Association of Morphological, Ecological, and Genetic Diversity of Aerva javanica Populations Growing in the Eastern Desert of Egypt


Abstract
:1. Introduction
2. Materials and Methods
2.1. The Study Area and Plant Material
2.2. Soil Sampling and Analysis
2.3. DNA Extraction and ISSR-PCR Analysis
2.4. Statistical Analysis
3. Results
3.1. Genetic Diversity among Aerva javanica Populations based on ISSR Fingerprinting
3.2. Principal Coordinate Analysis (PCoA)
3.3. Soil Analysis
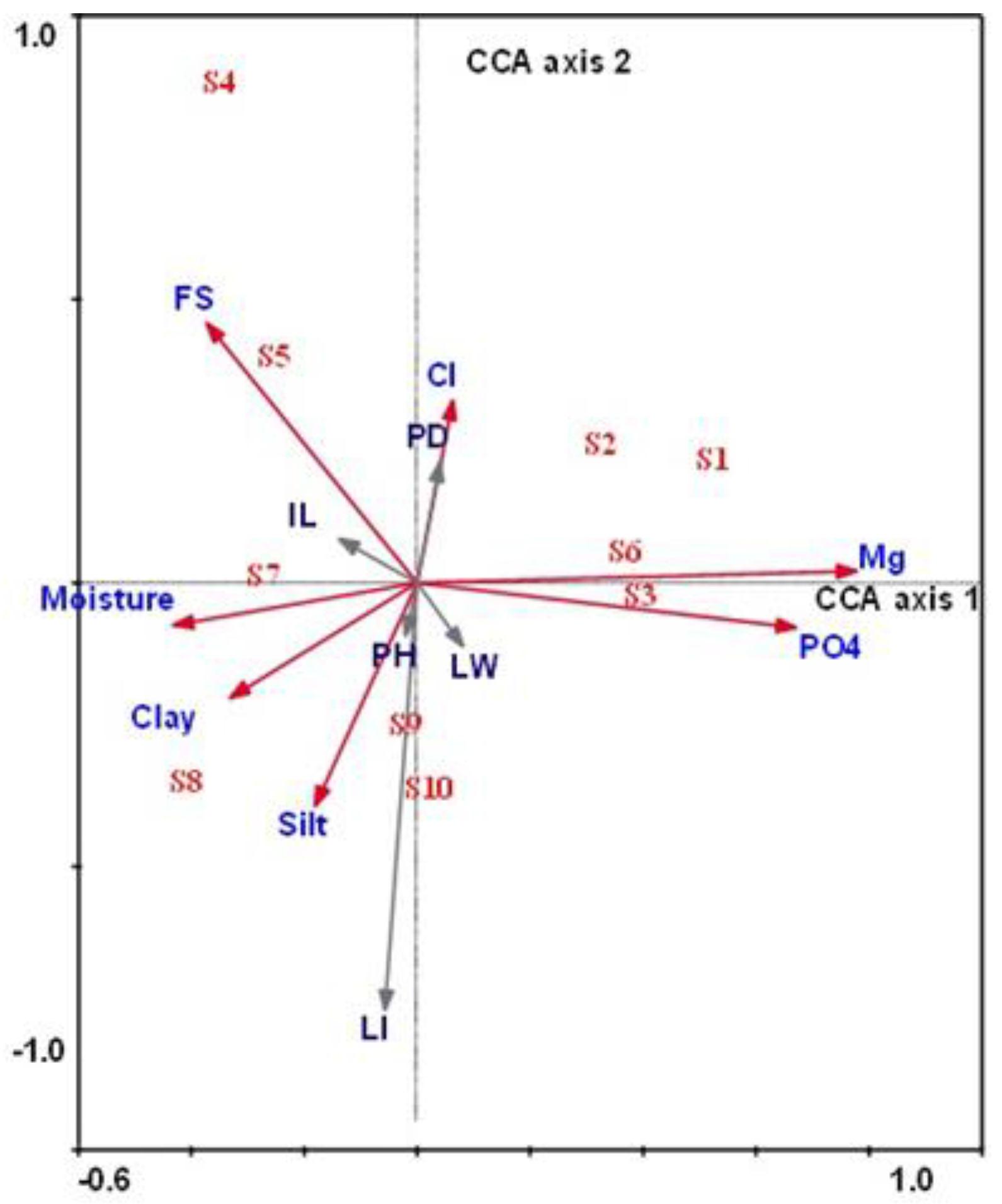
3.4. Compositional Ordination of Morphological Traits of Aerva javanica and Their Soil Variables
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Judd, W.S.; Campbell, C.S.; Kellog, E.A.; Stevens, P.F.; Donoghue, M.J. Plant Systematics: A Phylogenetic Approach, 3rd ed.; Sinauer Associates: Sunderland, MA, USA, 2007; ISBN 978-0-87893-407-2. [Google Scholar]
- Soliman, M.A. Cytogenetical studies on Aerva javanica (Amaranthaceae). Flora Mediterr. 2006, 16, 333–339. [Google Scholar]
- Boulos, L. Flora of Egypt Checklist; Revised Annotated Edition; Al Hadara Publishing: Cairo, Egypt, 2009. [Google Scholar]
- Raza, M.A.; Younas, M.; Buerkert, A.; Schlecht, E. Ethno-botanical remedies used by pastoralists for the treatment of livestock diseases in Cholistan desert, Pakistan. J. Ethnopharmacol. 2014, 151, 333–342. [Google Scholar] [CrossRef]
- Al-Shehri, M.; Moustafa, M. Anticancer, antibacterial, and phytochemicals derived from extract of Aerva javanica (Burm.f.) Juss. ex Schult. grown naturally in Saudi Arabia. Trop. Conserv. Sci. 2019, 12, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Movaliya, V.; Zaveri, M. Pharmacognostical studies on roots of Aerva javanica. Int. J. Pharm. Sci. Rev. Res. 2012, 16, 34–37. [Google Scholar]
- Soliman, M.S.A.; El-Tarras, A.; El-Awady, M.A. Cytologenetical Study of some Wild Plants from Taif, Saudi Arabia. Nat. Sci. 2011, 9, 101–104. [Google Scholar]
- Salama, F.; Abd El-Ghani, M.; El-Tayeh, N.; Amro, A.; El-Naggar, S. Adaptive responses of Aerva javanica Burm.f. Shult. to severe aridity in the Egyptian deserts. Egypt. J. Bot. 2018, 58, 171–184. [Google Scholar] [CrossRef] [Green Version]
- Salama, F.M.; El-Naggar, S.M. Phytosociology of wadi system west of Qusseir province. Feddes Repert. 1991, 102, 453–468. [Google Scholar] [CrossRef]
- Abd El-Ghani, M.M. Environmental correlates of species distribution in arid desert ecosystems of eastern Egypt. J. Arid Environ. 1998, 38, 297–313. [Google Scholar] [CrossRef]
- Shawky, R.A.; El-Khouly, A.A. Plant species diversity of some wadis at Red Sea Coast, Egypt. Environ. Resour. Ecol. J. 2017, 1, 1–14. [Google Scholar]
- Abd El-Ghani, M.M. Floristics and environmental relations in two extreme desert zones of western Egypt. Glob. Ecol. Biogeogr. 2000, 9, 499–515. [Google Scholar] [CrossRef]
- Abd El-Ghani, M.M.; Amer, W.M. Soil-vegetation relationships in a coastal desert plain of southern Sinai, Egypt. J. Arid Environ. 2003, 55, 607–628. [Google Scholar] [CrossRef]
- Salama, F.M.; Abd El-Ghani, M.M.; El-Tayeh, N.A.; Amro, A.; Abdrabbu, H.S. Correlations between soil variables and weed communities in major crops of the desert reclaimed lands in southern Egypt. Rend. Lincei 2017, 28, 363–378. [Google Scholar] [CrossRef]
- Salama, F.; Abd El-Ghani, M.; Gadalla, M.; Ramadan, T.; Galal, H.; Gaafar, A. Vegetation Patterns and Floristic Composition along Elevation Gradient on Jabal Musa, South Sinai, Egypt. Catrina Int. J. Environ. Sci. 2018, 17, 41–57. [Google Scholar]
- Ottewell, K.M.; Bickerton, D.C.; Byrne, M.; Lowe, A.J. Bridging the gap: A genetic assessment framework for population-level threatened plant conservation prioritization and decision-making. Divers. Distrib. 2016, 22, 174–188. [Google Scholar] [CrossRef] [Green Version]
- Tharachand, C.; Immanuel Selvaraj, C.; Mythili, M.N. Molecular markers in characterization of medicinal plants: An overview. Res. Plant Biol. 2012, 2, 1–12. [Google Scholar]
- Krishnan, P.N.; Decruse, S.W.; Radha, R.K. Conservation of medicinal plants of Western Ghats, India and its sustainable utilization through in vitro technology. Vitr. Cell. Dev. Biol. Plant. 2011, 47, 110–122. [Google Scholar] [CrossRef]
- Ganie, S.H.; Upadhyay, P.; Das, S.; Prasad Sharma, M. Authentication of medicinal plants by DNA markers. Plant Gene. 2015, 4, 83–99. [Google Scholar] [CrossRef] [Green Version]
- Zietkiewicz, E.; Rafalski, A.; Labuda, D. Genome fingerprinting by simple sequence repeat (SSR)-anchored polymerase chain reaction amplification. Genomics 1994, 20, 176–183. [Google Scholar] [CrossRef]
- Godwin, I.D.; Aitken, E.A.B.; Smith, L.W. Application of inter simple sequence repeat (ISSR) markers to plant genetics. Electrophoresis 1997, 18, 1524–1528. [Google Scholar] [CrossRef]
- Singh, D.R.; Srivastava, A.K.; Srivastava, A.; Srivastava, R.C. Genetic diversity among three Morinda species using RAPD and ISSR markers. Indian J. Biotechnol. 2011, 10, 285–293. [Google Scholar]
- Badr, A.; El-Shazly, H.H.; Helail, N.S.; El Ghanim, W. Genetic diversity of Artemisia populations in central and north Saudi Arabia based on morphological variation and RAPD polymorphism. Plant Syst. Evol. 2012, 298, 871–886. [Google Scholar] [CrossRef]
- Badr, A.; Morsy, W.; Abdelfattah, S.; Shams, S.; Shehab, A. Genetic diversity in Artemisia monosperma and Artemisia judaica populations in Egypt based on morphological, karyological and molecular variations. J. Med. Plants Res. 2012, 6, 66–78. [Google Scholar]
- Badr, A.; Kamel, M.; Zaki, H. Genetic diversity of Colocynth (Citrullus colocynthis Schrader) populations in the Eastern Desert of Egypt as revealed by morphological variation and ISSR polymorphism. Feddes Repert. 2018, 129, 173–184. [Google Scholar] [CrossRef]
- Abd El-Ghani, M.; Salama, F.; Salem, B.; El-hadidy, A.; Abdel-aleem, M. Phytogeography of the Eastern Desert flora of Egypt. Wulfenia 2017, 24, 97–120. [Google Scholar]
- Ryan, J.; Rashid, A. Application of soil and plant analysis for applied research and development in West Asia-North Africa: An international center’s perspective. Commun. Soil Sci. Plant Anal. 2006, 37, 2185–2198. [Google Scholar] [CrossRef]
- Sparks, D.L. Methods of Soil Analysis; Chemical Methods; Soil Science Society of America, Inc.: Madison, WI, USA, 1996. [Google Scholar]
- Estefan, G.; Sommer, R.; Ryan, J. Methods of Soil, Plant, and Water Analysis; A manual for the West Asia and North Africa Region; International Center for Agricultural Research in the Dry Areas (ICARDA): Aleppo, Syria, 2013. [Google Scholar]
- Watanabe, F.S.; Olsen, S.R. Test of an Ascorbic Acid Method for Determining Phosphorus in Water and NaHCO3 Extracts from Soil1. Soil Sci. Soc. Am. J. 1965, 29, 677. [Google Scholar] [CrossRef]
- Green, S.; Salkind, N. Using SPSS for Windows and Macintosh; Books a La Carte; Pearson: London, UK, 2016. [Google Scholar]
- Wilkinson, L. Systat. Wiley Interdiscip. Rev. Comput. Stat. 2010, 2, 256–257. [Google Scholar] [CrossRef]
- Kovach, W. MVSP-A MultiVariate Statistical Package for Windows, version 3.1; Kovach Computing Services: Pentraeth, Wales, UK, 1999. [Google Scholar]
- Ter Braak, C.J.F. Canonical correspondence analysis: A new eigenvector technique for multivariate direct gradient analysis. Ecology 1986, 67, 1167–1179. [Google Scholar] [CrossRef] [Green Version]
- Ter Braak, C.J.F. Update Notes: CANOCO, version 3.1; Agricultural Mathematics Group: Wageningen, The Netherlands, 1990; p. 35. [Google Scholar]
- Lee, S.R.; Choi, J.E.; Lee, B.Y.; Yu, J.N.; Lim, C.E. Genetic diversity and structure of an endangered medicinal herb: Implications for conservation. AoB Plants 2018, 10, ply021. [Google Scholar] [CrossRef]
- Carvalho, A.; Lima-Brito, J.; MaçÃs, B.; Guedes-Pinto, H. Genetic diversity and variation among botanical varieties of old Portuguese wheat cultivars revealed by ISSR assays. Biochem. Genet. 2009, 47, 276–294. [Google Scholar] [CrossRef]
- Gichimu, B.M.; Owuor, B.O.; Mwai, G.N.; Dida, M.M. Morphological characterization of some wild and cultivated watermelon (Citrullus sp.) accessions in Kenya. ARPN J. Agric. Biol. Sci. 2009, 4, 10–18. [Google Scholar]
- Vavilov, N.I. Origin and Geography of Cultivated Plants; Cambridge University Press: Cambridge, UK, 1992; p. 498. ISBN 0-521-40427-4. [Google Scholar]
- Kumar, A.; Mishra, P.; Singh, S.C.; Sundaresan, V. Efficiency of ISSR and RAPD markers in genetic divergence analysis and conservation management of Justicia adhatoda L., a medicinal plant. Plant Syst. Evol. 2014, 300, 1409–1420. [Google Scholar] [CrossRef]
- El-Domyati, F.M.; Younis, R.A.A.; Edris, S.; Mansour, A.; Sabir, J.; Bahieldin, A. Molecular markers associated with genetic diversity of some medicinal plants in Sinai. J. Med. Plants Res. 2011, 5, 1918–1929. [Google Scholar]
- Joshi, S.P.; Gupta, V.S.; Aggarwal, R.K.; Ranjekar, P.K.; Brar, D.S. Genetic diversity and phylogenetic relationship as revealed by inter simple sequence repeat (ISSR) polymorphism in the genus Oryza. Theor. Appl. Genet. 2000, 100, 1311–1320. [Google Scholar] [CrossRef]
- Rawashdeh, I.M.; Haddad, N.I.; Amri, A. Diversity assessment of Achillea fragrantissima (Forskal) Schultz Bip. populations collected from different ecosystems in Jordan. Res. Crop. 2009, 10, 632–642. [Google Scholar]
- Morsy, A.A. Molecular variations of Achillea fragrantissima (Forssk.) SCH. BIP. growing in five areas of South Sinai. Int. J. Agric. Biol. 2007, 9, 11–17. [Google Scholar]
- Badr, A.; El-Shazly, H.; Ahmed, H.; Hamouda, M.; El-Khateeb, E.; Sakr, M. Genetic diversity of Achillea fragrantissima in Egypt inferred from phenotypic variations and ISSR markers associated with traits of plant size and seed yield. Plant Genet. Resour. 2017, 15, 239–247. [Google Scholar] [CrossRef]
- Gemeinholzer, B.; Bachmann, K. Examining morphological and molecular diagnostic character states of Cichorium intybus L. (Asteraceae) and C. spinosum L. Plant Syst. Evol. 2005, 253, 105–123. [Google Scholar] [CrossRef]
- Duminil, J.; Di Michele, M. Plant species delimitation: A comparison of morphological and molecular markers. Plant Biosyst. 2009, 143, 528–542. [Google Scholar] [CrossRef]
- Pandey, S.; Kumar, S.; Mishra, U.; Rai, A.; Singh, M.; Rai, M. Genetic diversity in Indian ash gourd (Benincasa hispida) accessions as revealed by quantitative traits and RAPD markers. Sci. Hortic. 2008, 118, 80–86. [Google Scholar] [CrossRef]
- Abd El-Ghani, M.; Abo El-Kheir, M.; Abdel-Dayem, M.; Abd El-Hamid, M. Vegetation analysis and soil characteristics of five common desert climbing plants in Egypt. Turk. J. Botany 2011, 35, 561–580. [Google Scholar]
- Salama, F.; El-Ghani, M.; Gadallah, M.; El-Naggar, S.; Amro, A. Variations in Vegetation Structure, Species Dominance and Plant Communities in South of the Eastern Desert-Egypt. Not. Sci. Biol. 2014, 6, 41–58. [Google Scholar] [CrossRef] [Green Version]
- Walker, J.M.; Sunkar, R. Plant Stress Tolerance—Methods and Protocols; Humana Press: Totowa, NJ, USA, 2010; ISBN 9781607617013. [Google Scholar]
- Sayed, A.; Mohamed, A.A.; Salama, F.M. Ecophysiological studies on three desert plants growing in Wadi Natash, Eastern Desert, Egypt. J. Biol. Earth Sci. 2013, 3, 135–143. [Google Scholar]
- Salama, F.; Sayed, S.; Abd El-Gelil, A. Ecophysiological responses of Calligonum polygonoides and Artemisia judaica plants to severe desert aridity. Turk. J. Bot. 2015, 39, 253–266. [Google Scholar] [CrossRef] [Green Version]
- Shaltout, K.H.; El-Kady, H.F.; Al-Sodany, Y.M. Vegetation analysis of the Mediterranean region of Nile Delta. Vegetatio 1995, 116, 73–83. [Google Scholar]
- Thada, V.; Jaglan, V. Comparison of jaccard, dice, cosine similarity coefficient to find best fitness value for web retrieved documents using genetic algorithm. Int. J. Innov. Eng. Technol. 2013, 2, 202–205. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name Primer | Sequence | Monomorphic Bands | Polymorphic Bands | Unique Bands | Total Bands | Polymorphism (%) |
|---|---|---|---|---|---|---|
| ISSR- 1 | 5′-AGAGAGAGAGAGAGAGC-3′ | 0 | 9 | 5 | 14 | 100 |
| ISSR- 4 | 5′-ACACACACACACACACG-3′ | 4 | 14 | 2 | 20 | 80 |
| ISSR- 5 | 5′-GTGTGTGTGTGTGTGTYG-3′ | 0 | 13 | 0 | 13 | 100 |
| ISSR- 10 | 5′-GACAGACAGACAGACAAT-3′ | 1 | 6 | 1 | 8 | 88 |
| ISSR- 12 | 5′-ACACACACACACACACC-3′ | 2 | 10 | 3 | 15 | 87 |
| ISSR- 13 | 5′-AGAGAGAGAGAGAGAGT-3′ | 5 | 5 | 1 | 11 | 55 |
| ISSR- 14 | 5′-CTCCTCCTCCTCCTCTT-3′ | 1 | 5 | 1 | 7 | 86 |
| ISSR- 18 | 5′-HVHCACACACACACACAT-3′ | 3 | 2 | 0 | 5 | 40 |
| ISSR- 19 | 5′-HVHTCCTCCTCCTCCTCC-3′ | 2 | 5 | 1 | 8 | 75 |
| ISSR- 20 | 5′-HVHTGTGTGTGTGTGTGT-3′ | 2 | 7 | 3 | 12 | 83 |
| Total | 20 | 76 | 17 | 113 | 79.4 |
| No | Character | Populations | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | ||
| 1 | Plant height (cm) | 110 * ± 5.0 | 100 * ± 3.0 | 44.3 ± 3.0 | 68 * ± 2.0 | 65 * ± 2.0 | 80 * ± 3.0 | 120 * ± 5.0 | 65 * ± 4.0 | 120 * ± 6.0 | 150 * ± 5.0 |
| 2 | Plant diameter (cm) | 200 * ± 6.0 | 170 * ± 5.0 | 67.7 ± 4.0 | 240 * ± 2.0 | 210 * ± 3.0 | 280 * ± 6.0 | 109 * ± 3.0 | 115 * ± 1.0 | 230 * ± 4.0 | 115 * ± 2.0 |
| 3 | Leaf length (cm) | 2.06 * ± 0.1 | 1.8 * ± 0.1 | 2.09 ± 0.1 | 1.39 * ± 0.1 | 1.71 * ± 0.1 | 1.95 * ± 0.1 | 1.65 * ± 0.1 | 2.82 * ± 0.1 | 1.63 * ± 0.1 | 2.78 * ± 0.1 |
| 4 | Leaf width (mm) | 6.8 * ± 0.5 | 5.6 * ± 0.4 | 5.6 ± 0.3 | 6.4 * ± 0.4 | 5.0 * ± 0.4 | 5.6 * ± 0.2 | 5.4 * ± 0.1 | 7.2 * ± 0.4 | 4.4 * ± 0.5 | 6.8 * ± 0.5 |
| 5 | Leaf index | 3.03 * ± 0.08 | 3.21 * ± 0.41 | 3.72 ± 0.02 | 2.17 * ± 0.02 | 3.43 * ± 0.07 | 3.48 * ± 0.3 | 3.06 * ± 0.13 | 3.91 * ± 0.08 | 3.71 * ± 0.66 | 4.09 * ± 0.15 |
| 6 | Inflorescence length (cm) | 2.81 * ± 0.2 | 3.01 * ± 0.3 | 2.40 ± 0.1 | 4.57 * ± 0.3 | 3.60 * ± 0.4 | 3.99 * ± 0.1 | 1.69 * ± 0.2 | 3.63 * ± 0.2 | 3.30 * ± 0.4 | 2.93 * ± 0.2 |
| No. | Gravel % | Coarse Sand % | Fine Sand % | Silt % | Clay % | Moisture (%) | pH | TDS (mg/L) | EC (µs/cm) | Na (mg/g) | K (mg/g) | Ca (mg/g) | Mg (mg/g) | Cl (mg/g) | PO4 (mg/g) |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | 27.91 ± 4.5 | 9.7 ± 2.08 | 58.0 ± 3.4 | 2.3 ± 0.3 | 2.1 ± 0.11 | 0.08 ± 0.0 | 7.9 ± 0.24 | 116.6 ± 4.4 | 188.9 ± 86.2 | 2.9 ± 2.1 | 1.5 ± 0.43 | 0.49 ± 0.4 | 0.17 ± 0.07 | 0.15 ± 0.04 | 0.5 ± 0.4 |
| 2 | 19.31 ± 6.4 | 19.9 ± 7.5 | 49.1 ± 5.4 | 6.2 ± 8.2 | 5.4 ± 6.9 | 0.11 ± 0.01 | 8.6 ± 0.08 | 85.56 ± 7.2 | 145.9 ± 17.8 | 2.5 ± 0.74 | 2.5 ± 0.47 | 0.21 ± 0.03 | 0.11 ± 0.03 | 0.10 ± 0.01 | 0.28 ± 0.1 |
| 3 | 24.81 ± 2.1 | 55.5 ± 3.3 | 10.8 ± 1.9 | 4.6 ± 1.7 | 4.4 ± 1.56 | 0.29 ± 0.0 | 7.2 ± 0.09 | 2106 ± 135 | 3380.0 ± 645 | 11.5 ± 3.2 | 2.8 ± 1.02 | 4.2 ± 0.7 | 0.25 ± 0.21 | 0.60 ± 0.30 | 0.74 ± 0.7 |
| 4 | 0.02 ± 0.01 | 18.07 ± 2.8 | 60.7 ± 4.5 | 15.8 ± 1.9 | 5.4 ± 0.21 | 0.78 ± 0.0 | 8.3 ± 0.14 | 74.4 ± 5.4 | 123.2 ± 9.95 | 0.22 ± 0.1 | 0.03 ± 0.01 | 0.23 ± 0.0 | 0.08 ± 0.01 | 1.04 ± 0.02 | 0.23 ± 0.01 |
| 5 | 2.17 ± 1.86 | 3.9 ± 3.3 | 66.7 ± 22.9 | 7.5 ± 4.8 | 19.7 ± 22 | 0.23 ± 0.2 | 8.4 ± 0.48 | 139.6 ± 67.4 | 232.5 ± 113.2 | 2.6 ± 1.4 | 2.5 ± 0.93 | 0.34 ± 0.0 | 0.08 ± 0.03 | 0.09 ± 0.01 | 0.21 ± 0.02 |
| 6 | 30.27 ± 3.6 | 21.3 ± 2.6 | 44.0 ± 4.0 | 2.0 ± 0.4 | 2.4 ± 1.01 | 0.04 ± 0.0 | 8.0 ± 0.19 | 76.6 ± 19.9 | 126.8 ± 31.9 | 2.3 ± 0.93 | 1.5 ± 0.32 | 0.16 ± 0.0 | 0.12 ± 0.05 | 0.08 ± 0.02 | 0.34 ± 0.2 |
| 7 | 22.70 ± 18.8 | 9.8 ± 5.6 | 45.1 ± 8.3 | 10.1 ± 6.9 | 12.2 ± 8.9 | 0.50 ± 0.1 | 7.9 ± 0.16 | 281.7 ± 20.1 | 469.0 ± 335.6 | 4.8 ± 1.35 | 5.4 ± 0.75 | 0.28 ± 0.0 | 0.06 ± 0.03 | 0.07 ± 0.02 | 0.23 ± 0.03 |
| 8 | 15.84 ± 3.08 | 20.5 ± 3.9 | 58.2 ± 7.9 | 3.4 ± 2.7 | 2.1 ± 0.10 | 0.13 ± 0.0 | 8.5 ± 0.12 | 112.3 ± 27.6 | 186.8 ± 45.6 | 1.8 ± 0.60 | 2.5 ± 0.77 | 0.44 ± 0.2 | 0.05 ± 0.02 | 0.21 ± 0.04 | 0.3 ± 0.06 |
| 9 | 3.08 ± 4.52 | 3.6 ± 4.6 | 29.5 ± 2.4 | 36.0 ± 0.8 | 27.8 ± 6.8 | 0.61 ± 0.3 | 8.1 ± 0.12 | 131.2 ± 18.3 | 216.9 ± 24.1 | 1.5 ± 0.29 | 2.5 ± 0.20 | 0.23 ± 0.1 | 0.12 ± 0.01 | 0.16 ± 0.01 | 0.3 ± 0.10 |
| 10 | 2.1 ± 2.52 | 4.4 ± 3.6 | 30.7 ± 4.1 | 37.5 ± 1.7 | 25.3 ± 2.8 | 0.51 ± 0.3 | 8.2 ± 0.10 | 134.2 ± 15.1 | 220.1 ± 14.5 | 1.7 ± 0.25 | 2.9 ± 0.22 | 0.27 ± 0.1 | 0.12 ± 0.04 | 0.15 ± 0.01 | 0.3 ± 0.11 |
| Axes | 1 | 2 |
|---|---|---|
| Eigenvalues | 0.488 | 0.413 |
| Species–environment correlations | 0.998 | 0.990 |
| Cumulative percentage variance of species–environment relation | 25.7 | 47.4 |
| FS (%) | −0.356 * | 0.447 * |
| Silt (%) | −0.173 | −0.384 * |
| Clay (%) | −0.316 * | −0.198 |
| Moisture content | −0.412 * | −0.073 |
| Mg (mg g−1 dry soil) | 0.742 ** | 0.021 |
| Cl (mg g−1 dry soil) | 0.061 | 0.315 * |
| PO4 (mg g−1 dry soil) | 0.640 ** | −0.077 |
| Species–environment correlations | 0.926 | 0.744 |
| Cumulative percentage variance of species–environment relation: | 30.7 | 47.7 |
| Plant height (PH) | −0.0196 | −0.0906 |
| Plant diameter (PD) | 0.0391 | 0.2085 |
| Leaf width (LW) | 0.0771 | −0.1093 |
| Leaf index (LI) | −0.0543 | −0.7358 |
| Inflorescence length(IL) | −0.1317 | 0.0762 |
| 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | |
|---|---|---|---|---|---|---|---|---|---|---|
| 1 | 1 | |||||||||
| 2 | 0.953 | 1 | ||||||||
| 3 | 0.715 | 0.729 | 1 | |||||||
| 4 | 0.847 | 0.836 | 0.745 | 1 | ||||||
| 5 | 0.799 | 0.819 | 0.836 | 0.925 | 1 | |||||
| 6 | 0.848 | 0.888 | 0.831 | 0.819 | 0.81 | 1 | ||||
| 7 | 0.816 | 0.823 | 0.817 | 0.87 | 0.911 | 0.851 | 1 | |||
| 8 | 0.818 | 0.849 | 0.86 | 0.889 | 0.921 | 0.874 | 0.959 | 1 | ||
| 9 | 0.829 | 0.851 | 0.816 | 0.913 | 0.942 | 0.857 | 0.951 | 0.974 | 1 | |
| 10 | 0.815 | 0.817 | 0.792 | 0.885 | 0.918 | 0.884 | 0.932 | 0.922 | 0.943 | 1 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
El-Tayeh, N.A.; Galal, H.K.; Soliman, M.I.; Zaki, H. Association of Morphological, Ecological, and Genetic Diversity of Aerva javanica Populations Growing in the Eastern Desert of Egypt. Agronomy 2020, 10, 402. https://doi.org/10.3390/agronomy10030402
El-Tayeh NA, Galal HK, Soliman MI, Zaki H. Association of Morphological, Ecological, and Genetic Diversity of Aerva javanica Populations Growing in the Eastern Desert of Egypt. Agronomy. 2020; 10(3):402. https://doi.org/10.3390/agronomy10030402
Chicago/Turabian StyleEl-Tayeh, Noha A., Hanaa K. Galal, Magda I. Soliman, and Hoida Zaki. 2020. "Association of Morphological, Ecological, and Genetic Diversity of Aerva javanica Populations Growing in the Eastern Desert of Egypt" Agronomy 10, no. 3: 402. https://doi.org/10.3390/agronomy10030402