
Notes to the Taxonomic Affiliation of the Bulbophyllym Sect. Physometra (Orchidaceae, Epidendroideae) Based on Molecular Phylogenetic Analyses

Abstract
:1. Introduction
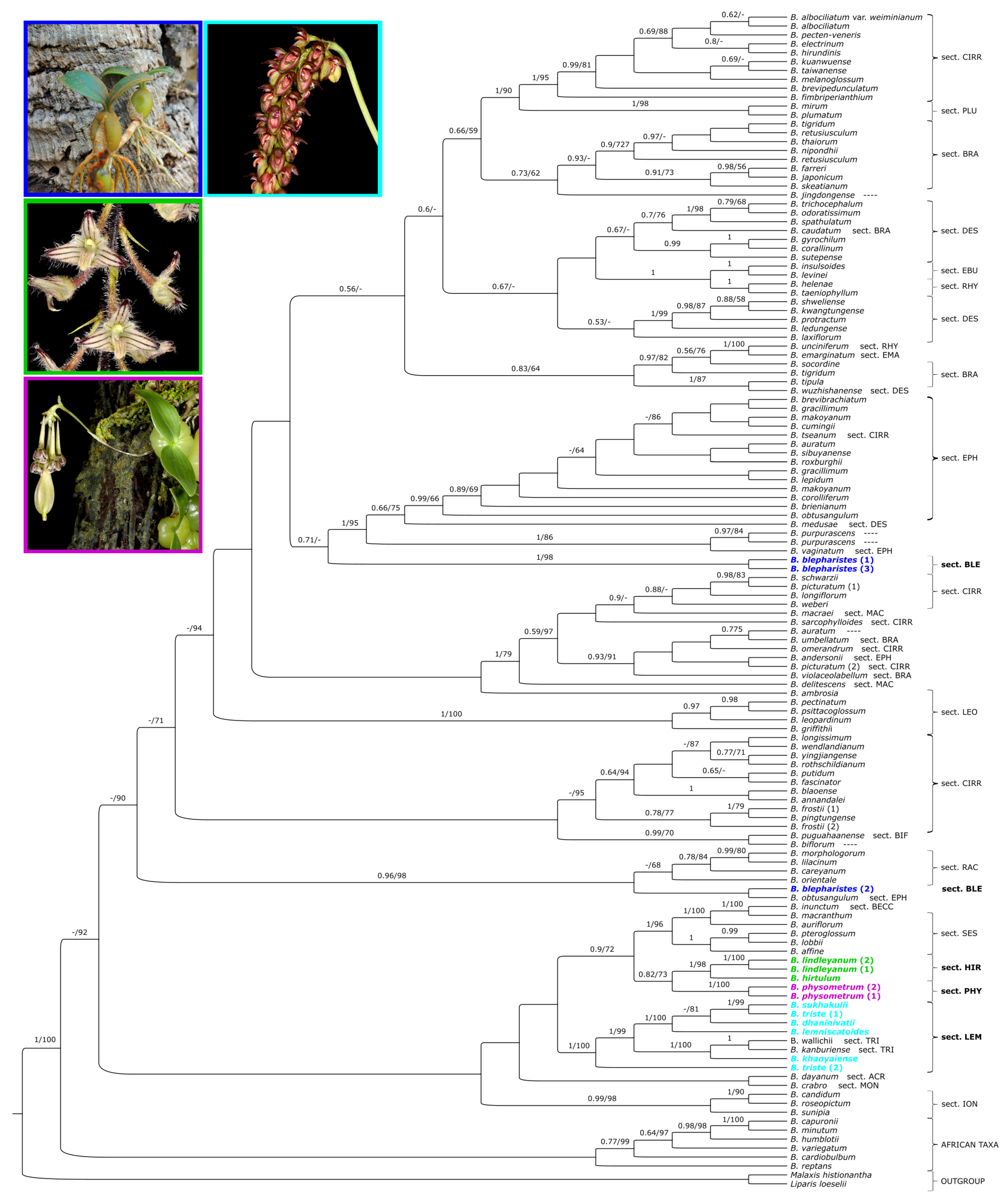
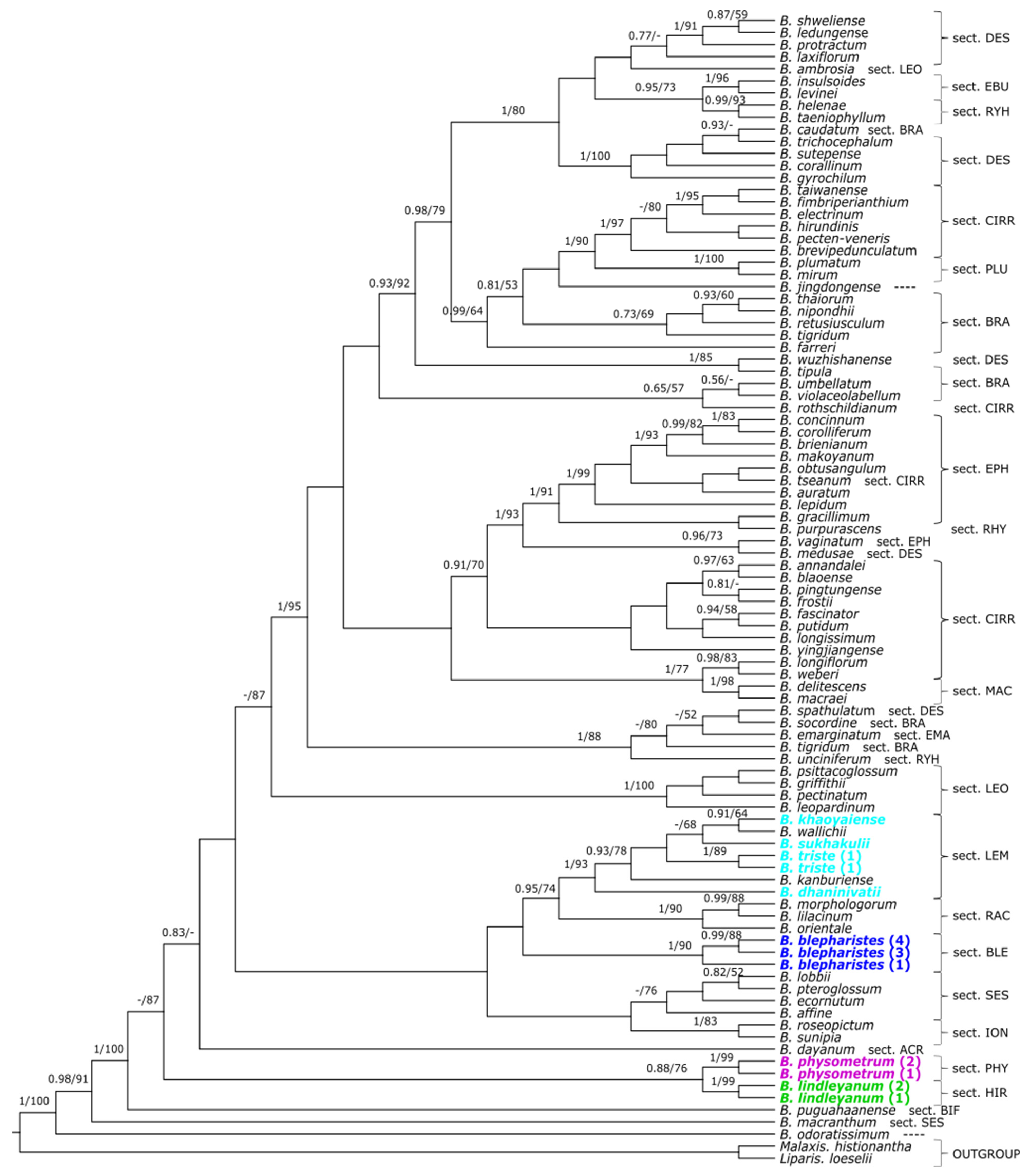
2. Results
3. Discussion
3.1. Phylogeny of Bulbophyllum
3.2. Relationship with the Section Hirtula
3.3. Flowers Dimorphism
3.4. Two-Leaved Pseudobulb
3.5. Mimicry of Flower or Fruit?
4. Materials and Methods
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- POWO. Plants of the World Online. Facilitated by the Royal Botanic Gardens, Kew. Published on the Internet. Available online: http://www.plantsoftheworldonline.org/ (accessed on 16 January 2023).
- Sieder, A.; Rainer, H.; Kiehn, M. CITES Checklist for Bulbophyllum and Allied Taxa (Orchidaceae); Botanical Garden of the University of Vienna Press: Vienna, Austria, 2007; pp. 1–319. [Google Scholar]
- Pridgeon, A.M.; Cribb, P.J.; Chase, M.W.; Rasmussen, F.N. Genera Orchidacearum Vol. 6 Epidendroideae; Oxford University Press: New York, NY, USA, 2014; pp. 1–687. [Google Scholar]
- Vermeulen, J.J. Orchids of Borneo: Bulbophyllum; Royal Botanic Gardens Kew: London, UK, 1991; Volume 3. [Google Scholar]
- Dressler, R.L. Phylogeny and Classification of the Orchid Family; Dioscorides Press: Portland, OR, USA, 1993; pp. 1–311. [Google Scholar]
- Smidt, E.C.; Borba, E.L.; Gravendeel, B.; Fischer, G.A.; van den Berg, C. Molecular phylogeny of the Neotropical sections of Bulbophyllum (Orchidaceae) using nuclear and plastid spacers. Taxon 2011, 60, 1050–1064. [Google Scholar] [CrossRef]
- Fischer, G.A.; Gravendeel, B.; Sieder, A.; Andriantiana, J.; Heiselmayer, P.; Cribb, P.J.; Kiehn, M. Evolution of resupination in Malagasy species of Bulbophyllum (Orchidaceae). Mol. Phylogenetics Evol. 2007, 45, 358–376. [Google Scholar] [CrossRef]
- Hu, A.-Q.; Gale, S.W.; Liu, Z.-J.; Suddee, S.; Hsu, T.-C.; Fischer, G.A.; Saunders, R.M.K. Molecular phylogenetics and floral evolution of the Cirrhopetalum alliance (Bulbophyllum, Orchidaceae): Evolutionary transitions and phylogenetic signal variation. Mol. Phylogenetics Evol. 2020, 143, 106689. [Google Scholar] [CrossRef]
- Vermeulen, J.J.; Suksathan, P.; Watthana, S. A new species and new section in Bulbophyllum (Orchidaceae; Epodendroideae; Malaxideae). Phytotaxa 2017, 302, 147–180. [Google Scholar] [CrossRef]
- Vermeulen, J.J.; O’Byrne, P.; Lamb, A. Bulbophyllum of Borneo; National History Publications: Kota Kinabalu, Malaysia, 2015; pp. 1–736. [Google Scholar]
- Hosseini, S.; Dadkhah, K.; Go, R. Molecular systematics of genus Bulbophyllum (Orchidaceae) in Peninsular Malaysia based on combined nuclear and plastid DNA sequences. Biochem. Syst. Ecol. 2016, 65, 40–48. [Google Scholar] [CrossRef]
- Gamisch, A.; Comes, H.P. Clade-age-dependent diversification under high species turnover shapes species richness disparities among tropical rainforest lineages of Bulbophyllum (Orchidaceae). BMC Evol. Biol. 2019, 19, 1–16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zavala-Páez, M.; Vieira, L.D.N.; Baura, V.A.D.; Balsanelli, E.; Souza, E.M.D.; Cevallos, M.C.; Smidt, E.D.C. Comparative plastid genomics of neotropical Bulbophyllum (Orchidaceae; Epidendroideae). Front. Plant Sci. 2020, 11, 799. [Google Scholar] [CrossRef] [PubMed]
- Tang, H.; Tang, L.; Shao, S.; Peng, Y.; Li, L.; Luo, Y. Chloroplast genomic diversity in Bulbophyllum section Macrocaulia (Orchidaceae, Epidendroideae, Malaxideae): Insights into species divergence and adaptive evolution. Plant Divers. 2021, 43, 350–361. [Google Scholar] [CrossRef]
- Geber, M.A.; Dawson, T.E.; Delph, L.F. Gender and Sexual Dimorphism in Flowering Plants; Springer Verlag: Berlin/Heidelberg, Germany, 1999. [Google Scholar]
- Harder, L.D.; Barrett, S.C.H. Ecology and Evolution of Flowers; Oxford University Press: New York, NY, USA, 2006. [Google Scholar]
- Ashman, T.L. Floral scent in whole-plant context: Sniffing out patterns of sexual dimorphism in floral scent. Funct. Ecol. 2009, 23, 852–862. [Google Scholar] [CrossRef]
- Barrett Spencer, C.H.; Hough, J. Sexual dimorphism in flowering plants. J. Exp. Bot. 2013, 64, 67–82. [Google Scholar] [CrossRef] [Green Version]
- Bell, G. On the function of flowers. Proc. R. Soc. Lond. B 1985, 224, 223–265. [Google Scholar] [CrossRef]
- Tsuji, K.; Kobayashi, K.; Hasegawa, E.; Yoshimura, J. Dimorphic flowers modify the visitation order of pollinators from male to female flowers. Sci. Rep. 2020, 10, 9965. [Google Scholar] [CrossRef]
- Romero, G.A.; Nelson, C.E. Sexual dimorphism in Catasetum orchids: Forcible pollen emplacement and male flower competition. Science 1986, 232, 1538–1540. [Google Scholar] [CrossRef] [PubMed]
- Brandt, K.; Machado, I.C.; Navarro, D.M.D.A.F.; Dötterl, S.; Ayasse, M.; Milet-Pinheiro, P. Sexual dimorphism in floral scents of the neotropical orchid Catasetum arietinum and its possible ecological and evolutionary significance. AoB Plants 2020, 12, plaa030. [Google Scholar] [CrossRef]
- Delph, L.F. Flower size dimorphism in plants with unisexual flowers. Flor. Biol. Stud. Flor. Evol. Anim.-Pollinated Plants 1996, 217–237. [Google Scholar] [CrossRef]
- Humeau, L.; Pailler, T.; Thompson, J.D. Flower size dimorphism in diclinous plants native to La Réunion Island. Plant Syst. Evol. 2003, 240, 163–173. [Google Scholar] [CrossRef]
- Wyatt, R. Inflorescence architecture: How flower number, arrangement, and phenology affect pollination and fruit-set. Am. J. Bot. 1982, 69, 585–594. [Google Scholar] [CrossRef]
- Ishii, H.S.; Hirabayashi, Y.; Kudo, G. Combined effects of inflorescence architecture, display size, plant density and empty flowers on bumble bee behaviour: Experimental study with artificial inflorescences. Oecologia 2008, 156, 341–350. [Google Scholar] [CrossRef]
- Bateman, R.M.; Rudall, P.J. Evolutionary and morphometric implications of morphological variation among flowers within an inflorescence: A case-study using European orchids. Ann. Bot. 2006, 98, 975–993. [Google Scholar] [CrossRef] [Green Version]
- Bröderbauer, D.; Diaz, A.; Weber, A. Reconstructing the origin and elaboration of insect-trapping inflorescences in the Araceae. Am. J. Bot. 2012, 99, 1666–1679. [Google Scholar] [CrossRef] [Green Version]
- Chartier, M.; Gibernau, M.; Renner, S.S. The evolution of pollinator–plant interaction types in the Araceae. Evolution 2014, 68, 1533–1543. [Google Scholar] [CrossRef]
- Jin, B.; Wang, L.; Wang, J.; Teng, N.J.; He, X.D.; Mu, X.J.; Wang, Y.L. The structure and roles of sterile flowers in Viburnum macrocephalum f. keteleeri (Adoxaceae). Plant Biol. 2010, 12, 853–862. [Google Scholar] [CrossRef] [PubMed]
- Morales, C.L.; Traveset, A.; Harder, L.D. Sterile flowers increase pollinator attraction and promote female success in the Mediterranean herb Leopoldia comosa. Ann. Bot. 2013, 111, 103–111. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, B.; Sinnott-Armstrong, M.; Schlutius, C.; Zuluaga, J.C.P.; Spriggs, E.L.; Simpson, R.G.; Donoghue, M.J. Sterile marginal flowers increase visitation and fruit set in the hobblebush (Viburnum lantanoides, Adoxaceae) at multiple spatial scales. Ann. Bot. 2019, 123, 381–390. [Google Scholar] [CrossRef] [Green Version]
- Szlachetko, D.L.; Kolanowska, M. Reconsideration of Heteranthocidium (Oncidiinae, Orchidaceae): New species and taxonomic transfers. Plant Syst. Evol. 2015, 301, 1793–1805. [Google Scholar] [CrossRef] [Green Version]
- Vermeulen, J.J. A taxonomic revision of Bulbophyllum (Orchidaceae) 2. Sections Altisceptrum and Hirtula. Gard. Bull. Singap. 2002, 54, 1–151. [Google Scholar]
- Vermeulen, J.J. Orchid Monographs: A Taxonomic Revision of the Continental African Bulbophyllinae; Brill Archive: Leiden, The Netherlands, 1987; Volume 2. [Google Scholar]
- Smidt, E.; Silva-Pereira, V.; Borba, E.; van den Berg, C. Richness, distribution and important areas to preserve Bulbophyllum in the Neotropics. Lankesteriana 2007, 7, 107–113. [Google Scholar] [CrossRef] [Green Version]
- Jersakova, J.; Spaethe, J.; Strweinzer, M.; Neumayer, J.; Paulus, H.; Dotterl, S.; Johnson, S.D. Does Traunsteinera globose (the globe orchid) dupe its pollinators through generalized food deception or mimicry? Bot. J. Linn. Soc. 2016, 180, 269–294. [Google Scholar] [CrossRef] [Green Version]
- Gigord, L.D.B.; Macnair, M.R.; Stritesky, M.; Smithson, A. The potential for floral mimicry in rewardless orchid: An experimental study. R. Soc. 2002, 269, 1389–1395. [Google Scholar] [CrossRef] [PubMed]
- Johnson, S.D. Batesian mimicry in the non-rewarding orchid Disa pulchra, and its consequences for pollinator behaviour. Biol. J. Linn. Soc. 2000, 71, 119–132. [Google Scholar] [CrossRef]
- Goodrich, K.R.; Jürgens, A. Pollination systems involving floral mimicry of fruit: Aspects of their ecology and evolution. New Phytol. 2018, 217, 74–81. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karremans, A.P.; Bogarín, D.; Otárola, M.F.; Sharma, J.; Watteyn, C.; Warner, J.; Whitworth, A. First evidence for multimodal animal seed dispersal in orchids. Curr. Biol. 2022, 33, 364–371. [Google Scholar] [CrossRef]
- Cameron, K.M. Plant ecology: Vanilla lures both insects and mammals to disperse its seeds and fruits. Curr. Biol. 2023, 33, R63–R65. [Google Scholar] [CrossRef]
- Sut, S.; Maggi, F.; Dall’Acqua, S. Bioactive secondary metabolites from orchids (Orchidaceae). Chem. Biodivers. 2017, 14, e1700172. [Google Scholar] [CrossRef]
- Nelson, A.S.; Whitehead, S.R. Fruit secondary metabolites shape seed dispersal effectiveness. Trends Ecol. Evol. 2021, 36, 1113–1123. [Google Scholar] [CrossRef] [PubMed]
- Nishida, R. Chemical ecology of insect–plant interactions: Ecological significance of plant secondary metabolites. Biosci. Biotechnol. Biochem. 2014, 78, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Katte, T.; Hong Tan, K.; Su, Z.-H.; Ono, H.; Nishida, R. Floral fragrances in two closely related fruit fly orchid, Bulbophyllum hortorum and B. macranthoides (Orchidaceae): Assortments of phenylbutanoids to attract tephritid fruit fly males. Appl. Entomol. Zool. 2020, 55, 55–64. [Google Scholar] [CrossRef]
- Nakahira, M.; Ono, H.; Wee, S.L.; Tan, K.H.; Nishida, R. Floral synomone diversification of Bulbophyllum sibling species (Orchidaceae) in attracting fruit fly pollinators. Biochem. Syst. Ecol. 2018, 81, 86–95. [Google Scholar] [CrossRef] [Green Version]
- Nishida, R.; Howcroft, N.H.; Tan, K.H.; Su, Z.-H.; Ono, H. Floral synomone components of fruit fly-attracting orchids, Bulbophyllum sinapis and B. hahlianum, in Papua New Guinea. Biochem. Syst. Ecol. 2022, 105, 401–405. [Google Scholar] [CrossRef]
- Givnish, T.J.; Spalink, D.; Ames, M.; Lyon, S.P.; Hunter, S.J.; Zuluaga, A.; Iles, W.J.D.; Clements, M.A.; Arroyo, M.T.K.; Leebens-Mack, J.; et al. Orchid phylogenetics and multiple drivers of their extraordinary diversification. Proc. Biol. Sci. 2015, 282, 20151553. [Google Scholar] [CrossRef]
- Chase, M.W.; Hills, H.H. Silica gel: An ideal material for field preservation of leaf samples for DNA studies. Taxon 1991, 40, 215–220. [Google Scholar] [CrossRef]
- Douzery, E.J.P.; Pridgeon, A.M.; Kores, P.; Linder, H.P.; Kurzweil, H.; Chase, M.W. Molecular phylogenetics of Disae (Orchidaceae): A contribution from nuclear ribosomal ITS sequences. Am. J. Bot. 1999, 86, 887–899. [Google Scholar] [CrossRef] [PubMed]
- Górniak, M.; Paun, O.; Chase, M.W. Phylogenetic relationships within Orchidaceae based on a low-copy nuclear coding gene, Xdh: Congruence with organellar and nuclear ribosomal DNA results. Mol. Phylogenet. Evol. 1999, 56, 784–795. [Google Scholar] [CrossRef]
- Molvray, M.; Kores, P.J.; Chase, M.W. Polyphyly of mycoheterotrophic orchids and functional influences on floral and molecular characters. In Monocots: Systematic and Evolution; Morrison, D.A., Wilson, K.L., Eds.; CSIRO Publishing: Collingwood, Australia, 2000; pp. 441–448. [Google Scholar]
- Cuénoud, P.; Savolainen, V.; Chatrou, L.W.; Powell, M.; Grayer, R.J.; Chase, M.W. Molecular phylogenetics of Caryophyllales based on nuclear 18S rDNA and plastid rbcL, atpB and matK DNA sequences. Am. J. Bot. 2002, 89, 132–144. [Google Scholar] [CrossRef]
- Katoh, K.; Standley, D.M. MAFTT multiple sequence alignment software version 7: Improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef] [Green Version]
- Edler, D.; Klein, J.; Antonelli, A.; Silvestro, D. RaxmlGUI 2.0: A graphical interface and toolkit for phylogenetic analyses using RAxML. Methods Ecol. Evol. 2021, 12, 373–377. [Google Scholar] [CrossRef]
- Ronquist, F.; Teslenko, M.; Pvander, M.; Huelsenbeck, J. MrBayes 3.2: Efficient Bayesian phylogenetic inference and model choice across a large model space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef] [Green Version]
- Miller, M.A.; Pfeiffer, W.; Schwartz, T. Creating the CIPRES Science Gateway for inference of large phylogenetic trees. In 2010 Gateway Computing Environments Workshop (GCE); IEEE: New Orleans, LA, USA, 2010; pp. 1–8. [Google Scholar]
- Drummond, A.J.; Rambaut, A. BEAST: Bayesian evolutionary analysis by sampling 690 trees. BMC Evol. Biol. 2007, 7, 1–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kores, P.J.; Molvray, M.; Weston, P.H.; Hopper, S.D.; Brown, A.P.; Cameron, K.M.; Chase, M.W. A phylogenetic analysis of Diurideae (Orchidaceae) based on plastid DNA sequence data. Am. J. Bot. 2001, 88, 1903–1914. [Google Scholar] [CrossRef]
- Huelsenbeck, J.P.; Ronquist, F. MrBayes: Bayesian inference of phylogeny. Bioinformatics 2001, 17, 754–755. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample Name | Place of Acquisition | Voucher Number |
|---|---|---|
| Bulbophyllum blepharistes (3) | Greenhouse of the University of Gdansk | UGDA.0000254 |
| Bulbophyllum lindleyanum (1) | Botanical Garden of Heidelberg | UGDA.0073190 |
| Bulbophyllum physometrum (1) | Greenhouse of the University of Gdansk | UGDA.0000001 |
| Bulbophyllum physometrum (2) | Private collection of Tadeusz Kusibab (Cracow) | UGDA.0073171 |
| Dataset | No. of Taxa | Informative Features | Uninformative Features | Constant Features | Total Features |
|---|---|---|---|---|---|
| nuclear combined | 132 | 529 | 232 | 686 | 1147 |
| matK gene | 95 | 155 | 109 | 873 | 1137 |
| Nuclear + plastid (ITS1-5.8S-ITS2, Xdh, matK) | 47 | 462 | 255 | 1818 | 2535 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nowak, S.; Olędrzyńska, N.; Szlachetko, D.L.; Dudek, M. Notes to the Taxonomic Affiliation of the Bulbophyllym Sect. Physometra (Orchidaceae, Epidendroideae) Based on Molecular Phylogenetic Analyses. Int. J. Mol. Sci. 2023, 24, 9709. https://doi.org/10.3390/ijms24119709
Nowak S, Olędrzyńska N, Szlachetko DL, Dudek M. Notes to the Taxonomic Affiliation of the Bulbophyllym Sect. Physometra (Orchidaceae, Epidendroideae) Based on Molecular Phylogenetic Analyses. International Journal of Molecular Sciences. 2023; 24(11):9709. https://doi.org/10.3390/ijms24119709
Chicago/Turabian StyleNowak, Sławomir, Natalia Olędrzyńska, Dariusz L. Szlachetko, and Magdalena Dudek. 2023. "Notes to the Taxonomic Affiliation of the Bulbophyllym Sect. Physometra (Orchidaceae, Epidendroideae) Based on Molecular Phylogenetic Analyses" International Journal of Molecular Sciences 24, no. 11: 9709. https://doi.org/10.3390/ijms24119709