similarly, fruit mesocarp and stem bark aqueous extracts ameliorated CCl4-induced hepatotoxicity in rats, as measured by liver enzyme activity, blood parameters and histopathology [59]. Ethanolic extracts of bark protected hepatocytes against paracetamol and CCl4-induced hepatotoxicity in rats, analogous to silymarin [60]. Bioactive compounds, primarily obtained from dietary sources, contain a wide range of free radical scavenging constituents, including polyphenols, alkaloids and phytosterols, which are responsible for hepatoprotective effects [61]. Desert date is rich in polyphenols, phytosterols and saponins. It has depicted very good antioxidant potential and thus increased the cellular antioxidant defense system, which may be responsible for the hepatoprotective effects of desert date.
4.4. Anticancer Properties
Cancer is a major health problem. Radiotherapy, chemotherapy and surgical removal are the current treatment methods. However, these methods have varied disadvantages such as drug resistance and toxic effects on nontargeted tissues. Therefore, researchers are searching for naturally available plant-based bioactive compounds for cancer therapy [62]. Among the plant-based bioactive compounds, saponins and phytosterols have significant importance in reducing the risk of cancer [63,64]. Various steroidal saponins isolated from various tissues of B. aegyptiaca are reported to display anticancer activities. For example, a mixture of balanitin-6 and balanitin-7 (28:72) isolated from kernels show growth inhibition in human cancer cell lines in vitro [31]. Balanitin-6/balanitin-7 exhibited higher antiproliferative activity than well-known natural cancer therapeutic agents, such as etoposide and oxaliplatin. Balanitin-6/balanitin-7 displayed its highest activity against A549 nonsmall cell lung cancer (IC50, 0.3 µM) and U373 glioblastoma (IC50, 0.5 µM) cell lines. Balanitoside extracted from the fruit also showed anticancer activity against Ehrlich ascites carcinoma (EAC)-bearing Swiss albino mice [36]. Mice injected intraperitoneally with balanitoside (10 mg/kg body weight) displayed decreases in liver and serum enzyme levels. Issa et al. [65] studied an aqueous extract of pulp on the development and growth of EAC and metastasis to the liver and spleen. Treatment with the extract (400 mg/kg) inhibited tumor growth and proliferation in ascetic fluid, inducing a significant decrease in tumor volume, total cell volume and viable cell count and prolonged mouse survival. The authors also recorded significant decreases in levels of lipid peroxidation and increased superoxide dismutase and catalase activity and P53 (a tumor suppressor protein) expression. The saponin, balanitin-7 isolated from seed kernels, showed antiproliferative activity [32]. These agents showed potent antiproliferative activity against MCF-7 human breast cancer cells and HT-29 human colon cancer cells, with IC50 values of 2.4 and 3.3 µM, respectively.
4.5. Anti-Inflammatory Properties
Inflammation is a pattern of response to injury, which involves the accumulation of cells, exudates in irritated tissue, which allows protection from further damage. A variety of in vitro and in vivo experiments has shown that certain flavonoids and saponins possess anti-inflammatory activity [66]. The mechanism by which flavonoids and saponins exert their anti-inflammatory effects involves the inhibition of cyclooxygenase and lipoxygenase activities [67]. Desert date exhibited potent anti-inflammatory activity; for example, Speroni et al. [39] studied the in vivo anti-inflammatory activity of methanol and butanol extracts and two saponins, viz. balanin-B1 and balanin-B2, isolated from B. aegyptiaca bark in rats with edema induced by carrageenin. Both extracts exhibited a significant reduction of rat paw edema. The inhibition produced by methanol extract, butanol extract, balanin-B1 and balanin-B2 were 32%, 68%, 62% and 59%, respectively. Likewise, the influence of seed oil on liver and kidney fractions in rat serum was evaluated [68]. Seed oil (100 mg/kg) in the rat diet decreased nitrogen oxide and lipid peroxidation. Further, mRNA and protein expression of tumor necrosis factor-α and interleukin-6 were downregulated, leading to a reduction of cyclooxygenase-2, reflecting anti-inflammatory activity.
4.6. Antidiabetic Activity
Diabetes is a chronic disease that occurs either when the pancreas does not produce enough insulin or when the body cannot effectively use the insulin it produces. Several medicinal plants have demonstrated hypoglycemic and hyperglycemic activities; these activities seem to be mediated through increased insulin secretion via stimulation of pancreatic cells, interfering with dietary glucose absorption or through insulin-sensitizing action [69]. Kamel et al. [38] demonstrated the antidiabetic effect of an aqueous extract of fruit in streptozotocin (STZ)-induced diabetic mice after oral administration. They also identified steroidal saponins, 26-O-ß-D-glucopyranosyl-(25R)-furost-5-ene-3ß,22,26-triol-3-O-[α-L-rhamnophyranosyl-(1→1)]-[ß-D-xylopyranosyl-(1→3)]-[α-L-rhamnopyronosyl-(1→4)]-ß-D-glucopyranoside and its 22-methyl ether in the extract and recognized two additional saponins, 26-O-ß-D-glycopyranosyl-(25R)-furost-5-ene-3ß,22,26,-triol-3-O-[2,4-di-O-α-L-rhamnopyranosyl)-ß-D-glucopyranoside and its methyl ether. A combination of saponins exhibited greater antidiabetic activity than individual saponins. Gad et al. [70] administered fruit extracts (1.5 g/kg body weight) to STZ-induced diabetic rats and studied the glycogen content of liver and kidney and on some key enzymes of liver involved in carbohydrate metabolism. STZ (50 mg/kg body weight) caused a five-fold increase in blood glucose level, an 80% reduction in serum insulin level, a 58% decrease in liver glycogen and a seven-fold increase in kidney glycogen content. A marked increment in the activity of glucose-6-phosphatase activity and decreased activity of glucose-6-phosphate dehydrogenase and phosphofructokinase were recorded. Treatment of rats with fruit extract reduced blood glucose levels by 24% and significantly decreased liver glucose-6-phosphatase activity. The authors also demonstrated that the extract inhibited α-amylase activity in vitro. The major component in the extract was diosgenin, based on high-performance thin-layer chromatography. Additionally, Al-Malki et al. [71] showed that ethyl acetate extract containing β-sitosterol modulated oxidative stress induced by streptozotocin.
Table 6.
Biological activities of compounds isolated from various parts of desert date.
| Compound | Part | Activity | Model/Method | Reference |
|---|---|---|---|---|
| Balanitin 1 | Root and stem bark | Molluscicide | Biomphalaria glabrata | [29] |
| Balanitin 2 | Root and stem bark | Molluscicide | Biomphalaria glabrata | [29] |
| Balanitin 3 | Root and stem bark | Molluscicide | Biomphalaria glabrata | [29] |
| Balanitin 4 | Seed | Anticancer | P-388 Lymphocytic leukemia cell line | [30] |
| Balanitin 5 | Seed | Anticancer | P-388 Lymphocytic leukemia cell line | [30] |
| Balanitin 6 | Seed | Anticancer | Different cancer cell lines including the P-388 Lymphocytic leukemia cell line and female mice injected with L1210 syngeneic murine leukemia cells | [30,31] |
| Balanitin 7 | Fruit, root, seed | Anticancer | 1.Different cancer cell lines and female mice injected with L1210 syngeneic murine leukemia cells 2. P-388 Lymphocytic leukemia cell line 3.Human breast cancer cells (MCF-7) and human colon cancer cells (HT-29) | [30,31,32] |
| Nematocidal | Caenorhabditis elegans | [72] | ||
| Deltonin | Seed | Molluscicidal | Biomphalaria glabrata | [33] |
| Balanitoside | Fruit | Anticancer | Ehrlich ascites carcinoma bearing Swiss albino mice | [36] |
| Antidiabetic | Streptozotocin-induced diabetes in Wistar rats | [73] | ||
| Balanin B2 | Stem bark | Anti-inflammatory | Carrageenin-induced paw edema in male Sprague Dawley rats | [39] |
| 26-(O-β-D-glucopyranosyl)-22-O-methylfurost- 5-ene-3β,26-diol-3-O-β-D-glucopyranosyl-(1→4)-[α-L-rhamnopyranosyl-(1→2)]-β-D-glucopyranoside | Fruit | Antidiabetic | α-Glucosidase and aldose reductase inhibitory activities (in vitro) and streptozotocin-induced diabetes in male albino Wistar rats (in vivo) | [40] |
| 25R and 25S epimers of 26-O-β-D-glucopyranosyl-furost-5-ene-3,22,26- triol 3-O-[α-L-rhamnopyranosyl-( 1→3)- β-D-glucopyranosyl-(1→2)]- α-L-rhamnopyranosyl-(1→4)-β- D-glucopyranoside | Fruit | Aldose reductase inhibitor | Aldose reductase inhibition activity on rat liver homogenate | [41] |
| 26-O-β-D-glucopyranosyl-(25R)-furost-5-ene-3,22,26-triol 3-O-[ β-D-glucopyranosyl-( 1→2)]- α-L-rhamnopyranosyl-(1→4)-β-D-glucopyranoside | Fruit | Aldose reductase inhibitor | Aldose reductase inhibition activity on rat liver homogenate | [41] |
| 26-O-β-D-glucopyranosyl-(25R)-furost-5,20-diene-3,26-diol 3-O-[α-L-rhamnopyranosyl-(1→3)- β-D-glucopyranosyl-(1→2)]- α-L-rhamnopyranosyl-(1→4)-β-D-glucopyranoside | Fruit | Aldose reductase inhibitor | Aldose reductase inhibition activity on rat liver homogenate | [41] |
| 25R and 25S epimers of 26-O-β-D-glucopyranosyl-furost-5,20-diene-3,26-diol 3-O-[ β-D-glucopyranosyl-(1→2)]- α-L-rhamnopyranosyl-(1→4)-β-D-glucopyranoside | Fruit | Aldose reductase inhibitor | Aldose reductase inhibition activity on rat liver homogenate | [41] |
| Balanin B1 | Stem bark | Anti-inflammatory | Carrageenin-induced paw edema in male Sprague Dawley rats | [39] |
| Antioxidant | ROS scavenging activity by Briggs-Rauscher oscillating reaction | [39] |
Hassanin et al. [74] tested a crude ethanolic fruit extract and its butanolic and dichloromethane fractions on stress-activated protein kinase/c-Jun N-terminal kinase (SAPK-JNK) signaling in experimental diabetic rats. Six groups of male Wistar rats were used: normal control, diabetic, diabetic rats treated with crude, butanol or dichloromethane factions (50 mg/kg body weight), and diabetic rats were treated with gliclazide as a reference drug. Treatments continued for one month. Extract treatments produced a reduction in plasma glucose, hemoglobin A1c, lactic acid, lipid profile and malondialdehyde levels, which induced an increase in insulin and reduced glutathione (GSH) levels and catalase and superoxide dismutase activities. Moreover, the authors observed the downregulation of apoptosis signal-regulating kinase 1, c-Jun N-terminal kinase 1 and protein 53 and the upregulation of insulin receptor substrate 1 in rat pancreas. Glucose transporter 4 was upregulated in rat muscle. Liquid chromatography and high-resolution mass spectrometry (LC-HRMS) analysis identified balanitin-2, hexadecenoic acid, methyl protodioscin and 26-(O-β-D-glucopyranosyl)-3-β-[4-O-(β-D-glucopyranosyl)-2-O-(α-L-rhamnopyranosyl)-β-D-glucopyranosyloxy]-22,26-dihydroxyfurost-5-ene in crude extract and balanitin-1 and trigonelloside C in butanol and dichloromethane fractions of crude extract. Ezzat et al. [40] isolated several compounds from pericarp, including stigmasterol-3-O-β-D-glucopyranoside (a), a pregnane glucoside: pregn-5-ene-3β,16β,20(R)-trio1-3-O-β-D-glucopyranoside (b); a furostanol saponin: 26-(O-β-D-glucopyranosyl)-22-O-methylfurost-5-ene-3β,26-diol-3-O-β-D-glucopyranosyl-(1→4)-[α-L-rhamnopyranosyl-(1→2)]-β-D-glucopyranoside (c). The latter component possessed significant α-glucodidase (AG) and aldose reductase inhibitory activities in streptozotocin-induced diabetic Wistar rats. Compound (c) also caused a significant increment in insulin and C-peptide levels.
4.7. Molluscicidal Activity
Regarding the effects of fruit extracts on juvenile and adult Bulinus globosus and B. truncatus, two Planorbid (ramshorn) freshwater snails have been reported [75]. LC95 values were 16.9 and 19.7 μg/mL and 14.2 and 12.0μg/mL for juvenile and adults of B. globosus and B. truncatus, respectively. Seed, endocarp, mesocarp and whole fruit extracts were assessed against adult Biomphalaria pfeifferi, another Planorbid snail, and Lymnaea natalensis, a Lymnaeid pond snail [76]. LC90 values were 77.70, 120.04, 89.50 and 97.55 mg/L against Biomphalariapfeifferi for seed, endocarp, mesocarp and whole fruit extracts, respectively, and 102.30, 138.21, 115.42 and 127.69 mg/L against Lymnaeanatalensis. Furthermore, the molluscicidal activity of seed oil on Monacha cartusiana, a Hygromiid land snail, has been demonstrated [77]. Bioactive compounds were identified as saponins, such as diosgenin, yamogenin and 3,5-spirostadiene.
4.8. Other Activities
Several studies demonstrate the antinematode and antiplasmodial activities of B. aegyptiaca extracts. Shalaby et al. [78] showed the effects of methanolic fruit extracts on enteral and parenteral stages of Trichinella spiralis (pork worm). The authors also evaluated the effectiveness of methanolic extract against preadult migrating larvae and encysted larvae of Trichinella spiralis in rats and compared them with the commonly used anthelmintic chemical, albendazole. Methanolic extract (1000 mg/kg body weight) for five successive days throughout the parasite lifecycle led to a marked reduction in migrating and encysted larvae by 81.7% and 61.7%, respectively. In another study, the efficacy of a methanolic extract on Toxocara vitulorum (roundworm), a major parasite in cattle and buffalo, was assessed [79]. They incubated parasites in a ringer solution containing 10, 30, 60, 120 and 240 µg/mL of ethanolic extract for 24 h. The most prominent activity at 240 µg/mL caused the disorganization of body cuticle musculature. Kusch et al. [17] evaluated a crude extract of seeds for antiplasmodial activity. An IC50 value for chloroquine-susceptible Plasmodium falciparum NF54 was 68.26 µg/µL. The compound responsible for this activity was 6-phenyl-2(H)-1,2,4-triazin-5-one oxime. The authors also showed that two phenolic compounds, 2,6-di-tert-butyl-phenol and 2,4-di-tert-butylphenol, displayed antiplasmodial activity at IC50 values of 50.29 and 47.82 µM, respectively.
5. Conclusions
B. aegyptiaca or desert date is an underutilized tree species. The nutritional status of the fruits, leaves and seeds indicate that this species could be exploited as a food source. Seed oil might also be a good source of biodiesel. Leaves and young shoots are nutritionally rich and could be exploited as cattle feed. Furthermore, fruits, leaves, roots and the bark of stem and roots are substantial sources of bioactive phytochemicals that display a host of possibly useful biological properties. B. aegyptiaca might prove to be a valuable source of bioactive agents for use in human and veterinary medicine.
Author Contributions
Conceptualization and methodology, H.N.M. and G.G.Y.; validation, H.N.M. and G.G.Y.; investigation, H.N.M., G.G.Y., Y.H.D. and A.I.; formal analysis H.N.M. and G.G.Y.; resources, H.N.M. and Y.H.D.; data curation, H.N.M., G.G.Y. and Y.H.D.; writing—original draft preparation, H.N.M. and G.G.Y.; writing—review and editing, H.N.M., Y.H.D. and A.I.; visualization, H.N.M., G.G.Y., Y.H.D. and A.I. All authors have read and agreed to the published version of the manuscript.
Funding
The authors extend their appreciation to the Deputyship for Research and Innovation, “Ministry of Education“ Saudi Arabia for funding this research work through the project number IFKSURP-59.
Acknowledgments
The authors thankful to the Researchers Support and Services Unit (RSSU) for their technical support.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Orwa, C.; Mutua, A.; Kindt, R.; Simons, A.; Jamnadass, R.H. Agroforestree Database: A Tree Reference and Selection Guide Version 4.0. World Agroforestry Centre, Kenya. 2009. Available online: http://apps.worldagroforestry.org/treedb2/speciesprofile.php?Spid=279 (accessed on 10 November 2020).
- Sarker, S.D.; Barholomew, B.; Nash, R.J. Alkaloids from Balanites aegyptiaca. Fitoterapia 2000, 71, 328–330. [Google Scholar] [CrossRef]
- Ojo, O.O.; Nadro, M.S.; Tell, I.O. Protection of rats by extracts of some common Nigerian trees against acetaminophen-induced hepatotoxicity. Afr. J. Biotechnol. 2006, 5, 755–760. [Google Scholar]
- IwuMaurice, M. Handbook of African Medicinal Plants; CRC Press: Boca Raton, FL, USA, 1993; p. 129. [Google Scholar]
- Achaglinkame, M.A.; Aderibigbe, R.O.; Hensel, O.; Sturm, B.; Korese, K. Nutritional characteristics of four underutilized wild fruits of dietary interest in Ghana. Foods 2019, 8, 104. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sagna, M.B.; Diallo, A.; Sarr, P.S.; Ndiaye, O.; Goffner, D.; Guisse, A. Biochemical composition and nutritional value of Balanites aegyptiaca(L.) Del fruit pulp from Northern Ferlo in Senegal. Afr. J. Biotechnol. 2014, 13, 336–342. [Google Scholar]
- Amadou, I. Date fruits: Nutritional composition of dates (Balanites aegyptiaca Delile and Phoenix dactylifera L.). In Nutritional Composition of Fruit Cultivars; Simmonds, M.S.J., Preedy, V.R., Eds.; Elsevier Inc.: Amsterdam, The Netherlands, 2016; pp. 215–233. [Google Scholar]
- Cook, J.A.; VanderJagt, D.J.; Pastuszyn, A.; Mounkaila, G.; Glew, R.S.; Glew, R.H. Nutrient content of two indigenous plant foods of western Sahel: Balanites aegyptiaca and Maerua crassifolia. J. Food Comp. Anal. 1998, 11, 221–230. [Google Scholar] [CrossRef]
- Samuel, A.L.; Temple, V.J.; Ladeji, O. Chemical and nutrition evaluation of the seed kernel of Balanites aegyptiaca. Niger. J. Biotechnol. 1997, 8, 57–63. [Google Scholar]
- Mohamed, A.M.; Wolf, W.; Spieb, W.E.L. Physical, morphological and chemical characteristics, oil recovery and fatty acid composition of Balanites aegyptiaca Del. kernels. Plant. Foods Hum. Nutr. 2002, 57, 179–189. [Google Scholar] [CrossRef]
- Chapagain, B.P.; Yehosha, Y.; Wiesman, Z. Desert date (Balanites aegyptiaca) as an arid lands sustainable bioresource for biodiesel. Bioresour. Technol. 2009, 100, 1221–1226. [Google Scholar] [CrossRef]
- Kubmarawa, D.; Andenyang, I.F.H.; Magomya, A.M. Amino acid profile of two non-conventional leafy vegetables, Sesamum indicum and Balanites aegyptiaca. Afr. J. Biotechnol. 2008, 7, 3502–3504. [Google Scholar]
- Khamis, G.; Saleh, A.M.; Habeeb, T.H.; Hozzein, W.N.; Wadaan, M.A.M.; Papernbrock, J.; AbdElgawad, H. Provenance effect on bioactive phytochemicals and nutritional and health benefits of the desert date Balanites aegyptiaca. J. Food Biochem. 2020, 44, e13229. [Google Scholar] [CrossRef]
- Becker, B. The contribution of wild plants to human nutrition in the Ferlo (Northern Senegal). Agrofor. Syst. 1983, 1, 257–267. [Google Scholar] [CrossRef]
- Li, A.N.; Li, S.; Zhang, Y.J.; Xu, X.R.; Chen, Y.M.; Li, H.B.; Li, H.B. Resources and biological activities of natural polyphenols. Nutrients 2014, 6, 6020–6047. [Google Scholar] [CrossRef] [PubMed]
- Tsao, R. Chemistry and biochemistry of dietary polyphenols. Nutrients 2010, 2, 1231–1246. [Google Scholar] [CrossRef] [PubMed]
- Kusch, P.; Deininger, S.; Specht, S.; Maniako, R.; Haubrich, S.; Pommerening, T.; Lin, P.K.T.; Hoerauf, A.; Kaiser, A. In vitro and in vivo antimalarial activity of seeds from Balanites aegyptiaca: Compounds of the extract show growth inhibition and activity against plasmodial aminopeptidase. J. Parsitol Res. 2011, 368692. [Google Scholar]
- Hassan, D.M.; Anigo, K.M.; Umar, I.A.; Alegbejo, J.O. Evaluation of phytoconstituent of Balanites aegyptiaca leaves and fruit-mesocarp extracts. M.O.J. Bioorg. Org. Chem. 2017, 1, 228–232. [Google Scholar]
- Meda, R.N.T.; Vlase, L.; Lamien-Meda, A.; Lamien, C.E.; Muntean, D.; Tiperciuc, B.; Oniga, I.; Nacoulma, O.G. Identification and quantification of phenolic compounds from Balanites aegyptiaca (L.) Del (Balanitaceae) galls and leaves by HPLC-MS. Nat. Prod. Res. 2011, 25, 93–99. [Google Scholar] [CrossRef]
- Farag, M.A.; Porzel, A.; Wessjohann, L.A. Unraveling the active hypoglycemic agent trigonelline in Balanites aegyptiaca date fruit using metabolite fingerprinting by NMR. J. Pharm. Biomed. Anal. 2015, 115, 383–387. [Google Scholar] [CrossRef]
- Maksoud, S.A.; Hadidi, M.N.E. The flavonoids of Balanites aegyptiaca (Balanitaceae) from Egypt. Plant. Syst. Evol. 1988, 160, 153–158. [Google Scholar] [CrossRef]
- Shafik, N.H.; Shafek, R.E.; Michel, H.N.; Eskander, E.F. Phytochemical study and antihyperglycemic effect of Balanites aegyptiaca kernel extract on alloxan induced diabetic male rat. J. Chem. Pharma. Res. 2016, 8, 128–136. [Google Scholar]
- Seida, A.A.; Kinghorn, A.D.; Cordell, G.A.; Farnsworth, N.R. Isolation of bergapten and marmesin from Balanites aegyptiaca. Planta Med. 1981, 43, 92–93. [Google Scholar] [CrossRef]
- Hosny, M.; Khalifa, T.; Calis, I.; Wright, A.D.; Sticher, O. Balanitoside, a furostanal glycoside and 6-methyl-diosgenin from Balanites aegyptiaca. Phytochemistry 1992, 31, 3565–3569. [Google Scholar] [CrossRef]
- Hardman, R.; Sofowora, E.A. Isolation and characterization of yamogenin from Balanites aegyptiaca. Phytochemistry 1970, 9, 645–649. [Google Scholar] [CrossRef]
- Samuelsson, G.; Farah, M.H.; Cleason, P.; Hagos, M.; Thulin, M.; Hedberg, O.; Warfa, A.M.; Hassan, A.O.; Elmi, A.H.; Abdurahman, A.D.; et al. Inventory of plants used in traditional medicine in Somalia. I. Plants of the families of Acanthaceae-Chenopodiaceae. J. Ethnopharmacol. 1991, 35, 25–63. [Google Scholar] [CrossRef]
- Ashaal, H.A.; Farghaly, A.A.; Abd El Aziz, M.M.; Ali, M.A. Phytochemical investigation and medicinal evaluation of fixed oil of Balanites aegyptiaca fruits (Balanitaceae). J. Ethnopharmacol. 2010, 127, 495–501. [Google Scholar] [CrossRef]
- Kamel, M.S.; Koskinen, A. Pregnane glycosides from fruits of Balanites aegyptiaca. Phytochemistry 1995, 40, 1773–1775. [Google Scholar] [CrossRef]
- Liu, H.W.; Naknishi, K. The structures of balanitins, potent molluscicides isolated from Balanites aegyptiaca. Tetrahedron 1982, 38, 513–519. [Google Scholar] [CrossRef]
- Pettit, G.R.; Doubek, D.L.; Herald, D.L.; Numata, A.; Takahasi, C.; Fujiki, R.; Miyamoto, T. Isolation and structure of cytostatic steroidal saponins from the African medicinal plant Balanites aegyptiaca. J. Nat. Prod. 1991, 54, 1491–1502. [Google Scholar] [CrossRef]
- Gnoula, C.; Megalizzi, V.; De Neve, N.; Sauvage, S.; Ribaucour, F.; Guissou, P.; Duez, P.; Dubois, J.; Ingrassia, L.; Lefranc, F.; et al. Balanitin-6 and −7: Diosgenyl saponins isolated from Balanites aegyptiaca Del. display significant anti-tumor activity in vitro and in vivo. Int. J. Oncol. 2008, 32, 5–15. [Google Scholar] [CrossRef]
- Beit-Yannai, E.; Ben-Shabat, S.; Goldschmidt, N.; Chapagain, B.P.; Liu, R.H.; Wiesman, Z. Antiproliferative activity of steroidal saponins from Balanites aegyptiaca—An in vtiro study. Phytochem. Lett. 2011, 4, 43–47. [Google Scholar] [CrossRef]
- Brimer, L.; ElSheik, S.H.; Furu, P. Preliminary investigation of the disposition of the molluscicidal saponin deltonin from Balanites aegyptiaca in a snail species (Biomphalaria glabrata) and in mice. J. Pestic. Sci. 2007, 32, 213–221. [Google Scholar] [CrossRef] [Green Version]
- Farid, H.; Haslinger, E.; Kunert, O.; Wegner, C.; Hamburger, M. New steroidal glycosides from Balanites aegyptiaca. Helv. Chim. Acta 2002, 85, 1019–1026. [Google Scholar] [CrossRef]
- Kamel, M.S. A furostanolsaponin from fruits of Balanites aegyptiaca. Phytochemistry 1998, 48, 755–757. [Google Scholar] [CrossRef]
- Al-Ghannam, S.M.; Hamdy-Ahamed, H.; Zein, N.; Zahran, F. Antitumor activity of balanitoside extracted from Balanites aegyptiaca fruit. J. Appl. Pharm. Sci. 2013, 3, 179–191. [Google Scholar]
- Staerk, D.; Chapagain, B.P.; Lindin, T.; Wiesman, Z.; Jaroszewski, J.W. Structural analysis of complex saponins of Balanites aegyptiaca by 800 MHz1H NMR spectroscopy. Magn. Reson. Chem. 2006, 44, 923–928. [Google Scholar] [CrossRef] [Green Version]
- Kamel, M.S.; Ohtani, K.; Kurokawa, T.; Assaf, M.H.; El-Shanawany, M.A.; Ali, A.A.; Kasai, R.; Ishibashi, S.; Tanaka, O. Studies on Balanites aegyptiaca fruits, an antidiabetic Egyptian folk medicine. Chem. Pharm. Bull. 1991, 39, 1229–1233. [Google Scholar] [CrossRef] [Green Version]
- Speroni, E.; Cervellati, R.; Innocenti, G.; Cost, S.; Guerra, M.C.; Dall Acqua, S.; Govani, P. Anti-inflammatory, anti-nociceptive and antioxidant activities of Balanites aegyptiaca (L.) Delile. J. Ethnopharmacol. 2005, 98, 117–125. [Google Scholar] [CrossRef]
- Ezzat, S.M.; Motaal, A.A.; Awdan, S.A.W.E. In vitro and in vivo antidiabetic potential of extracts and a furostanol saponin from Balanites aegyptiaca. Pharm. Biol. 2017, 55, 1931–1936. [Google Scholar] [CrossRef]
- Motaal, A.A.; El-Askary, H.; Crockett, S.; Kunert, O.; Sakr, B.; Shaker, S.; Grigore, A.; Albulescu, R.; Bauer, R. Aldose reductase inhibition of a saponin-rich fraction and new furostanol saponin derivatives from Balanites aegyptiaca. Phytomedicine 2015, 22, 829–836. [Google Scholar] [CrossRef]
- Seida, A.A. Isolation, Identification and Structure Elucidation of Cytotoxic and Antitumor Principles from Ailanthus integrifolia, Amyris pinnata and Balanites aegyptiaca. Ph.D. Thesis, University of Illinois, Chicago, IL, USA, 1979. [Google Scholar]
- Venugopala, K.N.; Rashmi, V.; Odhav, B. Review on natural coumarin lead compounds for their pharmacological activity. BioMed Res. Int. 2013, 2013, 963248. [Google Scholar] [CrossRef] [Green Version]
- Roberts, M.F.; Wink, M. Alkaloids-Biochemistry, Ecological Functions and Medical Applications; Plenum Press: New York, NY, USA, 1998. [Google Scholar]
- Panda, N.; Banerjee, S.; Mandal, N.B.; Sahu, N.P. Pregnane glycosides. Nat. Prod. Commun. 2006, 1, 665–695. [Google Scholar] [CrossRef]
- Challinor, V.L.; De Voss, J.J. Open-chain steroidal glycosides, a diverse class of plant saponins. Nat. Prod. Rep. 2013, 30, 429–454. [Google Scholar] [CrossRef] [PubMed]
- El Aziz, M.M.A.; Ashour, A.S.; Melad, A.S.G. A review on saponins from medicinal plants: Chemistry, isolation, and determination. J. Nanomed Res. 2019, 7, 282–288. [Google Scholar]
- Murthy, H.N.; Bapat, V.A. Importance of Underutilized Fruits and Nuts. In Bioactive Compounds in Underutilized Fruits and Nuts, Murthy, H., Bapat, V., Eds.; Reference Series in Phytochemistry; Springer: Cham, Switzerland, 2020; pp. 3–19. [Google Scholar]
- Hassan, L.E.A.; Dahham, S.S.; Saghir, S.A.M.; Mohammed, A.M.A.; Eltayeb, N.M.; Majis, A.M.S.A.; Majid, A.S.A. Chemotherapeutic potentials of the stem bark of Balanites aegyptiaca (L.) Delile: An antiangiogenic, antitumor and antioxidant agent. B.M.C. Complement. Altern. Med. 2016, 16, 396. [Google Scholar] [CrossRef] [Green Version]
- Khalil, N.S.A.; Abou-Elhamd, A.S.; Wasfy, S.I.A.; El Mileegy, I.M.H.; Hamed, M.Y.; Ageely, H.M. Antidiabetic and antioxidant impacts of desert date (Balanites aegyptiaca) and parsley (Petroselinum sativum) aqueous extracts: Lessons from experimental rats. J. Diabet. Res. 2016, 8408326. [Google Scholar]
- Kahsay, T.; Muluget, A.; Unnithan, C.R. Antioxidant and antibacterial activities of Balanites aegyptiaca Delil from Northern Ethiopia. Am. J. Pharma Tech. Res. 2014, 4, 415–422. [Google Scholar]
- Rasouli, H.; Farzaei, M.H.; Khodarahmi, R. Polyphenols and their benefits: A review. Int. J. Food Prop. 2017, 20, S1700–S1741. [Google Scholar] [CrossRef] [Green Version]
- Yoshida, Y.; Niki, E. Antioxidant effects of phytosterol and its components. J. Nutr. Sci. Vitaminol. 2003, 49, 277–280. [Google Scholar] [CrossRef]
- Cowan, M.M. Plant Products as Antimicrobial Agents. Clin. Microbiol. Rev. 1999, 12, 564–582. [Google Scholar] [CrossRef] [Green Version]
- Anani, K.; Adjrah, Y.; Ameyapoh, Y.; Karou, S.D.; Agbonon, A.; de Souza, C.; Gbeassor, M. Effects of hydroethanolic extracts of Balanites aegyptiaca (L.) Delile (Balanitaceae) on some resistant pathogens bacteria isolated from wounds. J. Ethnophamacol. 2015, 164, 16–21. [Google Scholar] [CrossRef]
- Chapagain, B.P.; Wiesman, Z.; Tsror (Lahkim), L. In vitro study of the antifungal activity of saponin-rich extracts against prevalent phytopathogenic fungi. Ind. Crop Prod. 2007, 26, 109–115. [Google Scholar] [CrossRef]
- Runyoro, D.K.B.; Matee, M.I.N.; Ngassapa, O.D.; Joseph, C.C.; Mbwambo, Z.H. Screening of Tanzanian medicinal plants for anti-candida activity. B.M.C. Complement. Altern. Med. 2006, 6, 11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thirupathi, K.; Krishna, D.R.; Ravi Kumar, B.; Apparao, A.V.N.; Krishna Mohan, G. Hepatoprotective effect of leaves of Balanitesroxburghii against carbon tetrachloride-induced hepatic damage in rats. Curr. Trends Biotechnol. Pharm. 2009, 3, 219–224. [Google Scholar]
- Mariam, S.; Onyenibe, N.S.; Oyelola, O.B. Aqueous extract of Balanites aegyptiaca Del Fruit mesocarp protects against CCl4—Induced liver damage in rats. Br. J. Pharm. Res. Int. 2013, 3, 917–928. [Google Scholar] [CrossRef]
- Jaiprakash, B.; Rajkumar, A.; Karadi, R.V.; Savadi, R.V.; Hukkeri, V.L. Hepatoprotective activity of bark of Balanites aegyptiaca Linn. J. Nat. Rem. 2003, 3, 205–207. [Google Scholar]
- Ganesan, K.; Jayachandran, M.; Xu, B. A critical review on hepatoprotective effects of bioactive food components. Crit. Rev. Food Sci. Nutr. 2018, 58, 1165–1229. [Google Scholar] [CrossRef] [PubMed]
- Choudhari, A.S.; Mandave, P.C.; Deshpande, M.; Ranjekar, P.; Prakash, O. Phytochemicals in cancer treatment: From preclinical studies to clinical practice. Front. Pharmacol. 2020, 10, 1614. [Google Scholar] [CrossRef] [Green Version]
- Grattan, B.J., Jr. Plant sterols as anticancer nutrients: Evidence for their role in breast cancer. Nutrients 2013, 5, 359–387. [Google Scholar] [CrossRef] [Green Version]
- Man, S.; Gao, W.; Zhang, Y.; Huang, L.; Liu, C. Chemical study and medical application of saponins as anti-cancer agents. Fitoterapia 2010, 81, 703–714. [Google Scholar] [CrossRef]
- Issa, N.M.; Mansour, F.K.; El-Safti, F.A.; Nooh, H.Z.; El-Sayed, I.H. Effect of Balanites aegyptiaca on Ehrlich Ascitic carcinoma growth and metastasis in Swiss mice. Exp. Toxicol. Pathol. 2015, 67, 435–441. [Google Scholar] [CrossRef]
- Yuan, G.; Wahlqvist, M.L.; He, G.; Yang, M.; Li, D. Natural products and anti-inflammatory activity. Asia Pac. J. Clin. Nutr. 2006, 15, 143–152. [Google Scholar]
- Kim, H.P.; Mani, I.; Iversen, L.; Ziboh, V.A. Effects of naturally occurring flavonoids and bioflavonoids on epidermal cyclooxygenase and lipoxygenase from guinea-pigs. Prostaglandins Leukot. Essent. Fatty Acids 1998, 58, 17–24. [Google Scholar] [CrossRef]
- Ahmed, M.M.; Eid, M.M. Evaluation of Balanites aegyptiaca oil as untraditional source of oil and its antiinflammatory activity. J. Drug Res. Egypt 2015, 36, 1–11. [Google Scholar]
- Grover, J.K.; Yadav, S.S.; Vats, V.J. Medicinal plants of India with anti-diabetic potential. J. Ethnopharmacol. 2002, 81, 81–100. [Google Scholar] [CrossRef]
- Gad, M.Z.; El-Sawalhi, M.M.; Ismail, M.F.; El-Tanbouly, N.D. Biochemical study of the antidiabetic action of the Egyptian plants Fenugreek and Balanites. Mol. Cell. Biochem. 2006, 281, 173–183. [Google Scholar] [CrossRef] [PubMed]
- Al-Malki, A.L.; Barbour, E.K.; Abulnaja, K.O.; Moselhy, S.S. Management of hyperglycaemia by ethyl acetate extract of Balanites aegyptiaca (Desert date). Molecules 2015, 20, 14425–14434. [Google Scholar] [CrossRef] [Green Version]
- Gnoula, C.; Guissou, P.; Duez, P.; Frederich, M.; Dubois, J. Nematocidal compounds from the seeds of Balanites aegyptiaca: Isolation and structure elucidation. Int. J. Phamacol. 2007, 3, 280–284. [Google Scholar]
- Makena, W.; Hamman, W.O.; Buraimoh, A.A.; Dibal, N.I.; Obaje, S.G. Therapeutic effects of balanitoside in streptozotocin-induced diabetic rats. J. Tiabah Univ. Med. Sci. 2018, 13, 402–406. [Google Scholar] [CrossRef]
- Hassanin, K.M.A.; Mahmoud, M.A.; Hassan, H.M.; Abdel-Razik, A.H.; Aziz, L.N.; Rateb, M.E. Balanites aegyptiaca ameliorates insulin secretion and decreases pancreatic apoptosis in diabetic rats: Role of SAPK/JNK pathway. Biomed. Pharmacother. 2018, 102, 1084–1091. [Google Scholar] [CrossRef] [Green Version]
- Anto, F.; Aryeetey, M.E.; Anyorigiya, T.; Asoala, V.; Kpikpi, J. The relative susceptibilities of juvenile and adult Bulinusglobosus and Bulinustruncatus to the molluscicidal activities in the fruit of Ghanaian Blighiasapida, Blighiaunijugata and Balanites aegyptiaca. Ann. Trop. Med. Parasitol. 2005, 99, 211–217. [Google Scholar] [CrossRef]
- Molla, E.; Giday, M.; Erko, B. Laboratory assessment of the molluscicidal and cercariacidal activities of Balanites aegyptiaca. Asian Pac. J. Trop. Biomed. 2013, 3, 657–662. [Google Scholar] [CrossRef] [Green Version]
- Dawidar, A.E.M.; Mortada, M.M.; Raghib, H.M.; Abdel-Mogib, M. Molluscicidal activity of Balanites aegyptiaca against Monacha cartusiana. Pharma. Biol. 2012, 50, 1326–1329. [Google Scholar] [CrossRef] [PubMed]
- Shalaby, M.A.; Moghazy, F.M.; Shalaby, H.A.; Nasr, S.M. Effect of methanoic extract of Balanites aegyptiaca fruits on enteral and parenteral stages of Trichonella spiralis in rats. Parasitol. Res. 2010, 107, 17–25. [Google Scholar] [CrossRef] [PubMed]
- Shalaby, H.A.; El-Namaky, A.H.; Khalil, F.A.; Kandil, O.M. Efficacy of methonolic extract of Balanites aegyptiaca fruits on Toxocara vitulorum. Vet. Parasitol. 2012, 183, 386–392. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Morphology of Balanites aegyptiaca (L.) Delile: (A) habit, (B) leaves, (C) stem bark, (D) ripened fruits, (E) rind (left) and pulp (right), (F) seed kernels and (G) seed oil.
Figure 1.
Morphology of Balanites aegyptiaca (L.) Delile: (A) habit, (B) leaves, (C) stem bark, (D) ripened fruits, (E) rind (left) and pulp (right), (F) seed kernels and (G) seed oil.

Figure 2.
Structures of phenolic compounds isolated from desert date.

Figure 3.
Structures of flavonoids and coumarins isolated from desert date.

Figure 4.
Structures of alkaloids, steroids and pregnane glycosides isolated from desert date.

Figure 5.
Structures of spirostanolsaponins isolated from desert date.

Figure 6.
Structures of furostanoland open-chain steroidal saponins of desert date.

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Murthy, H.N.; Yadav, G.G.; Dewir, Y.H.; Ibrahim, A. Phytochemicals and Biological Activity of Desert Date (Balanites aegyptiaca (L.) Delile). Plants 2021, 10, 32. https://doi.org/10.3390/plants10010032
AMA Style
Murthy HN, Yadav GG, Dewir YH, Ibrahim A. Phytochemicals and Biological Activity of Desert Date (Balanites aegyptiaca (L.) Delile). Plants. 2021; 10(1):32. https://doi.org/10.3390/plants10010032
Chicago/Turabian StyleMurthy, Hosakatte Niranjana, Guggalada Govardhana Yadav, Yaser Hassan Dewir, and Abdullah Ibrahim. 2021. "Phytochemicals and Biological Activity of Desert Date (Balanites aegyptiaca (L.) Delile)" Plants 10, no. 1: 32. https://doi.org/10.3390/plants10010032
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.