
Endoparasitic Gall Mites: Two New Novophytoptus Species (Eriophyoidea, Phytoptidae) from Southern African Sedges (Cyperaceae, Carex) and New Hypotheses on the Phylogeny of Novophytoptines †
 , , , and
, , , and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
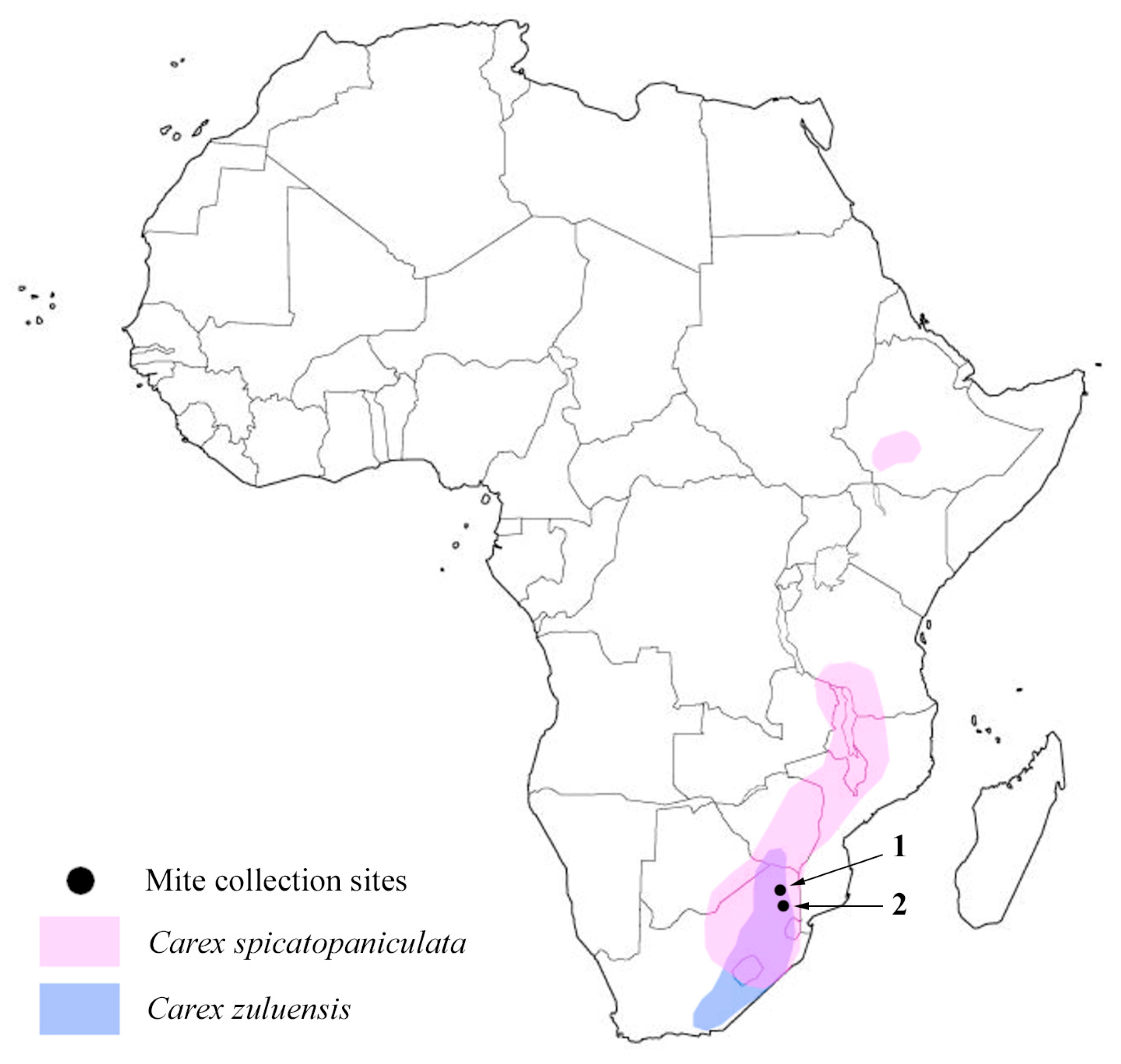
:1. Introduction
2. Material and Methods
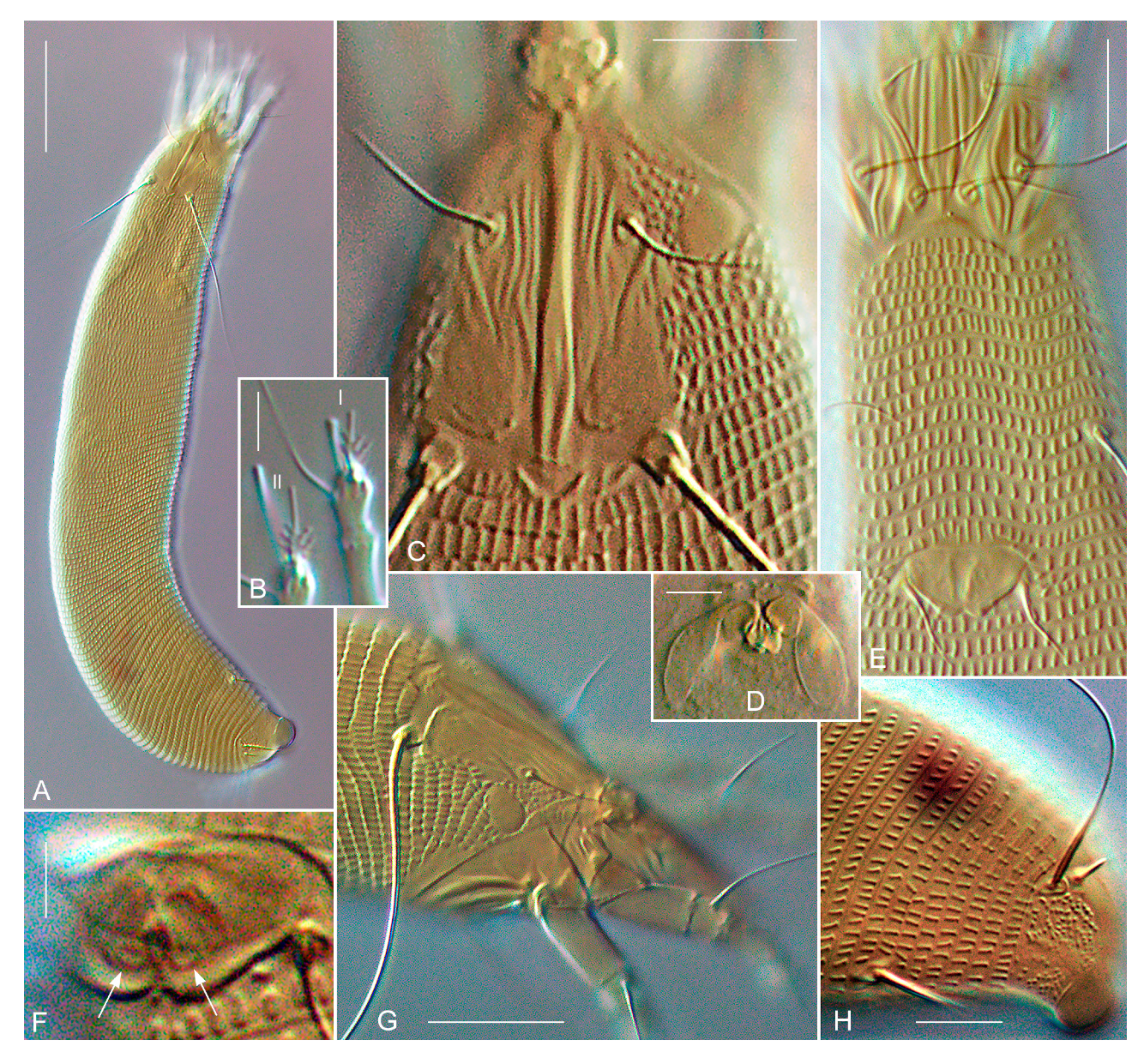
3. Taxonomy
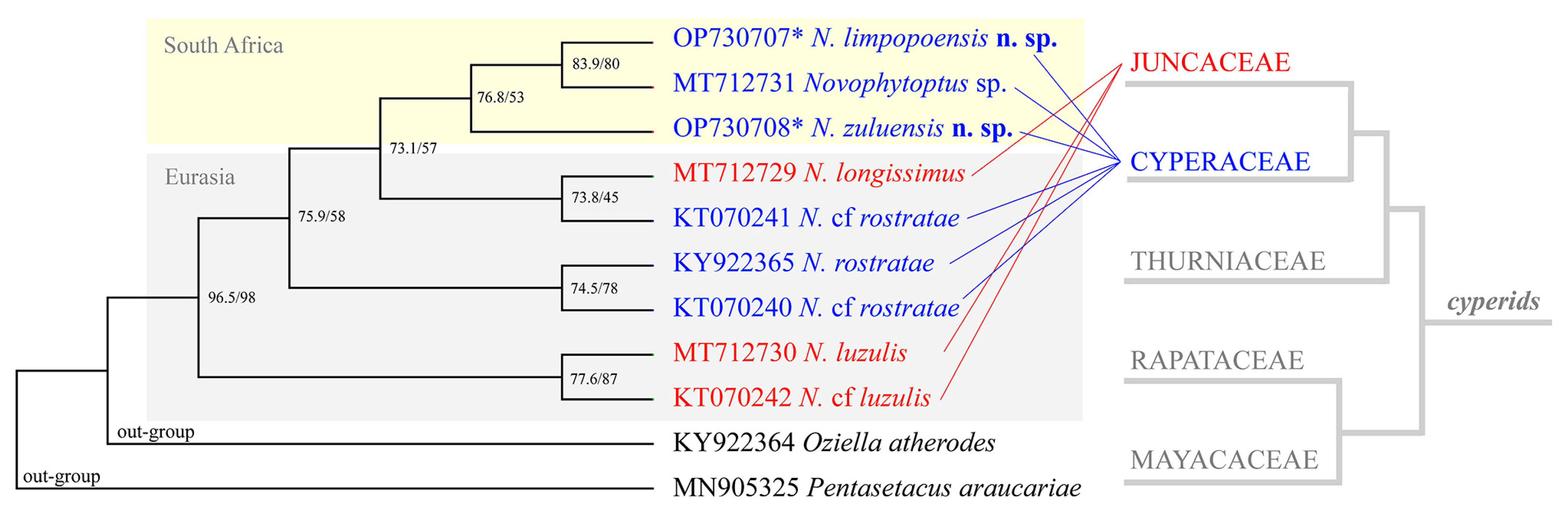
4. Molecular Phylogenetic Analyses
5. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Oldfield, G.N. Diversity and host plant specificity. In Eriophyoid Mites: Their Biology, Natural Enemies and Control. World Crop Pests 6; Lindquist, E.E., Sabelis, M.W., Bruin, J., Eds.; Elsevier Science Publishing: Amsterdam, The Netherlands, 1996; Volume 6, pp. 199–216. [Google Scholar] [CrossRef]
- Amrine, J.W., Jr.; Stasny, T.A.H.; Flechtmann, C.H.W. Revised Keys to the World Genera of the Eriophyoidea (Acari: Prostigmata); Indira Publishing House: West Bloomfield, MI, USA, 2003; 244p. [Google Scholar]
- Sullivan, G.; Ozman-Sullivan, S.K. Alarming evidence of widespread mite extinctions in the shadows of plant, insect and vertebrate extinctions. Austral Ecol. 2021, 46, 163–176. [Google Scholar] [CrossRef]
- Lindquist, E.E. External anatomy and notation of structures. In Eriophyoid Mites: Their Biology, Natural Enemies and Control. World Crop Pests 6; Lindquist, E.E., Sabelis, M.W., Bruin, J., Eds.; Elsevier Science Publishing: Amsterdam, The Netherlands, 1996; Volume 6, pp. 3–31. [Google Scholar] [CrossRef]
- Skoracka, A.; Smith, L.; Oldfield, G.; Cristofaro, M.; Amrine, J.W. Host-plant specificity and specialization in eriophyoid mites and their importance for the use of eriophyoid mites as biocontrol agents of weeds. Exp. Appl. Acarol. 2010, 51, 93–113. [Google Scholar] [CrossRef]
- Paponova, S.S.; Chetverikov, P.E.; Pautov, A.A.; Yakovleva, O.V.; Zukoff, S.N.; Vishnyakov, A.E.; Sukhareva, S.I.; Krylova, E.G.; Dodueva, I.E.; Lutova, L.A. Gall mite Fragariocoptes setiger (Eriophyoidea) changes leaf developmental program and regulates gene expression in the leaf tissues of Fragaria viridis (Rosaceae). Ann. Appl. Biol. 2018, 172, 33–46. [Google Scholar] [CrossRef]
- de Lillo, E.; Pozzebon, A.; Valenzano, D.; Duso, C. An intimate relationship between eriophyoid mites and their host plants–A review. Front. Plant Sci. 2018, 9, 1786. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Poulin, R. Evolutionary Ecology of Parasites, 2nd ed.; Princeton University Press: Princeton, NJ, USA, 2011; 360p. [Google Scholar] [CrossRef]
- Dowling, A. The evolution of parasitism and host associations in mites. In Parasite Diversity and Diversification: Evolutionary Ecology Meets Phylogenetics; Morand, S., Krasnov, B., Littlewood, D., Eds.; Cambridge University Press: Cambridge, UK, 2015; pp. 265–288. [Google Scholar] [CrossRef]
- Krantz, G.W. Habit and Habitats. In A manual of Acarology, 3rd ed.; Krantz, G.W., Walter, D.E., Eds.; Texas Tech University Press: Lubbock, TX, USA, 2009; pp. 64–82. [Google Scholar]
- Nuzzaci, G.; Alberti, G. Internal anatomy and physiology. In Eriophyoid Mites: Their Biology, Natural Enemies and Control. World Crop Pests 6; Lindquist, E.E., Sabelis, M.W., Bruin, J., Eds.; Elsevier Science Publishing: Amsterdam, The Netherlands, 1996; Volume 6, pp. 101–150. [Google Scholar] [CrossRef]
- Sidorchuk, E.A.; Schmidt, A.R.; Ragazzi, E.; Roghi, G.; Lindquist, E.E. Plant-feeding mite diversity in Triassic amber (Acari: Tetrapodili). J. Syst. Palaeontol. 2014, 13, 129–151. [Google Scholar] [CrossRef]
- Bolton, S.J.; Bauchan, G.R.; Chetverikov, P.E.; Ochoa, R.; Klompen, H. A rudimentary sheath for the smallest of “biting” chelicerae: The mouthparts of Cunliffea (Nematalycidae) and a new hypothesis on the origin of the stylet sheath of Eriophyoidea (Acariformes). Int. J. Acarol. 2018, 44, 374–381. [Google Scholar] [CrossRef]
- Bolton, S.J.; Chetverikov, P.E.; Klompen, H. Morphological support for a clade comprising two vermiform mite lineages: Eriophyoidea (Acariformes) and Nematalycidae (Acariformes). Syst. Appl. Acarol. 2017, 22, 1096–1131. [Google Scholar] [CrossRef]
- Klimov, P.B.; Chetverikov, P.E.; Dodueva, I.E.; Vishnyakov, A.E.; Bolton, S.J.; Paponova, S.S.; Lutova, L.A.; Tolstikov, A.V. Symbiotic bacteria of the gall-inducing mite Fragariocoptes setiger (Eriophyoidea) and phylogenomic resolution of the eriophyoid position among Acari. Sci. Rep. UK 2022, 12, 3811. [Google Scholar] [CrossRef]
- Haupt, J.; Coineau, Y. Ultrastructure and functional morphology of a nematalycid mite (Acari: Actinotrichida: Endostigmata: Nematalycidae): Adaptations to mesopsammal life. Acta Zool. 1999, 80, 97–112. [Google Scholar] [CrossRef]
- Chetverikov, P.E. Hidden diversity of endoparasitic eriophyoid mites: Two new Novophytoptus Roivainen, 1947 (Acari: Eriophyoidea: Phytoptidae) species from the parenchymatous tissues of rushes (Juncaceae). Zootaxa 2015, 4006, 481–505. [Google Scholar] [CrossRef] [Green Version]
- Chetverikov, P.E.; Amrine, J.; Bauchan, G.; Ochoa, R.; Sukhareva, S.I.; Vishnyakov, A.E. Supplementary description of Novophytoptus stipae Keifer 1962 (Acariformes, Eriophyoidea) with LT-SEM observation on mites from putatively conspecific populations: Cryptic speciation or polyphagy of novophytoptines on phylogenetically remote hosts? Syst. Appl. Acarol. 2017, 22, 253–270. [Google Scholar] [CrossRef]
- Bouchenak-Khelladi, Y.; Muasya, A.M.; Linder, H.P. A revised evolutionary history of Poales: Origins and diversification. Bot. J. Linn. Soc. 2014, 175, 4–16. [Google Scholar] [CrossRef] [Green Version]
- Flechtmann, C.H. Eriophyid mites (Acari: Eriophyidae) from Brazilian sedges (Cyperaceae). Int. J. Acarol. 2004, 30, 157–164. [Google Scholar] [CrossRef]
- Amrine, J.W., Jr.; Manson, D.C.M. Preparation, mounting and descriptive study of eriophyoid mites. In Eriophyoid Mites: Their Biology, Natural Enemies and Control. World Crop Pests 6; Lindquist, E.E., Sabelis, M.W., Bruin, J., Eds.; Elsevier Science Publishing: Amsterdam, The Netherlands, 1996; Volume 6, pp. 383–396. [Google Scholar] [CrossRef]
- Chetverikov, P.E. Video projector: A digital replacement for camera lucida for drawing mites and other microscopic objects. Syst. Appl. Acarol. 2016, 21, 1278–1280. [Google Scholar] [CrossRef]
- Gehrke, B. Synopsis of Carex (Cyperaceae) from sub-Saharan Africa and Madagascar. Bot. J. Linn. Soc. 2011, 166, 51–99. [Google Scholar] [CrossRef]
- Klimov, P.B.; OConnor, B.M.; Chetverikov, P.E.; Bolton, S.J.; Pepato, A.R.; Mortazavi, A.L.; Tolstikov, A.V.; Bauchan, G.R.; Ochoa, R. Comprehensive phylogeny of acariform mites (Acariformes) provides insights on the origin of the four-legged mites (Eriophyoidea), a long branch. Mol. Phylogenet. Evol. 2018, 119, 105–117. [Google Scholar] [CrossRef]
- Katoh, K.; Misawa, K.; Kuma, K.; Miyata, T. MAFFT: A novel method for rapid multiple sequence alignment based on fast Fourier transformation. Nucleic Acids Res. 2002, 30, 3059–3066. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Katoh, K.; Rozewicki, J.; Yamada, K.D. MAFFT online service: Multiple sequence alignment, interactive sequence choice and visualization. Brief. Bioinform. 2017, 20, 1160–1166. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2017, 33, 1870–1874. [Google Scholar] [CrossRef] [Green Version]
- Minh, B.Q.; Schmidt, H.A.; Chernomor, O.; Schrempf, D.; Woodhams, M.D.; von Haeseler, A.; Lanfear, R. IQ-TREE 2: New Models and Efficient Methods for Phylogenetic Inference in the Genomic Era. Mol. Biol. Evol. 2020, 37, 1530–1534. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kalyaanamoorthy, S.; Minh, B.Q.; Wong, T.K.F.; von Haeseler, A.; Jermiin, L.S. ModelFinder: Fast model selection for accurate phylogenetic estimates. Nat. Methods 2017, 14, 587–589. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Skoracka, A.; Boczek, J. Novophytoptus glyceriae sp. n. and N. ammophilae sp. n., the new species of eriophyoid mites (Acari: Eriophyoidea) from grasses. Acarina 2000, 8, 95–101. [Google Scholar]
- Roivainen, H. Eriophyid news from Finland. Entomol. Fenn. 1947, 3, 1–51. [Google Scholar]
- Chetverikov, P.E.; Sukhareva, S.I. Supplementary descriptions and biological notes on eriophyid mites (Acari: Eriophyidae) of the genus Novophytoptus Roivainen, 1947. Acarina 2007, 15, 261–268. [Google Scholar]
- Petanović, R.; Chetverikov, P.; Smiljanić, D. Eriophyoid mites (Acari: Eriophyoidea) from sedges (Carex spp., Cyperaceae) in Serbia. Acta Entomol. Serbica 2007, 12, 107–113. [Google Scholar]
- Skoracka, A.; Dabert, M. The cereal rust mite Abacarus hystrix (Acari: Eriophyoidea) is a complex of species: Evidence from mitochondrial and nuclear DNA sequences. Bull. Entomol. Res. 2010, 100, 263–272. [Google Scholar] [CrossRef] [PubMed]
- Skoracka, A.; Kuczyński, L.; de Mendonça, R.S.; Dabert, M.; Szydło, W.; Knihinicki, D.; Truol, G.; Navia, D. Cryptic species within the wheat curl mite Aceria tosichella (Keifer)(Acari: Eriophyoidea), revealed by mitochondrial, nuclear and morphometric data. Invertebr. Syst. 2012, 26, 417–433. [Google Scholar] [CrossRef] [Green Version]
- Smith, I.M. A new species of eriophyoid mite with eye-like structures, and remarks on the genus Phytoptus (Acari: Prostigmata: Phytoptidae). Can. Entomol. 1977, 109, 1097–1102. [Google Scholar] [CrossRef]
- Flechtmann, C.H.; Arana, M.; Ciarrocchi, F.; Chetverikov, P.E.; Amrine, J.W., Jr. Rediscovery and redescription of two eriophyid mites (Acari, Prostigmata, Eriophyidae) from Baccharis salicifolia (Asteraceae), from Argentina with remarks on the eriophyoid coverflap base. Acarologia 2015, 55, 387–396. [Google Scholar] [CrossRef]
- Lebrun, J.-P.; Stork, A.L. Tropical African Flowering Plants: Ecology and Distribution; Cyperaceae; Éditions des Conservatoire et Jardin Botaniques: Genève, Switzerland, 2020; Volume 11, 368p. [Google Scholar]
- Herman, P.P.J.; Retief, E. Plants of thenorthern provinces of South Africa: Keys and diagnostic characters. Strelitzia 1997, 6, 1–681. [Google Scholar]
- Retief, E.; Meyer, N.L. Cyperaceae. In Plants of the Free State: Inventory and Identification Guide; Retief, E., Meyer, N.L., Eds.; Strelitzia, South African National Biodiversity Institute: Pretoria, South Africa, 2017; Volume 38, pp. 875–904. [Google Scholar]
- Global Plants. Available online: http://plants.jstor.org (accessed on 22 November 2022).
- Chetverikov, P.E.; Craemer, C.; Cvrković, T.; Klimov, P.B.; Petanović, R.U.; Romanovich, A.E.; Sukhareva, S.I.; Zukoff, S.N.; Bolton, S.; Amrine, J. Molecular phylogeny of the phytoparasitic mite family Phytoptidae (Acariformes: Eriophyoidea) identified the female genitalic anatomy as a major macroevolutionary factor and revealed multiple origins of gall induction. Exp. Appl. Acarol. 2021, 83, 31–68. [Google Scholar] [CrossRef]
- Lindquist, E.E.; Amrine, J.W., Jr. Systematics, diagnoses for major taxa, and keys to families and genera with species on plants of economic importance. In Eriophyoid Mites: Their Biology, Natural Enemies and Control. World Crop Pests 6; Lindquist, E.E., Sabelis, M.W., Bruin, J., Eds.; Elsevier Science Publishing: Amsterdam, The Netherlands, 1996; Volume 6, pp. 33–87. [Google Scholar] [CrossRef]
- Guo, J.F.; Li, H.S.; Wang, B.; Xue, X.F.; Hong, X.Y. DNA barcoding reveals the protogyne and deutogyne of Tegolophus celtis sp. nov. (Acari: Eriophyidae). Exp. Appl. Acarol. 2015, 67, 393–410. [Google Scholar] [CrossRef]
- Petanović, R. Towards an integrative approach to taxonomy of Eriophyoidea (Acari, Prostigmata)-An overview. Ecol. Montenegrina 2016, 7, 580–599. [Google Scholar] [CrossRef]
- Druciarek, T.; Lewandowski, M.; Tzanetakis, I. A new, sensitive and efficient method for taxonomic placement in the Eriophyoidea and virus detection in individual eriophyoids. Exp. Appl. Acarol. 2019, 78, 247–261. [Google Scholar] [CrossRef]
- Chetverikov, P.E.; Klimov, P.B.; He, Q. Vertical transmission and seasonal dimorphism of eriophyoid mites (Acariformes, Eriophyoidea) parasitic on the Norway maple: A case study. Royal Soc. Open Sci. 2022, 9, 220820. [Google Scholar] [CrossRef]
- Skoracka, A.; Magalhaes, S.; Rector, B.G.; Kuczyński, L. Cryptic speciation in the Acari: A function of species lifestyles or our ability to separate species? Exp. Appl. Acarol. 2015, 67, 165–182. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Druciarek, T.; Lewandowski, M.; Tzanetakis, I. Molecular phylogeny of Phyllocoptes associated with roses discloses the presence of a new species. Infect. Genet. Evol. 2021, 95, 105051. [Google Scholar] [CrossRef] [PubMed]
- Cvrković, T.; Chetverikov, P.; Vidović, B.; Petanović, R. Cryptic speciation within Phytoptus avellanae s.l. (Eriophyoidea: Phytoptidae) revealed by molecular data and observations on molting Tegonotus-like nymphs. Exp. Appl. Acarol. 2016, 68, 83–96. [Google Scholar] [CrossRef]
- Saccaggi, D.L.; Maboeei, P.; Powell, C.; Ngubane-Ndhlovu, N.P.; Allsopp, E.; Wesley-Smith, J.; van Asch, B. Towards Solving the “Colomerus vitis Conundrum”: Genetic Evidence Reveals a Complex of Highly Diverged Groups with Little Morphological Differentiation. Diversity 2022, 14, 342. [Google Scholar] [CrossRef]
- Larose, C.; Schwander, T. Nematode endoparasites do not codiversify with their stick insect hosts. Ecol. Evol. 2016, 6, 5446–5458. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sabelis, M.W.; Bruin, J. Evolutionary ecology: Life history patterns, food plant choice and dispersal. In Eriophyoid Mites: Their Biology, Natural Enemies and Control. World Crop Pests 6; Lindquist, E.E., Sabelis, M.W., Bruin, J., Eds.; Elsevier Science Publishing: Amsterdam, The Netherlands, 1996; Volume 6, pp. 329–366. [Google Scholar]









Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chetverikov, P.E.; Craemer, C.; Gankevich, V.D.; Kremenetskaya, M.V.; Kuzmin, I.V.; Zhuk, A.S. Endoparasitic Gall Mites: Two New Novophytoptus Species (Eriophyoidea, Phytoptidae) from Southern African Sedges (Cyperaceae, Carex) and New Hypotheses on the Phylogeny of Novophytoptines. Diversity 2023, 15, 416. https://doi.org/10.3390/d15030416
Chetverikov PE, Craemer C, Gankevich VD, Kremenetskaya MV, Kuzmin IV, Zhuk AS. Endoparasitic Gall Mites: Two New Novophytoptus Species (Eriophyoidea, Phytoptidae) from Southern African Sedges (Cyperaceae, Carex) and New Hypotheses on the Phylogeny of Novophytoptines. Diversity. 2023; 15(3):416. https://doi.org/10.3390/d15030416
Chicago/Turabian StyleChetverikov, Philipp E., Charnie Craemer, Vladimir D. Gankevich, Maria V. Kremenetskaya, Igor V. Kuzmin, and Anna S. Zhuk. 2023. "Endoparasitic Gall Mites: Two New Novophytoptus Species (Eriophyoidea, Phytoptidae) from Southern African Sedges (Cyperaceae, Carex) and New Hypotheses on the Phylogeny of Novophytoptines" Diversity 15, no. 3: 416. https://doi.org/10.3390/d15030416