
Annual Net Primary Productivity of Different Functional Groups as Affected by Different Intensities of Rainfall Reduction in the Semiarid Grasslands of the Gauteng Province in South Africa



Abstract
:1. Background
2. Materials and Methods
2.1. Site Description
2.2. Experimental Design and Data Collection
3. Statistical Analysis
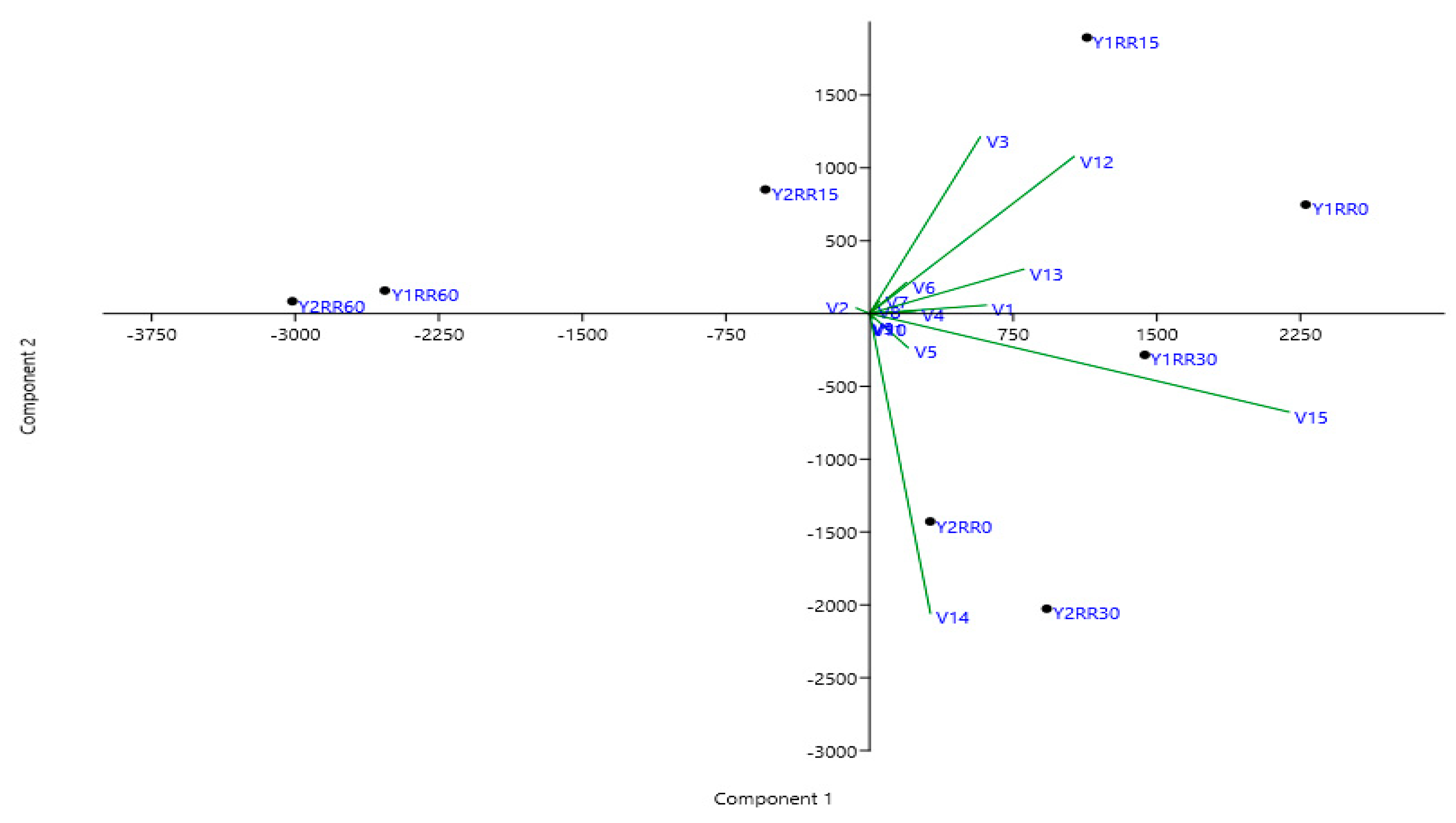
4. Results
5. Discussion
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

References
- IPCC. Climate Change 2013: The Physical Science Basis. Contribution of Working Group 1 to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change; Stocker, T.F., Qing, D., Plattner, G.-K., Tingor, M., Allen, S.K., Boschung, J., Nauels, A., Xia, Y., Bex, V., Midgley, P.M., Eds.; Cambridge University Press: Cambridge, UK; New York, NY, USA, 2013. [Google Scholar]
- Van Loon, A.F. Hydrological Drought Explained. WIREs Water 2015, 2, 359–392. [Google Scholar] [CrossRef]
- Tfwala, C.M.; van Rensburg, L.D.; Schall, R.; Dlamini, P. Drought dynamics and interannual rainfall variability on the Ghaap plateau, South Africa, 1918–2014. Phys. Chem. Earth 2018, 107, 1–7. [Google Scholar] [CrossRef]
- Dai, A. Drought under global warming: A review. WIREs Clim. Chang. 2011, 2, 45–65. [Google Scholar] [CrossRef] [Green Version]
- IPCC. Climate Change 2014: Synthesis Report. Contribution of Working Groups I, II and III to the Fifth Assessment Report of the Interovernmental Panel on Climate Change; Core Writing Team, Pachauri, R.K., Meyer, L.A., Eds.; IPCC: Geneva, Switzerland, 2014; p. 151. [Google Scholar]
- Knapp, A.K.; Hoover, D.L.; Wilcox, K.R.; Avolio, M.L.; Koerner, S.E.; La Pierre, K.J.; Loik, M.; Luo, Y.; Sala, O.E.; Smith, M.D. Characterizing differences in precipitation regimes of extreme wet and dry years: Implications for climate change experiments. Glob. Chang. Biol. 2015, 21, 2624–2633. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hui, D.; Yu, C.L.; Deng, Q.; Dzantor, E.K.; Zhou, S.; Dennis, S.; Sauve, R.; Johnson, T.L.; Fay, P.A.; Shen, W.; et al. Effects of precipitation changes on switchgrass photosynthesis, growth, and biomass: A mesocosm experiment. PLoS ONE 2018, 13, e0192555. [Google Scholar] [CrossRef]
- Huntington, T.G. Evidence for intensification of the global water cycle: Review and synthesis. J. Hydrol. 2006, 319, 83–95. [Google Scholar] [CrossRef]
- Fay, P.A.; Carlisle, J.D.; Knapp, A.K.; Blair, J.M.; Collins, S. Productivity responses to altered rainfall patterns in a C4 dominated grassland. Oecologia 2003, 137, 245–251. [Google Scholar] [CrossRef]
- Manabe, S.; Wetherald, R.T.; Milly, P.C.D.; Delworth, T.L.; Stouffer, R.J. Century-scale change in water availability: CO2-quadrupling experiment. Clim. Chang. 2004, 64, 59–76. [Google Scholar] [CrossRef]
- Volk, M.; Niklaus, P.A.; Korner, C. Soil moisture effects determine CO2 responses of grassland species. Oecologia 2000, 125, 380–388. [Google Scholar] [CrossRef]
- IPCC. International Panel of Climatic Changes; Fourth Assessment Report (AR4); IPCC: New York, NY, USA, 2007. [Google Scholar]
- Volaire, F.; Barkaoui, K.; Norton, M. Designing resilient and sustainable grasslands for a drier future: Adaptive strategies, functional traits and biotic interactions. Eur. J. Agron. 2014, 52, 81–89. [Google Scholar] [CrossRef] [Green Version]
- Trnka, M.; Olesen, J.E.; Kersebaum, K.C.; Skjelvag, A.O.; Eitzinger, J.; Seguin, B.; Peltonen-Sainio, P.; Rotter, R.; Iglesias, A.; Orlandini, S.; et al. Agroclimatic conditions in Europe under climate change. Glob. Chang. Biol. 2011, 17, 2298–2318. [Google Scholar] [CrossRef] [Green Version]
- Ciais, P.; Reichstein, M.; Viovy, N.; Granier, A.; Ogee, J.; Allard, V.; Aubinet, M.; Buchmann, N.; Bernhofer, C.; Carrara, A.; et al. Europe-wide reduction in primary productivity caused by the heat and drought in 2003. Nature 2005, 437, 529–533. [Google Scholar] [CrossRef]
- Lelievre, F.; Seddaiu, G.; Ledda, L.; Porqueddu, C.; Volaire, F. Water use efficiency and drought survival in Mediterranean perennial forage grasses. Field Crop Res. 2011, 121, 333–342. [Google Scholar] [CrossRef]
- Soussana, J.F.; Graux, A.I.; Tubiello, F.N. Improving the use of modelling for projections of climate change impacts on crops and pastures. J. Exp. Bot. 2010, 61, 2217–2228. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Magandana, T.P.; Hassen, A.; Tesfamariam, E.H. Seasonal Herbaceous Structure and Biomass Production Response to Rainfall Reduction and Resting Period in the Semi-Arid Grassland Area of South Africa. Agronomy 2020, 10, 1807. [Google Scholar] [CrossRef]
- Kleine, S.; Weissinger, L.; Müller, C. Impact of drought on plant populations of native and invasive origins. Oecologia 2017, 183, 9–20. [Google Scholar] [CrossRef] [PubMed]
- Norton, M.R.; Lelievre, F.; Fukai, S.; Volaire, F. Measurement of summer dormancy in temperate perennial pasture grasses. Austratian J. Agric. Res. 2008, 59, 498–509. [Google Scholar] [CrossRef]
- Norton, M.R.; Volaire, F.; Lelievre, F.; Fukai, S. Identification and measurement of summer dormancy in temperate perennial grasses. Crop Sci. 2009, 49, 2347–2352. [Google Scholar] [CrossRef]
- Gioria, M.; Osborne, B.A. Resource competition in plant invasions: Emerging patterns and research needs. Front. Plant Sci. 2014, 5, 501. [Google Scholar] [CrossRef] [Green Version]
- Jury, M.R. Climate trends in southern Africa. S. Afr. J. Sci. 2013, 109, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Soil Classification Working Group. Soil Classification: A Taxonomic System for South Africa; Department of Agricultural Development: Pretoria, South Africa, 1991.
- Talore, D.G. Quantitative and Qualitative Herbage Yield, and Carbon Sequestration in Subtropical Grasslands Subjected to Different Precipitation, Grazing Management and Land Use Type. Ph.D. Thesis, University of Pretoria, Pretoria, South Africa, 2016; p. 90. [Google Scholar]
- Van Oudtshoorn, F. Guide to Grasses of Southern Africa, 3rd ed.; Briza Publications: Pretoria, South Africa, 2012; ISBN 978-1-920217-35-8. [Google Scholar]
- SAS (Statistical Analysis System). User’s Guide; SAS/STAT® 9.2; SAS Institute Inc.: Cary, NC, USA, 2014. [Google Scholar]
- Hoover, D.L.; Knapp, A.K.; Smith, M.D. Resistance and resilience of a grassland ecosystem to climate extremes. Ecology 2014, 95, 2646–2656. [Google Scholar] [CrossRef] [Green Version]
- Nippert, J.B.; Knapp, A.K. Soil water partitioning contributes to species coexistence in tallgrass prairie. Oikos 2007, 116, 1017–1029. [Google Scholar] [CrossRef]
- Prevéy, J.S.; Seastedt, T.R. Effects of precipitation change and neighboring plantson population dynamics of Bromus tectorum. Oecologia 2015, 179, 765–775. [Google Scholar] [CrossRef]
- Gilo, B.N.; Kelkay, T.Z. Changes in vegetation structure and aboveground biomass in response to traditional rangelandmanagement practices in Borana, southern Ethiopia. Afr. J. Range Forage Sci. 2017, 34, 21–31. [Google Scholar] [CrossRef]
- Greco, S.A.; Cavagnaro, J.B. Effects of drought in biomass production and allocation in three varieties of Trichloris crinite P. (Poaceae) a forage grass from the arid Monte region of Argentina. Plant Ecol. 2002, 164, 125–135. [Google Scholar] [CrossRef]
- Heitschmidt, R.K.; Klement, K.D.; Haferkamp, M.R. Interactive effects of drought and grazing on northern Great Plains rangelands. Rangel. Ecol. Manag. 2005, 58, 11–19. [Google Scholar] [CrossRef]
- Power, S.A.; Barnett1, K.L.; Ochoa-Hueso, R.; Facey, S.L.; Gibson-Forty, E.V.J.; Hartley, S.E.; Nielsen, U.N.; Tissue, D.T.; Johnson, S.N. DRI-Grass: A New Experimental Platform for Addressing Grassland Ecosystem Responses to Future Precipitation Scenarios in South-East Australia. Front. Plant Sci. 2016, 7, 1373. [Google Scholar] [CrossRef] [Green Version]
- Van Der Weijde, T.; Huxley, L.M.; Hawkins, S.; Sembiring, E.H.; Farrar, K.; Dolstra, O.; Visser, G.F.R.; Trindade, L.M. Impact of drought stress on growth and quality of miscanthus for biofuel production. Glob. Chang. Biol. 2017, 9, 770–782. [Google Scholar] [CrossRef] [Green Version]
- Sala, O.E.; Parton, W.J.; Joyce, L.A.; Lauenroth, W.K. Primary production of the central grassland region of the United States. Ecology 1988, 69, 40–45. [Google Scholar] [CrossRef]
- Hsu, J.S.; Powell, J.; Adler, P.B. Sensitivity of mean annual primary production to precipitation. Glob. Chang. Biol. 2012, 18, 2246–2255. [Google Scholar] [CrossRef]
- Southon, G.E.; Green, E.R.; Jones, A.G.; Barker, C.G.; Power, S.A. Long-term nitrogen additions increase likelihood of climate stress and affect recovery from wildfire in a low land heath. Glob. Chang. Biol. 2012, 18, 2824–2837. [Google Scholar] [CrossRef] [PubMed]
- Koerner, S.E.; Collins, S.L. Interactive effects of grazing, drought, and fire on grassland plant communities in North America and South Africa. Ecology 2014, 95, 2014. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lenhart, P.A.; Eubanks, M.D.; Behmer, S.T. Water stress in grasslands: Dynamic responses of plants and insect herbivores. Oikos 2015, 124, 381–390. [Google Scholar] [CrossRef]
- Briggs, J.M.; Knapp, A.K. Determinants of C3 forb growth and production in a C4 dominated grassland. Plant Ecol. 2001, 152, 93–100. [Google Scholar] [CrossRef]
- Nippert, J.B.; Knapp, A.K.; Briggs, J.M. Intra-annual rainfall variability and grassland productivity: Can the past predict the future? Plant Ecol. 2006, 184, 65–74. [Google Scholar] [CrossRef]
- Gherardi, L.A.; Sala, O.E. Enhanced precipitation variability decreases grass- and increases shrub-productivity. Proc. Natl. Acad. Sci. USA 2015, 112, 12735–12740. [Google Scholar] [CrossRef] [Green Version]
- Sala, O.E.; Lauenroth, W.K.; Parton, W.J. Long term soil water dynamics in the shortgrass steppe. Ecology 1992, 73, 1175–1181. [Google Scholar] [CrossRef]
- Jackson, R.B.; Canadell, J.; Ehleringer, J.R.; Mooney, H.A.; Sala, O.E.; Schulze, E.D. A global analysis of root distributions for terrestrial biomes. Oecologia 1996, 108, 389–411. [Google Scholar] [CrossRef]
- Weltzin, J.F.; McPherson, G.R. Implications of precipitation redistribution for shifts in temperate savanna ecotones. Ecology 2000, 81, 1902–1913. [Google Scholar] [CrossRef]
- Sala, O.E.; Golluscio, R.A.; Lauenroth, W.K.; Soriano, A. Resource partitioning between shrubs and grasses in the Patagonian Steppe. Oecologia 1989, 81, 501–505. [Google Scholar] [CrossRef] [PubMed]
- Moore, G.A.; Sanford, P.; Wiley, T. Perennial Pastures for Western Australia; Bulletin 4690; Department of Agriculture and Food: Perth, Australia, 2006.
- Van den Berg, L.; Zeng, Y.J. Response of South African indigenous grass species to drought stress induced by polyethylene glycol (PEG) 6000. S. Afr. J. Bot. 2006, 72, 284–286. [Google Scholar] [CrossRef] [Green Version]
- Fernández, R.J.; Reynolds, J.F. Potential growth and drought tolerance of eight desert grasses: Lack of a trade-off? Oecologia 2000, 123, 90–98. [Google Scholar] [CrossRef] [PubMed]
- Snyman, H.A.; du Preez, C.C. Rangeland degradation in a semi-arid South Africa—II: Influence on soil quality. J. Arid Environ. 2005, 60, 483–507. [Google Scholar] [CrossRef]
- Snyman, H.A.; van Rensburg, W.L.J. Short-term effect of severe drought on veld condition and water use efficiency of grassveld in the central Orange Free State. Afr. J. Range Sci. 1990, 7, 249–256. [Google Scholar]
- Xu, Z.; Zhou, G. Responses of photosynthetic capacity to soil moisture gradient in perennial rhizome grass and perennial bunchgrass. BMC Plant Biol. 2011, 11, 21. [Google Scholar] [CrossRef] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Scientific Names | Functional Group | Life Form | Growth | Ecological Status | Grazing Value | Intensity of Rainfall Reduction (%) | |||
|---|---|---|---|---|---|---|---|---|---|
| Form | 0 | 15 | 30 | 60 | |||||
| Aristida congesta | Grass | WP | Tufted | Increaser II | Low | + | ++ | ||
| Brachiaria serrata | Grass | P | Tufted | Decreaser | Ave | + | |||
| Melinis repens | Grass | P | Tufted | Increaser II | Low | + | + | ||
| Cynodon dactylon | Grass | P | Creeping | Increaser II | High | + | ++++ | +++++ | |
| Digitaria eriantha | Grass | P | Tufted | Decreaser | High | +++++ | +++++ | ++++ | +++ |
| Elephantorrhiza elephantina | Forb | P | - | Shrub | Low | + | ++++ | ||
| Rhynchosia monophylla | Forb | P | - | Increaser II | Low | + | |||
| Eragrostis curvula | Grass | P | Tufted | Increaser II | Eve | +++ | ++++ | +++++ | +++++ |
| Heteropogon contortus | Grass | P | Tufted | Increaser II | Ave | +++ | + | +++ | +++++ |
| Hyparrhenia hirta | Grass | P | Tufted | Increaser I | Low | + | |||
| Hypoxis hemorocallida | Forb | A | - | Invader | Low | + | + | ||
| Ipomoea crassipes | Forb | A | - | Invader | Low | ++ | + | ++++ | +++++ |
| Nidorella resedifolia | Forb | A | - | Invader | Low | ++ | ++ | ++++ | ++++ |
| Setaria nigrirostris | Grass | P | Tufted | Increaser II | Low | ++++ | ++ | +++++ | |
| Setaria sphacelata | Grass | P | Tufted | Decreaser | High | +++++ | +++++ | +++++ | ++ |
| Themeda triandra | Grass | P | Tufted | Decreaser | High | ++++ | + | ++++ | |
| Verbena officinalis | Forb | A | - | Invader | Low | ++ | |||
| Intensity of Rain Reduction | Annual Net Primary Productivity (kg DM/ha) | ||||
|---|---|---|---|---|---|
| Grass ANPP | |||||
| Overall ANPP | Forbs ANPP | Decreaser Grass ANPP | Increaser Grass ANPP | Overall Grass ANPP | |
| 0% | 4979 a | 1666 a | 1841 a | 1472 ab | 3312 a |
| 15% | 3761 a | 316 b | 2029 a | 1416 ab | 3445 a |
| 30% | 5029 a | 2066 a | 1147 a | 1816 a | 2963 a |
| 60% | 1716 b | 846 a | 220 b | 650 b | 869 b |
| S.E | 658.8 | 312.75 | 500.06 | 327.05 | 562.38 |
| P-value | 0.0068 | <0.0001 | 0.0553 | 0.0598 | 0.0020 |
| Year | |||||
| 2017 | 4184 | 760 b | 1673 | 1752 a | 3424 a |
| 2018 | 3558 | 1688 a | 946 | 925 b | 1870 b |
| S.E | 465.9 | 221.1 | 353.6 | 231.3 | 397.7 |
| P-value | 0.3488 | 0.0462 | 0.4608 | 0.0226 | 0.0188 |
| Rainfall Reduction | % Contribution in Terms of ANPP | |||
|---|---|---|---|---|
| Grass ANPP | ||||
| Forbs | Decreaser | Increaser | Overall Grass | |
| 0% | 36 a | 33 | 31 | 64 b |
| 15% | 6 b | 44 | 50 | 94 a |
| 30% | 42 a | 22 | 36 | 58 b |
| 60% | 48 a | 17 | 35 | 52 b |
| S.E | 7.4 | 8.3 | 8.8 | 7.0 |
| P-value | <0.0001 | 0.2540 | 0.5341 | 0.0010 |
| Year | ||||
| 2017 | 23 b | 32 | 45 | 77 a |
| 2018 | 43 a | 26 | 31 | 57 b |
| S.E | 5.2 | 5.8 | 6.2 | 5.1 |
| P-value | 0.0222 | 0.7639 | 0.0877 | 0.0042 |
| Grass Species (%) | |||||||
|---|---|---|---|---|---|---|---|
| Intensity of Rainfall Reduction (%) | Eragrostis curvula Log | Cynodon dactylon Sqrt | Heteropogon contortus | Digitaria eriantha Log | Setaria nigrirostris Log | Themeda triandra | Setaria spacelata |
| 0 | 37.9 | 0.2 b | 7.7 | 23.0 | 12.3 a | 0.2 | 0.9 |
| 15 | 37.2 | 1.2 b | 9.7 | 40.6 | 4.5 bc | 0.1 | 1.7 |
| 30 | 54.5 | 0.9 b | 5.5 | 16.0 | 5.1 ab | 0.2 | 0.8 |
| 60 | 40.8 | 18.7 a | 1.5 | 30.3 | 0c | 0 | 3.4 |
| S.E | 11.64 | 4.3 | 3.18 | 8.92 | 3.47 | 0.06 | 1.41 |
| P-value | 0.2206 | 0.0253 | 0.3880 | 0.4242 | 0.0036 | 0.1248 | 0.5684 |
| Year | |||||||
| 2017 | 48.6 | 5.5 | 5.5 | 28.4 | 4.6 | 0.1 | 1.2 |
| 2018 | 36.6 | 5.0 | 6.8 | 26.6 | 6.3 | 0.1 | 2.1 |
| S.E | 8.23 | 3.04 | 2.25 | 6.31 | 2.45 | 0.04 | 1.03 |
| P-value | 0.5852 | 0.9586 | 0.8621 | 0.5919 | 0.9152 | 0.9335 | 0.5269 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Magandana, T.P.; Hassen, A.; Tesfamariam, E.H. Annual Net Primary Productivity of Different Functional Groups as Affected by Different Intensities of Rainfall Reduction in the Semiarid Grasslands of the Gauteng Province in South Africa. Agronomy 2021, 11, 730. https://doi.org/10.3390/agronomy11040730
Magandana TP, Hassen A, Tesfamariam EH. Annual Net Primary Productivity of Different Functional Groups as Affected by Different Intensities of Rainfall Reduction in the Semiarid Grasslands of the Gauteng Province in South Africa. Agronomy. 2021; 11(4):730. https://doi.org/10.3390/agronomy11040730
Chicago/Turabian StyleMagandana, Thabo Patrick, Abubeker Hassen, and Eyob H. Tesfamariam. 2021. "Annual Net Primary Productivity of Different Functional Groups as Affected by Different Intensities of Rainfall Reduction in the Semiarid Grasslands of the Gauteng Province in South Africa" Agronomy 11, no. 4: 730. https://doi.org/10.3390/agronomy11040730