
Allelopathy of the Medicinal Plant Dregea volubilis (L.f.) Benth. ex Hook.f. and Its Phytotoxic Substances with Allelopathic Activity


Abstract
:
1. Introduction
2. Materials and Methods
2.1. Plant Material and Extract Preparation
2.2. Growth Bioassay
2.3. Separation, Purification, and Identification of the Active Substances
2.4. Bioassay of the Isolated Active Substances
2.5. Statistical Analysis
3. Results
3.1. Allelopathic Effects of the D. volubilis Extracts
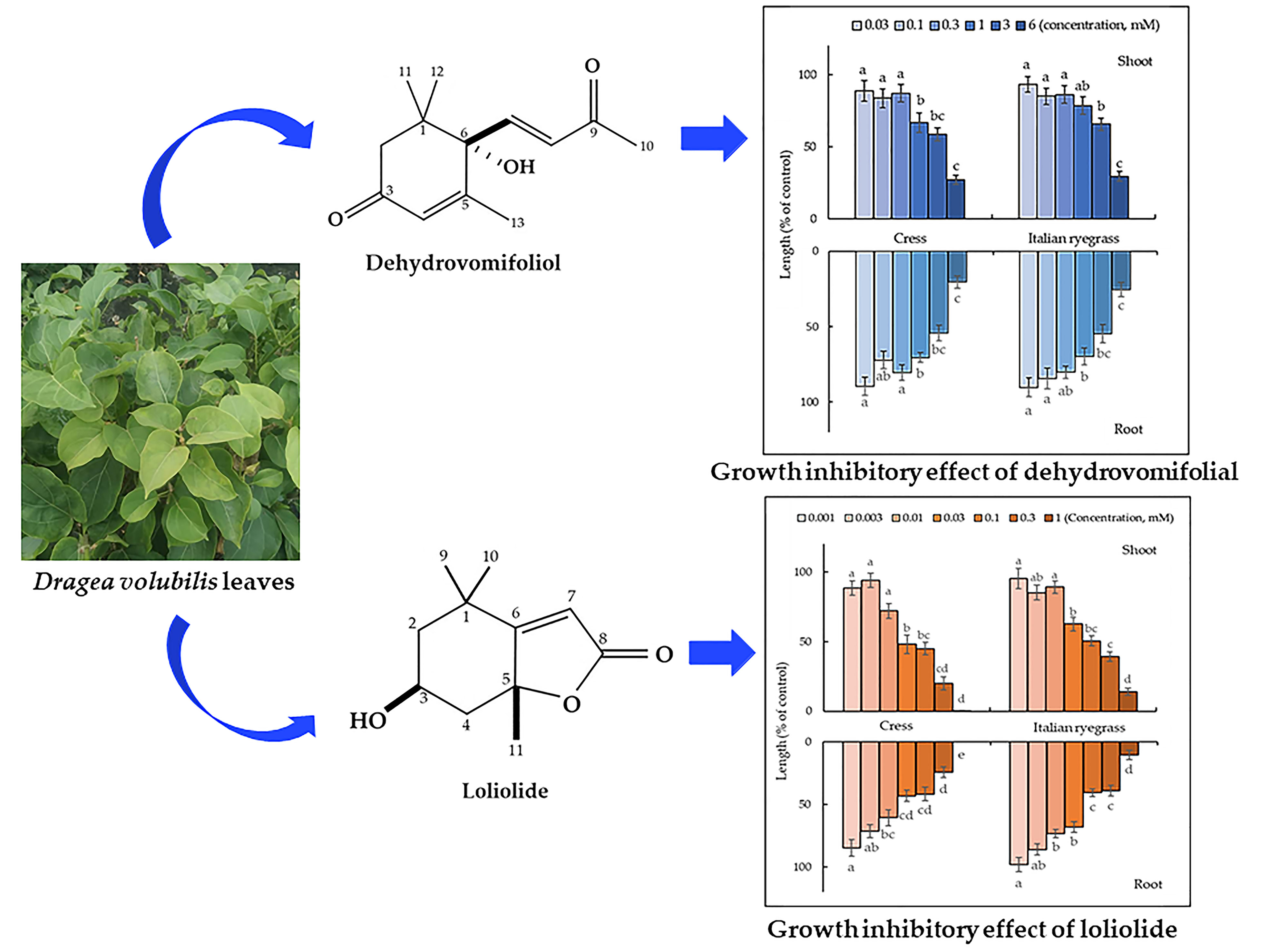
3.2. Isolation and Characterization of the Active Substances
3.3. Biological Activity of the Two Identified Compounds
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Marwat, K.B.; Khan, M.A.; Hashim, S.; Nawab, K.; Khattak, A.M. Integrated weed management in wheat. Pak. J. Bot. 2011, 43, 625–633. [Google Scholar]
- Hussain, M.I.; González, L.; Reigosa, M. Phytotoxic effect of allelochemicals and herbicides on photosynthesis, growth and carbon isotope discrimination in Lactuca sativa. Allelopath. J. 2010, 22, 441–449. [Google Scholar]
- Moss, S.; Ulber, L.; Hoed, I.D. A herbicide resistance risk matrix. Crop Prot. 2019, 115, 13–19. [Google Scholar] [CrossRef]
- Heap, I. International Survey of Herbicide Resistant Weeds. Available online: http://www.weedscience.org (accessed on 21 May 2020).
- Scavo, A.; Abbate, C.; Mauromicale, G. Plant allelochemicals: Agronomic, nutritional and ecological relevance in the soil system. Plant Soil. 2019, 442, 23–48. [Google Scholar] [CrossRef]
- Haig, T. Allelochemicals in plants. In Allelopathy in Sustainable Agriculture and Forestry; Zeng, R.S., Mallik, A.U., Luo, S.M., Eds.; Springer: New York, NY, USA, 2008; pp. 63–104. [Google Scholar]
- Cheng, F.; Cheng, Z. Research progress on the use of plant allelopathy in agriculture and the physiological and ecological mechanisms of allelopathy. Front. Plant Sci. 2015, 6, 1020. [Google Scholar] [CrossRef] [PubMed]
- Radhakrishnan, R.; Alqarawi, A.A.; Abd Allah, E.F. Bioherbicides: Current knowledge on weed control mechanism. Ecotoxicol. Environ. Saf. 2018, 158, 131–138. [Google Scholar] [CrossRef]
- Soltys, D.; Krasuska, U.; Bogatek, R.; Gniazdowska, A. Allelochemicals as bioherbicides—Present and perspectives. In Herbicides—Current Research and Case Studies in Use; Intech: Rijeka, Croatia, 2013; pp. 518–534. [Google Scholar]
- Duke, S.O.; Dayan, F.E.; Romagni, J.G.; Rimando, A.M. Natural products as sources of herbicide, current status and future trends. Weed Res. 2000, 40, 99–111. [Google Scholar] [CrossRef]
- Macías, F.A.; Molinillo, J.M.G.; Varela, R.M.; Galindo, J.G.G. Allelopathy—A natural alternative for weed control. Pest Manag. Sci. 2007, 63, 327–348. [Google Scholar] [CrossRef]
- Wink, M. Modes of action of herbal medicines and plant secondary metabolites. J. Med. 2015, 2, 251–286. [Google Scholar] [CrossRef]
- Islam, A.M.; Yeasmin, S.; Qasem, J.R.S.; Juraimi, A.S.; Anwar, M.P. Allelopathy of medicinal plants: Current status and future prospects in weed management. J. Agric. Sci. 2018, 9, 1569–1588. [Google Scholar] [CrossRef] [Green Version]
- Poonpaiboonpipat, T.; Krumsri, R.; Kato-Noguchi, H. Allelopathic and herbicidal effects of crude extract from Chromolaena odorata (L.) R.M.King and H.Rob. on Echinochloa crus-galli and Amaranthus viridis. Plants 2021, 10, 1609. [Google Scholar] [CrossRef] [PubMed]
- Kato-Noguchi, H.; Salam, M.A.; Ohno, O.; Suenaga, K. Nimbolide B and nimbic acid B, phytotoxic substances in neem leaves with allelopathic activity. Molecules 2014, 19, 6929–6940. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krumsri, R.; Ozaki, K.; Teruya, T.; Kato-Noguchi, H. Isolation and identification of two potent phytotoxic substances from Afzelia xylocarpa for controlling weeds. Appl. Sci. 2021, 11, 3542. [Google Scholar] [CrossRef]
- Rob, M.M.; Hossen, K.; Khatun, M.R.; Iwasaki, K.; Iwasaki, A.; Suenaga, K.; Kato-Noguchi, H. Identification and Application of Bioactive Compounds from Garcinia xanthochymus Hook. for Weed Management. Appl. Sci. 2021, 11, 2264. [Google Scholar] [CrossRef]
- Ladhari, A.; Omezzine, F.; Dellagreca, M.; Zarrelli, A.; Haouala, R. Phytotoxic activity of Capparis spinosa L. and its discovered active compounds. Allelopath. J. 2013, 32, 175–190. [Google Scholar]
- Endress, M.E.; Bruyns, P.V. A revised classification of the Apocynaceae sl. Bot. Rev. 2000, 66, 1–56. [Google Scholar] [CrossRef] [Green Version]
- Rahman, A.H.M.M.; Akter, M. Taxonomy and traditional medicinal uses of apocynaceae (Dogbane) family of Rajshahi District, Bangladesh. Res. Rev. J. Bot. Sci. 2015, 4, 1–12. [Google Scholar]
- Silveira, D.; de Melo, A.F.; Magalhães, P.O.; Fonseca-Bazzo, Y.M. Tabernaemontana species: Promising sources of new useful drugs. Stud. Nat. Prod. Chem. 2017, 54, 227–289. [Google Scholar]
- Naidoo, C.M.; Naidoo, Y.; Dewir, Y.H.; Murthy, H.N.; El-Hendawy, S.; Al-Suhaibani, N. Major bioactive alkaloids and biological activities of Tabernaemontana Species (Apocynaceae). Plants 2021, 10, 313. [Google Scholar] [CrossRef]
- Gulzar, A.; Siddiqui, M.B.; Bi, S. Phenolic acid allelochemicals induced morphological, ultrastructural, and cytological modification on Cassia sophera L. and Allium cepa L. Protoplasma 2016, 253, 1211–1221. [Google Scholar] [CrossRef]
- Bari, I.N.; Kato-Noguchi, H. Phytotoxic effects of Cerbera manghas L. leaf extracts on seedling elongation of four monocot and four dicot test species. Acta Agrobot. 2017, 70, 1–7. [Google Scholar] [CrossRef]
- Krumsri, R.; Boonmee, S.; Kato-Noguchi, H. Evaluation of the allelopathic potential of leaf extracts from Dischidia imbricata (Blume) Steud. on the seedling growth of six test plants. Not. Bot. Horti Agrobot. 2019, 47, 1019–1024. [Google Scholar] [CrossRef] [Green Version]
- Hossain, E.; Sarkar, D.; Maiti, A.; Chatterjee, M.; Mandal, S.C.; Gupta, J.K. Anti-inflammatory effect of a methanolic extract of leaves of Dregea volubilis. J. Ethnopharmacol. 2010, 132, 525–528. [Google Scholar] [CrossRef] [PubMed]
- Biswas, M.; Bhattacharya, S.; Mukhopadhyay, R.; Haldar, P.K. Dregea volubilis (L.f.) Benth. (Asclepiadaceae): An appraisal on pharmacognostic, phytochemical and pharmacological studies. Orient. Pharm. Exp. Med. 2018, 18, 1–8. [Google Scholar] [CrossRef]
- Hossain, E.; Chakroborty, S.; Milan, A.; Chattopadhyay, P.; Mandal, S.C.; Gupta, J.K. In vitro and in vivo antitumor activity of a methanol extract of Dregea volubilis leaves with its antioxidant effect. Pharm. Biol. 2012, 50, 338–343. [Google Scholar] [CrossRef]
- Chatterjee, A.; Pakrashi, S.C. The Treatise on Indian Medicinal Plants; Publication and Information Directorate: New Delhi, India, 1995; Volume 4, pp. 131–132. [Google Scholar]
- Rajadurai, M.; Vidhya, V.G.; Ramya, M. Ethno-medicinal plants used by the traditional healers of Pacchamalai hills, Tamil Nadu, India. J. Ethnobiol. Ethnomed. 2009, 3, 39–41. [Google Scholar]
- DeFilipps, R.A.; Krupnick, G.A. The medicinal plants of Myanmar. PhytoKeys 2018, 102, 1–341. [Google Scholar] [CrossRef]
- Biswas, M.; Bikash, M.N.; Palit, P.; Kumar, G.A.; Bannerjee, S.; Kanti, H.P. In vitro anti-leishmanial and anti-tumour activities of a pentacyclic triterpenoid compound isolated from the fruits of Dregea volubilis Benth Asclepiadaceae. Trop. J. Pharm. Res. 2009, 8, 127–131. [Google Scholar]
- Sahu, N.; Panda, N.; Mandal, N.B.; Banerjee, S.; Koike, K.; Nikaido, T. Polyoxypregnane glycosides from the flowers of Dregea volubilis. Phytochemistry 2002, 61, 383–388. [Google Scholar] [CrossRef]
- Panda, N.; Mandal, D.; Mandal, N.B.; Sahu, N.P.; Banerjee, S. Flavonoid and flavone C-glycosides from Dregea volubilis. Nat. Prod. Commun. 2006, 1, 731–733. [Google Scholar] [CrossRef]
- Thuy, N.T.K.; Phuong, P.T.; Mai, N.T.; Hien, T.T.T.; Cuc, N.T.; Tai, B.H.; Van Kiem, P.; Van Minh, C.; Kang, K.S.; Kim, S.H.; et al. Pregnane glycosides from Dregea volubilis and their α-glucosidase inhibitory activity. Phytochem. Lett. 2020, 37, 90–94. [Google Scholar] [CrossRef]
- Das, B.; De, A.; Das, M.; Das, S.; Samanta, A. A new exploration of Dregea volubilis flowers: Focusing on antioxidant and antidiabetic properties. S. Afr. J. Bot. 2017, 109, 16–24. [Google Scholar] [CrossRef]
- Biswas, M.; Biswas, K.; Ghosh, A.K.; Haldar, P.K. A pentacyclic triterpenoid possessing analgesic activity from the fruits of Dregea volubilis. Pharmacogn. Mag. 2009, 5, 90. [Google Scholar]
- Shankar, K.R.; Das, S.; Bujala, P. Phytochemical screening and in vitro antibacterial activity of ethanol and aqueous extracts of Dregea volubilis leaves. Biosci. Biotechnol. Res. Asia 2010, 7, 975–979. [Google Scholar]
- Purushoth, P.T.; Maheswaran, V.S.; Selvakumari, S.; Suriyapadminimoka, R.S.; Dileep, G. An antioxidant and anti- bacterial activity of Dregea volubilis leaves extract. Pharm. Lett. 2012, 4, 525–529. [Google Scholar]
- Natarajan, V.; Dhas, A.S.A.G. Effect of active fraction isolated from the leaf extract of Dregea volubilis [Linn.] Benth. on plasma glucose concentration and lipid profile in streptozotocin-induced diabetic rats. Springer Plus 2013, 2, 394. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kato, T.; Tsunakawa, M.; Sasaki, N.; Aizawa, H.; Fujita, K.; Kitahara, Y.; Takahashi, N. Growth and germination inhibitors in rice husks. Phytochemistry 1977, 16, 45–48. [Google Scholar] [CrossRef]
- Kai, H.; Baba, M.; Okuyama, T. Two new megastigmanes from the leaves of Cucumis sativus. Chem. Pharm. Bull. 2007, 55, 133–136. [Google Scholar] [CrossRef] [Green Version]
- Hodges, R.; Porte, A.L. The structure of loliolide: A terpene from Lolium perenne. Tetrahedron 1964, 20, 1463–1467. [Google Scholar] [CrossRef]
- Valdes, L.J. Loliolide from Salvia divinorum. J. Nat. Prod. 1986, 49, 171. [Google Scholar] [CrossRef]
- Kimura, J.; Maki, N. New loliolide derivatives from the brown alga Undaria pinnatifida. J. Nat. Prod. 2002, 65, 57–58. [Google Scholar] [CrossRef] [PubMed]
- Boonmee, S.; Kato-Noguchi, H. Allelopathic activity of Acacia concinna pod extracts. Emir. J. Food. Agric. 2017, 29, 250–255. [Google Scholar] [CrossRef] [Green Version]
- El-Mergawi, R.; El-Desoki, E.R. Allelopathic activities of celery extract and its fractions against Corchorus olitorius, Echinochloa crusgalli and Portulaca oleracea weeds. Adv. Hortic. Sci. 2018, 32, 503–510. [Google Scholar]
- Hossen, K.; Iwasaki, A.; Suenaga, K.; Kato-Noguchi, H. Phytotoxic activity and growth inhibitory substances from Albizia richardiana (Voigt.) King & Prain. Appl. Sci. 2021, 11, 1455. [Google Scholar]
- Rob, M.M.; Iwasaki, A.; Suzuki, R.; Suenaga, K.; Kato-Noguchi, H. Garcienone, a novel compound involved in allelopathic activity of Garcinia Xanthochymus hook. Plants 2019, 8, 301. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tuyen, P.T.; Xuan, T.D.; Anh, T.T.T.; Van, T.M.; Ahmad, A.; Elzaawely, A.A.; Khanh, T.D. Weed suppressing potential and isolation of potent plant growth inhibitors from Castanea crenata Sieb. et Zucc. Molecules 2018, 23, 345. [Google Scholar] [CrossRef] [Green Version]
- Rob, M.M.; Kato-Noguchi, H. Study of the allelopathic activity of Garcinia pedunculata Roxb. Plant Omics 2019, 12, 31–36. [Google Scholar] [CrossRef]
- Boonmee, S.; Suwitchayanon, P.; Krumsri, R.; Kato-Noguchi, H. Investigation of the allelopathic potential of Nephrolepis cordifolia (L.) C. Presl against dicotyledonous and monocotyledonous plant species. Environ. Control Biol. 2020, 58, 71–78. [Google Scholar] [CrossRef]
- Kyaw, E.H.; Kato-Noguchi, H. Assessment of allelopathic activity of Tradescantia spathacea Sw. for weed control. Biol. Futur. 2021, 72, 489–495. [Google Scholar] [CrossRef]
- Pan, L.; Sinden, M.R.; Kennedy, A.H.; Chai, H.; Watson, L.E.; Graham, T.L.; Kinghorn, A.D. Bioactive constituents of Helianthus tuberosus (Jerusalem artichoke). Phytochem. Lett. 2009, 2, 15–18. [Google Scholar] [CrossRef]
- Macías, F.A.; Oliva, R.M.; Varela, R.M.; Torres, A.; Molinillo, J.M. Allelochemicals from sunflower leaves cv. Peredovick. Phytochemistry 1999, 52, 613–621. [Google Scholar] [CrossRef]
- Kim, I.; Chin, Y.W.; Lim, S.W.; Kim, Y.C.; Kim, J. Norisoprenoids and hepatoprotective flavone glycosides from the aerial parts of Beta vulgaris var. cicla. Arch. Pharm. Res. 2004, 27, 600–603. [Google Scholar] [CrossRef] [PubMed]
- Cutillo, F.; D’Abrosca, B.; Dellagreca, M.; Fiorentino, A.; Zarrelli, A. Terpenoids and phenol derivatives from Malva silvestris. Phytochemistry 2006, 67, 481–485. [Google Scholar] [CrossRef] [PubMed]
- Machado, F.B.; Yamamoto, R.E.; Zanoli, K.; Nocchi, S.R.; Novello, C.R.; Schuquel, I.T.A.; Sakuragui, C.M.; Luftmann, H.; Ueda-Nakamura, T.; Nakamura, C.V.; et al. Evaluation of the antiproliferative activity of the leaves from Arctium lappa by a bioassay-guided fractionation. Molecules 2012, 17, 1852–1859. [Google Scholar] [CrossRef]
- Yang, Y.; Bakri, M.; Gu, D.; Aisa, H.A. Separation of (s)-dehydrovomifoliol from leaves of Nitraria sibirica Pall. by high-speed counter-current chromatography. J. Liq. Chromatogr. Relat. Technol. 2012, 36, 573–582. [Google Scholar] [CrossRef]
- Fiorucci, L.; Mantovanelli, G.C.; Bernardi, D.I.; Silva, A.A.; de Oliveira, R.S., Jr.; Ishii-Iwamoto, E.L.; Sarragiotto, M.H.; Baldoqui, D.C. Isolation of the constituents and evaluation of allelopathic potential of Raphanus sativus L. (Brassicaceae). Planta Med. 2014, 80, P1L61. [Google Scholar] [CrossRef]
- Mayer, H. Synthesis of optically active carotenoids and related compounds. Pure Appl. Chem. 1979, 51, 535–564. [Google Scholar] [CrossRef] [Green Version]
- Dietz, H.; Winterhalter, P. Phytotoxic constituents from Bunias orientalis leaves. Phytochemistry 1996, 42, 1005–1010. [Google Scholar] [CrossRef]
- Zhou, B.; Kong, C.H.; Li, Y.H.; Wang, P.; Xu, X.H. Crabgrass (Digitaria sanguinalis) allelochemicals that interfere with crop growth and the soil microbial community. J. Agric. Food Chem. 2013, 61, 5310–5317. [Google Scholar] [CrossRef]
- Bich, T.T.N.; Kato-Noguchi, H. Isolation and identification of a phytotoxic substance from the emergent macrophyte Centrostachys aquatica. Bot. Stud. 2014, 55, 1–5. [Google Scholar] [CrossRef] [Green Version]
- Kato-Noguchi, H.; Moriyasu, M.; Ohno, O.; Suenaga, K. Growth limiting effects on various terrestrial plant species by an allelopathic substance, loliolide, from water hyacinth. Aquat. Bot. 2014, 117, 56–61. [Google Scholar] [CrossRef]
- Islam, M.S.; Iwasaki, A.; Suenaga, K.; Kato-Noguchi, H. Isolation and identification of two potential phytotoxic substances from the aquatic fern Marsilea crenata. J. Plant Biol. 2017, 60, 75–81. [Google Scholar] [CrossRef]
- Okunade, A.L.; Wiemer, D.F. (-)-Loliolide, an ant-repellent compound from Xanthoxyllum setulosum. J. Nat. Prod. 1985, 48, 472–473. [Google Scholar] [CrossRef]
- Ragasa, C.Y.; De Luna, R.D.; Hofilena, J.G. Antimicrobial terpenoids from Pterocarpus indicus. Nat. Prod. Res. 2005, 19, 305–309. [Google Scholar] [CrossRef] [PubMed]
- Lu, H.; Xie, H.; Gong, Y.; Wang, Q.; Yang, Y. Secondary metabolites from the seaweed Gracilaria lemaneiformis and their allelopathic effects on Skeletonema costatum. Biochem. Syst. Ecol. 2011, 39, 397–400. [Google Scholar] [CrossRef]
- Yang, X.; Kang, M.; Lee, K.; Kang, S.; Lee, W.; Jeon, Y. Antioxidant activity and cell protective effect of loliolide isolated from Sargassum ringgoldianum subsp. coreanum. Algae 2011, 26, 201–208. [Google Scholar] [CrossRef]
- Pan, W.; Liu, K.; Guan, Y.; Tan, G.T.; Van Hung, N.; Cuong, N.M.; Soejarto, D.D.; Pezzuto, J.M.; Fong, H.H.S.; Hongjie, Z. Bioactive compounds from Vitex leptobotrys. J. Nat. Prod. 2014, 77, 663–667. [Google Scholar] [CrossRef] [Green Version]
- Yuan, Z.; Zheng, X.; Zhao, Y.; Liu, Y.; Zhou, S.; Wei, C.; Hu, W.; Shao, H. Phytotoxic compounds isolated from leaves of the invasive weed Xanthium spinosum. Molecules 2018, 23, 2840. [Google Scholar] [CrossRef] [Green Version]
- Zaman, F.; Iwasaki, A.; Suenaga, K.; Kato-Noguchi, H. Two allelopathic substances from Paspalum commersonii Lam. Acta Agric. Scand. Sect. B Plant Soil Sci. 2018, 68, 342–348. [Google Scholar]
- Cueva, C.; Moreno-Arribas, M.V.; Martinez-Alvarez, P.J.; Bills, G.; Vicente, M.F.; Basilio, A.; Lopez Rivas, C.; Requena, T.; Rodríguez, J.M.; Bartolomé, B. Antimicrobial activity of phenolic acids against commensal, probiotic and pathogenic bacteria. Res. Microbiol. 2010, 16, 372–382. [Google Scholar] [CrossRef]
- Sanchez-Maldonado, A.F.; Schieber, A.; Ganzle, M.G. Structure-function relationships of the antibacterial activity of phenolic acids and their metabolism by lactic acid bacteria. J. Appl. Microbiol. 2011, 111, 1176–1184. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, M.; Ueda, M.; Furumoto, T.; Kawanami, Y. Retarding activity of 6-O-acyl-D-alloses against plant growth. Biosci. Biotechnol. Biochem. 2010, 74, 216–217. [Google Scholar] [CrossRef] [PubMed] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Target Plant Species | I50 Value | Correlation Coefficient (r) | ||
|---|---|---|---|---|
| Shoot | Root | Shoot | Root | |
| Aqueous methanol extracts (mg D.W. equivalent extract/mL) | Aqueous methanol extracts | |||
| Italian ryegrass | 10.15 a | 5.32 b | −0.764 *** | −0.792 *** |
| Cress | 5.60 b | 3.21 c | −0.849 *** | −0.895 *** |
| Dehydrovomifolial (mM) | Dehydrovomifolial | |||
| Italian ryegrass | 4.60 a | 3.59 bc | −0.603 *** | −0.677 *** |
| Cress | 3.79 b | 3.24 c | −0.715 *** | −0.747 *** |
| Loliolide (mM) | Loliolide | |||
| Italian ryegrass | 0.102 a | 0.073 b | −0.918 *** | −0.888 *** |
| Cress | 0.029 c | 0.022 c | −0.870 *** | −0.878 *** |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kyaw, E.H.; Iwasaki, A.; Suenaga, K.; Kato-Noguchi, H. Allelopathy of the Medicinal Plant Dregea volubilis (L.f.) Benth. ex Hook.f. and Its Phytotoxic Substances with Allelopathic Activity. Agronomy 2022, 12, 303. https://doi.org/10.3390/agronomy12020303
Kyaw EH, Iwasaki A, Suenaga K, Kato-Noguchi H. Allelopathy of the Medicinal Plant Dregea volubilis (L.f.) Benth. ex Hook.f. and Its Phytotoxic Substances with Allelopathic Activity. Agronomy. 2022; 12(2):303. https://doi.org/10.3390/agronomy12020303
Chicago/Turabian StyleKyaw, Ei Han, Arihiro Iwasaki, Kiyotake Suenaga, and Hisashi Kato-Noguchi. 2022. "Allelopathy of the Medicinal Plant Dregea volubilis (L.f.) Benth. ex Hook.f. and Its Phytotoxic Substances with Allelopathic Activity" Agronomy 12, no. 2: 303. https://doi.org/10.3390/agronomy12020303