
Pulicaria incisa (Lam.) DC. as a Potential Source of Antioxidant, Antibacterial, and Anti-Enzymatic Bioactive Molecules: Phytochemical Constituents, In Vitro and In Silico Pharmacological Analysis

 ,
,  ,
,  , , , , , , and
, , , , , , and 
Abstract
:1. Introduction
2. Results and Discussion
2.1. Chemical Analysis
2.2. Antibacterial Activity
2.3. Antibiofilm Activity
2.4. Anti-Enzymatic Activities
2.5. Total Polyphenol Content (TPC) and Total Flavonoid Content (TFC)
2.6. Antioxidant Activity
2.7. Computational Analysis and Interaction Assay
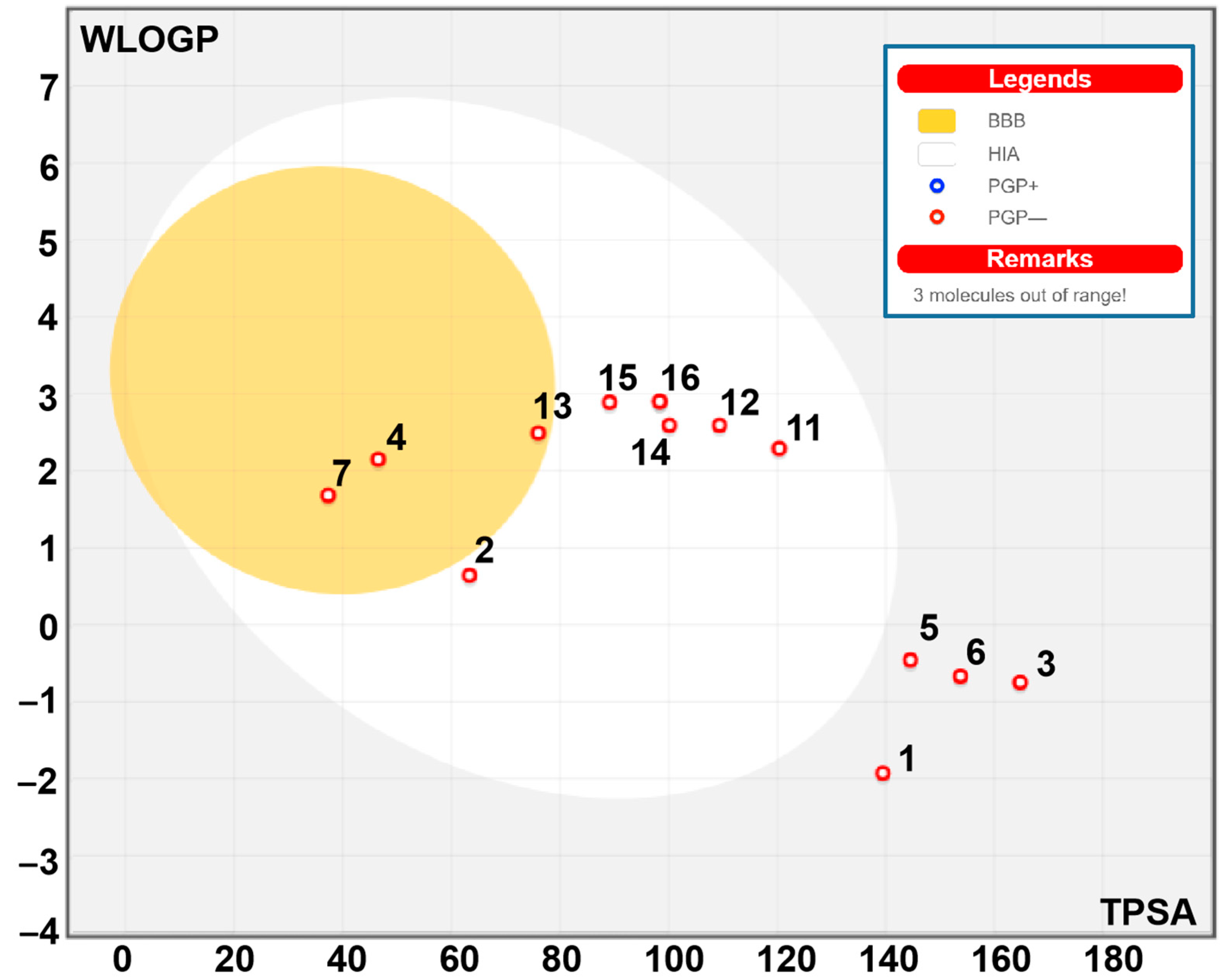
2.8. Bioavailability and Pharmacokinetics
3. Materials and Methods
3.1. Plant Identity and Extraction Methods
3.2. Chemical Analysis
3.3. Antibacterial Activity
3.3.1. Antibiofilm Experiments
3.3.2. Crystal Violet Assay
3.3.3. MTT Assay
3.4. Antioxidant Activity
3.4.1. DPPH Assay
3.4.2. FRAP Assay
3.4.3. ABTS Test
3.5. Analysis of Total Phenolic and Flavonoid Compounds
3.6. Anti-Enzymatic Activities
3.6.1. Cholinesterase Inhibition
3.6.2. α-Amylase Inhibition Assay
3.6.3. α-Glucosidase Inhibition Assay
3.7. Computational Analysis and Interaction Assay
Bioavailability and Pharmacokinetics
3.8. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Mossa, J.S.; Hifnawy, M.S.; Al-Yahya, M.A.; Al-Meshal, I.A.; Mekkawi, A.G. Aromatic Plants of Saudi Arabia-Part 8- GC/MS Analysis of Essential Oils of Pulicaria arabica and P. undulata. Int. J. Crude Drug Res. 1987, 25, 113–119. [Google Scholar] [CrossRef]
- Mohammed, H.A.; Al-Omar, M.S.; Khan, R.A.; Mohammed, S.A.A.; Qureshi, K.A.; Abbas, M.M.; Al Rugaie, O.; Abd-Elmoniem, E.; Ahmad, A.M.; Kandil, Y.I. Chemical Profile, Antioxidant, Antimicrobial, and Anticancer Activities of the Water-Ethanol Extract of Pulicaria undulata Growing in the Oasis of Central Saudi Arabian Desert. Plants 2021, 10, 1811. [Google Scholar] [CrossRef]
- Meng, X.; Zhou, J.; Sui, N. Mechanisms of Salt Tolerance in Halophytes: Current Understanding and Recent Advances. Open Life Sci. 2018, 13, 149–154. [Google Scholar] [CrossRef]
- Mohammed, H.A.; Emwas, A.-H.; Khan, R.A. Salt-Tolerant Plants, Halophytes, as Renewable Natural Resources for Cancer Prevention and Treatment: Roles of Phenolics and Flavonoids in Immunomodulation and Suppression of Oxidative Stress towards Cancer Management. Int. J. Mol. Sci. 2023, 24, 5171. [Google Scholar] [CrossRef]
- Mohamed, E.A.A.; Muddathir, A.M.; Osman, M.A. Antimicrobial activity, phytochemical screening of crude extracts, and essential oils constituents of two Pulicaria spp. growing in Sudan. Sci. Rep. 2020, 10, 17148. [Google Scholar] [CrossRef]
- Mohammed, H.A.; Khan, R.A.; Abdel-Hafez, A.A.; Abdel-Aziz, M.; Ahmed, E.; Enany, S.; Mahgoub, S.; Al-Rugaie, O.; Alsharidah, M.; Aly, M.S.A.; et al. Phytochemical Profiling, In Vitro and In Silico Anti-Microbial and Anti-Cancer Activity Evaluations and Staph GyraseB and h-TOP-IIβ Receptor-Docking Studies of Major Constituents of Zygophyllum coccineum L. Aqueous-Ethanolic Extract and Its Subsequent Fractions: An Approach to Validate Traditional Phytomedicinal Knowledge. Molecules 2021, 26, 577. [Google Scholar] [CrossRef] [PubMed]
- Elmann, A.; Telerman, A.; Mordechay, S.; Erlank, H.; Ofir, R. Antioxidant and astroprotective effects of a Pulicaria incisa infusion. Oxidative Med. Cell. Longev. 2012, 2012, 157598. [Google Scholar] [CrossRef]
- Chaib, F.; Allali, H.; Bennaceur, M.; Flamini, G. Chemical Composition and Antimicrobial Activity of Essential Oils from the Aerial Parts of Asteriscus graveolens (Forssk.) Less. and Pulicaria incisa (Lam.) DC.: Two Asteraceae Herbs Growing Wild in the Hoggar. Chem. Biodivers. 2017, 14, e1700092. [Google Scholar]
- Bakr, R.O.; Shahat, A.; Elissawy, A.E.; Ahmed, M.; Fayez, M.; Omayma, A. Eldahshan Evaluation of the hepatoprotective activity of Pulicaria incisa subspecies candolleana and in silico screening of its isolated phenolics. J. Ethnopharmacol. 2021, 271, 113767. [Google Scholar] [CrossRef] [PubMed]
- Shahat, E.A.; Bakr, R.O.; Eldahshan, O.A.; Ayoub, N.A. Chemical Composition and Biological Activities of the Essential Oil from Leaves and Flowers of Pulicaria incisa sub. candolleana (Family Asteraceae). Chem. Biodivers. 2017, 14, e1600156. [Google Scholar]
- Pecio, Ł.; Otify, A.M.; Saber, F.R.; El-Amier, Y.A.; Shalaby, M.E.; Kozachok, S.; Elmotayam, A.K.; Świątek, Ł.; Skiba, A.; Skalicka-Woźniak, K. Iphiona mucronata (Forssk.) Asch. & Schweinf. A Comprehensive Phytochemical Study via UPLC-Q-TOF-MS in the Context of the Embryo- and Cytotoxicity Profiles. Molecules 2022, 27, 7529. [Google Scholar] [CrossRef]
- Erenko, E.; Mazulin, O.V.; Smoylovska, G.P.; Grechana, O.V.; Mazulin, G.V. Aminoacid composition of species of Inula L. genus of Ukraine Flora. Farmatsevtychnyi Zhurnal 2012, 3, 94–98. [Google Scholar]
- Mohammed, H.A.; Almahmoud, S.A.; El-Ghaly, E.-S.M.; Khan, F.A.; Emwas, A.-H.; Jaremko, M.; Almulhim, F.; Khan, R.A.; Ragab, E.A. Comparative anticancer potentials of taxifolin and quercetin methylated derivatives against HCT-116 cell lines: Effects of O-methylation on taxifolin and quercetin as preliminary natural leads. ACS Omega 2022, 7, 46629–46639. [Google Scholar] [CrossRef]
- Yin, J.; Li, C.; Zhang, J.; Ding, H.; Han, L.; Yang, W.; Li, F.; Song, X.; Bie, S.; Yu, H.; et al. Comprehensive multicomponent characterization and quality assessment of Shuang-Huang-Lian powder injection using ultra-high-performance liquid chromatography-quadrupole time-of-flight-mass spectrometry and ultra-high-performance liquid chromatography-quadrupole-Orbitrap-mass spectrometry. Rapid Commun. Mass Spectrom. 2023, 37, e9479. [Google Scholar]
- Tu, Y.; Li, L.; Wang, Z.; Yang, L. Advances in analytical techniques and quality control of traditional Chinese medicine injections. J. Pharm. Anal. 2021, 206, 114353. [Google Scholar] [CrossRef] [PubMed]
- El-Naggar, S.A.; Abdel-Farid, I.B.; Elgebaly, H.A.; Germoush, M.O. Metabolomic profiling, antioxidant capacity and in vitro anticancer activity of some compositae plants growing in Saudi Arabia. Afr. J. Pharm. Pharmacol. 2015, 9, 764–774. [Google Scholar]
- Abhimannue, A.P.; Mohan, M.C. Inhibition of tumor necrosis factor-α and interleukin-1β production in lipopolysaccharide-stimulated monocytes by methanolic extract of Elephantopus scaber linn and identification of bioactive components. Appl. Biochem. Biotechnol. 2016, 179, 427–443. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.-H.; Bai, T.-C.; Shi, L.-L.; Hou, B.; Tang, R.; Zhang, R.-P.; Chen, X.-L. Antihyperlipidemic effect of Vaccinium dunalianum buds based on biological activity screening and LC-MS. J. Ethnopharmacol. 2023, 306, 116190. [Google Scholar] [CrossRef] [PubMed]
- El-Sabagh, O.A.; El-Toumy, S.A.; Mounir, R.; Farag, M.A.; Mahrous, E.A. Metabolite profiles of Pulicaria crispa and P. incisa in relation to their in-vitro/in-vivo antioxidant activity and hepatoprotective effect: A comparative mass spectrometry-based metabolomics. J. Pharm. Biomed. Anal. 2021, 194, 113804. [Google Scholar] [CrossRef]
- Al-Naqeb, G.; Rousová, J.; Kubátová, A.; Picklo Sr, M.J. Pulicaria jaubertii E. Gamal-Eldin reduces triacylglyceride content and modifies cellular antioxidant pathways in 3T3-L1 adipocytes. Chem. Biol. Interact 2016, 253, 48–59. [Google Scholar] [CrossRef]
- Askari, H.; Ghaedi, M.; Naghiha, R.; Salehi, A. In Vitro antibacterial and antifungal studies of Pulicaria undulate and Echinacea purpurea extracts in combination with nanowires (Ni: FeO (OH)) and nanoparticles (NiS). Jundishapur J. Nat. Pharm. Prod. 2020, 15, e64358. [Google Scholar] [CrossRef]
- Wang, Y.H.; Al-Rehaily, A.J.; Yousaf, M.; Ahmad, M.S.; Khan, I.A. Characterization and Discrimination of Different Pulicaria Species Using UHPLC-UV-MS QTOF (Quadrupole Time-of-Flight Mass Spectrometer). J. Chem. Soc. Pak. 2015, 37, 967–973. [Google Scholar]
- Ibraheim, Z.; Darwish, F. Further Constituents from Pulicaria Incisa. Bull. Fac. Pharm. 2002, 40, 167–173. [Google Scholar]
- Mohti, H.; Taviano, M.F.; Cacciola, F.; Dugo, P.; Mondello, L.; Marino, A.; Crisafi, G.; Benameur, Q.; Zaid, A.; Miceli, N. Inula viscosa (L.) Aiton leaves and flower buds: Effect of extraction solvent/technique on their antioxidant ability, antimicrobial properties and phenolic profile. Nat. Prod. Res. 2020, 34, 46–52. [Google Scholar] [CrossRef] [PubMed]
- Meng, J.; Zhang, Y.; Wang, G.; Ji, M.; Wang, B.; He, G.; Wang, Q.; Bai, F.; Xu, K.; Yuan, D.; et al. Conduction of a chemical structure-guided metabolic phenotype analysis method targeting phenylpropane pathway via LC-MS: Ginkgo biloba and soybean as examples. Food Chem. 2022, 390, 133155. [Google Scholar] [CrossRef] [PubMed]
- Zhao, H.; Zhu, M.; Wang, K.; Yang, E.; Su, J.; Wang, Q.; Cheng, N.; Xue, X.; Wu, L.; Cao, W. Identification and quantitation of bioactive components from honeycomb (Nidus Vespae). Food Chem. 2020, 314, 126052. [Google Scholar] [CrossRef]
- Waridel, P.; Wolfender, J.-L.; Lachavanne, J.-B.; Hostettmann, K. Identification of the polar constituents of Potamogeton species by HPLC-UV with post-column derivatization, HPLC-MSn and HPLC-NMR, and isolation of a new ent-labdane diglycoside. Phytochemistry 2004, 65, 2401–2410. [Google Scholar] [CrossRef]
- Ibraheim, Z.Z.; Salem, H.A. Phytochemical and pharmacological studies on Pulicaria orientalis Jaub & Sp. Bull. Pharm. Sci. 2002, 25, 189–200. [Google Scholar]
- Williams, C.A.; Harborne, J.B.; Greenham, J.R.; Grayer, R.J.; Kite, G.C.; Eagles, J. Variations in lipophilic and vacuolar flavonoids among European Pulicaria species. Phytochemistry 2003, 64, 275–283. [Google Scholar] [CrossRef] [PubMed]
- Wollenweber, E.; Christ, M.; Dunstan, R.H.; Roitman, J.N.; Stevens, J.F. Exudate flavonoids in some Gnaphalieae and Inuleae (Asteraceae). Z. Naturforschung 2005, 60, 671–678. [Google Scholar] [CrossRef]
- Xiao, L.; Huang, Y.; Wang, Y.; Xu, J.; He, X. Anti-neuroinflammatory benzofurans and lignans from Praxelis clematidea. Fitoterapia 2020, 140, 104440. [Google Scholar] [CrossRef] [PubMed]
- Abo-Elghiet, F.; Rushdi, A.; Ibrahim, M.H.; Mahmoud, S.H.; Rabeh, M.A.; Alshehri, S.A.; El Menofy, N.G. Chemical Profile, Antibacterial, Antibiofilm, and Antiviral Activities of Pulicaria crispa Most Potent Fraction: An In Vitro and In Silico Study. Molecules 2023, 28, 4184. [Google Scholar] [CrossRef]
- Naqvi, S.A.R.; Shah, S.M.A.; Kanwal, L.; Saeed, M.; Ul-Haq, A.; Nisar, J.; Nisar, Z.; Akram, M. Antimicrobial and Antihypercholesterolemic Activities of Pulicaria gnaphalodes. Dose-Response 2020, 18, 1–6. [Google Scholar] [CrossRef]
- Thinina, A.C.; Karim, H.; Alia, M.M.; Karim, A. Evaluation and quantification of the inhibition of biofilm and planktonic forms of Klebsiella pneumoniae by the polyphenolic extract of Pulicaria crispa. J. Adv. Pharm. Technol. Res. 2020, 11, 117–122. [Google Scholar] [CrossRef] [PubMed]
- Jabir, N.R.; Khan, F.R.; Tabrez, S. Cholinesterase targeting by polyphenols: A therapeutic approach for the treatment of Alzheimer’s disease. CNS Neurosci. Ther. 2018, 24, 753–762. [Google Scholar] [CrossRef] [PubMed]
- Etsassala, N.G.E.R.; Badmus, J.A.; Marnewick, J.L.; Iwuoha, E.I.; Nchu, F.; Hussein, A.A. Alpha-Glucosidase and Alpha-Amylase Inhibitory Activities, Molecular Docking, and Antioxidant Capacities of Salvia aurita Constituents. Antioxidants 2020, 9, 1149. [Google Scholar] [CrossRef]
- Katalinić, M.; Rusak, G.; Barović, J.D.; Šinko, G.; Jelić, D.; Antolović, R.; Kovarik, Z. Structural aspects of flavonoids as inhibitors of human butyrylcholinesterase. Eur. J. Med. Chem. 2010, 45, 186–192. [Google Scholar] [CrossRef]
- Calderaro, A.; Patanè, G.T.; Tellone, E.; Barreca, D.; Ficarra, S.; Misiti, F.; Laganà, G. The Neuroprotective Potentiality of Flavonoids on Alzheimer’s Disease. Int. J. Mol. Sci. 2022, 23, 14835. [Google Scholar] [CrossRef] [PubMed]
- Oboh, G.; Agunloye, O.M.; Adefegha, S.A.; Akinyemi, A.J.; Ademiluyi, A.O. Caffeic and chlorogenic acids inhibit key enzymes linked to type 2 diabetes (in vitro): A comparative study. J. Basic Clin. Physiol. Pharmacol. 2015, 26, 165–170. [Google Scholar] [CrossRef]
- Zardi-Bergaoui, A.; Znati, M.; Harzallah-Skhiri, F.; Jannet, H.B. Caryophyllene sequiterpenes from Pulicaria vulgaris Gaertn.: Isolation, Structure Determination, Bioactivity and Structure—Activity Relationship. Chem. Biodivers. 2019, 16, e1800483. [Google Scholar] [CrossRef]
- de la Luz Cádiz-Gurrea, M.; Zengin, G.; Kayacık, O.; Lobine, D.; Mahomoodally, M.F.; Leyva-Jiménez, F.J.; Segura-Carretero, A. Innovative perspectives on Pulicaria dysenterica extracts: Phyto-pharmaceutical properties, chemical characterization and multivariate analysis. J. Sci. Food Agric. 2019, 99, 6001–6010. [Google Scholar] [CrossRef]
- Bakthir, H.; Ali, N.A.; Arnold, N.; Teichert, A.; Wessjohann, L. Anticholinesterase activity of endemic plant extracts from Soqotra. Afr. J. Tradit. Complement. Altern. Med. 2011, 8, 296–299. [Google Scholar] [CrossRef] [PubMed]
- Giuberti, G.; Rocchetti, G.; Lucini, L. Interactions between phenolic compounds, amylolytic enzymes and starch: An updated overview. Curr. Opin. Food Sci. 2020, 31, 102–113. [Google Scholar] [CrossRef]
- Takahama, U.; Hirota, S. Interactions of flavonoids with α-amylase and starch slowing down its digestion. Food Funct. 2018, 9, 677–687. [Google Scholar] [CrossRef] [PubMed]
- Nyambe-Silavwe, H.; Williamson, G. Chlorogenic and phenolic acids are only very weak inhibitors of human salivary α-amylase and rat intestinal maltase activities. Food Res. Int. 2018, 113, 452–455. [Google Scholar] [CrossRef] [PubMed]
- Mohammed, H.A.; Abdelwahab, M.F.; El-Ghaly, E.S.M.; Ragab, E.A. Phytochemical Characterization, In Vitro Anti-Inflammatory, Anti-Diabetic, and Cytotoxic Activities of the Edible Aromatic Plant; Pulicaria jaubertii. Molecules 2021, 26, 203. [Google Scholar] [CrossRef]
- Mohamed, M.S.; Saleh, A.M.; Abdel-Farid, I.B.; El-Naggar, S.A. Growth, hydrolases and ultrastructure of Fusarium oxysporum as affected by phenolic rich extracts from several xerophytic plants. Pestic. Biochem. Physiol. 2017, 141, 57–64. [Google Scholar] [CrossRef]
- Marwah, R.G.; Fatope, M.O.; Al Mahrooqi, R.; Varma, G.B.; Al Abadi, H.; Al-Burtamani, S.K.S. Antioxidant capacity of some edible and wound healing plants in Oman. Food Chem. 2007, 101, 465–470. [Google Scholar] [CrossRef]
- Mothana, R.A.; Kriegisch, S.; Harms, M.; Wende, K.; Lindequist, U. Assessment of selected Yemeni medicinal plants for their in vitro antimicrobial, anticancer, and antioxidant activities. Pharm. Biol. 2011, 49, 200–210. [Google Scholar] [CrossRef]
- Foudah, A.I.; Alam, A.; Soliman, G.A.; Salkini, M.A.; Ahmed, E.I.; Yusufoglu, H.S. Pharmacognostical, antioxidant and antimicrobial studies of aerial part of Pulicaria crispa (Family: Asteraceae). Bull. Environ. Pharmacol. Life Sci. 2015, 4, 19–27. [Google Scholar]
- Kozarević Cilović, E.; Dautović, E.; Halilčević, D.; Softić, A.; Srabović, N.; Šarić-Kundalić, B.; Delic, N.; Kolarevic, L.; Mekić, L.; Ibišević, M.; et al. Antioxidant and Cytotoxic Activities of Pulicaria dysenterica Methanol Extracts. Int. Res. J. Pure Appl. Chem. 2022, 23, 23–32. [Google Scholar] [CrossRef]
- Farhat, M.B.; Chaouch-Hamada, R.; Sotomayor, J.A.; Landoulsi, A.; Jordán, M.J. Antioxidant potential of Salvia officinalis L. residues as affected by the harvesting time. Ind. Crops Prod. 2014, 54, 78–85. [Google Scholar] [CrossRef]
- Rahmouni, F.; Badraoui, R.; Ben-Nasr, H.; Bardakci, F.; Elkahoui, S.; Siddiqui, A.J.; Saeed, M.; Snoussi, M.; Saoudi, M.; Rebai, T. Pharmacokinetics and therapeutic potential of Teucrium polium against liver damage associated hepatotoxicity and oxidative injury in rats: Computational, biochemical and histological studies. Life 2022, 12, 1092. [Google Scholar] [CrossRef]
- Mhadhbi, N.; Issaoui, N.; Hamadou, W.S.; Alam, J.M.; Elhadi, A.S.; Adnan, M.; Nahili, H.; Badraoui, R. Physico-Chemical Properties, Pharmacokinetics, Molecular Docking and In-Vitro Pharmacological Study of a Cobalt (II) Complex Based on 2-Aminopyridine. ChemistrySelect 2022, 7, e20210359. [Google Scholar] [CrossRef]
- Badraoui, R.; Saoudi, M.; Hamadou, W.S.; Elkahoui, S.; Siddiqui, A.J.; Alam, J.M.; Jamal, A.; Adnan, M.; Suliemen, A.M.E.; Alreshidi, M.M.; et al. Antiviral Effects of Artemisinin and Its Derivatives against SARS-CoV-2 Main Protease: Computational Evidences and Interactions with ACE2 Allelic Variants. Pharmaceuticals 2022, 15, 129. [Google Scholar] [CrossRef]
- Alreshidi, M.; Badraoui, R.; Adnan, M.; Patel, M.; Alotaibi, A.; Saeed, M.; Ghandourah, M.; Al-Motair, K.A.; Arif, I.A.; Albulaihed, Y.; et al. Phytochemical profiling, antibacterial, and antibiofilm activities of Sargassum sp. (brown algae) from the Red Sea: ADMET prediction and molecular docking analysis. Algal Res. 2023, 69, 102912. [Google Scholar]
- Noumi, E.; Ahmad, I.; Bouali, N.; Patel, H.; Ghannay, S.; ALrashidi, A.A.; Abdulhakeem, M.A.; Patel, M.; Ceylan, O.; Badraoui, R.; et al. Thymus musilii Velen. Methanolic Extract: In Vitro and In Silico Screening of Its Antimicrobial, Antioxidant, Anti-Quorum Sensing, Antibiofilm, and Anticancer Activities. Life 2023, 13, 62. [Google Scholar] [CrossRef] [PubMed]
- Sarker, S.D.; Nahar, L.; Kumarasamy, Y. Microtitre plate-based antibacterial assay incorporating resazurin as an indicator of cell growth, and its application in the in vitro antibacterial screening of phytochemicals. Methods 2007, 42, 321–324. [Google Scholar] [CrossRef] [PubMed]
- Fratianni, F.; d’Acierno, A.; Ombra, M.N.; Amato, G.; De Feo, V.; Ayala-Zavala, J.F.; Coppola, R.; Nazzaro, F. Fatty Acid Composition, Antioxidant, and in vitro Anti-inflammatory Activity of Five Cold-Pressed Prunus Seed Oils, and Their Anti-biofilm Effect Against Pathogenic Bacteria. Front. Nutr. 2021, 8, 775751. [Google Scholar] [CrossRef] [PubMed]
- Nazzaro, F.; Polito, F.; Amato, G.; Caputo, L.; Francolino, R.; d’Acierno, A.; Fratianni, F.; Candido, V.; Coppola, R.; De Feo, V. Chemical composition of Essential Oils of bulbs and aerial parts of two cultivars of Allium sativum and their antibiofilm activity against food and nosocomial pathogens. Antibiotics 2022, 11, 724. [Google Scholar] [CrossRef]
- Xiang, J.; Apea-Bah, F.B.; Ndolo, V.U.; Katundu, M.C.; Beta, T. Profile of phenolic compounds and antioxidant activity of finger millet varieties. Food Chem. 2019, 275, 361–368. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Xu, Y.; Xiang, J.; Zheng, B.; Yuan, Y.; Luo, D.; Fan, J. Comparative evaluation on phenolic profiles, antioxidant properties and α-glucosidase inhibitory effects of different milling fractions of foxtail millet. J. Cereal Sci. 2021, 99, e103217. [Google Scholar] [CrossRef]
- Amamcharla, J.K.; Metzger, L.E. Modification of the ferric reducing antioxidant power (FRAP) assay to determine the susceptibility of raw milk to oxidation. Int. Dairy J. 2014, 34, 177–179. [Google Scholar] [CrossRef]
- Waterhouse, A.L. Determination of total phenolics. Curr. Protoc. Food Anal. Chem. 2002, 6, 1–8. [Google Scholar]
- Baba, S.A.; Malik, S.A. Determination of total phenolic and flavonoid content, antimicrobial and antioxidant activity of a root extract of Arisaema jacquemontii Blume. J. Taibah Univ. Sci. 2015, 9, 449–454. [Google Scholar] [CrossRef]
- Zheng, B.; Yuan, Y.; Xiang, J.; Jin, W.; Johnson, J.B.; Li, Z.; Wang, C.; Luo, D. Green extraction of phenolic compounds from foxtail millet bran by ultrasonic-assisted deep eutectic solvent extraction: Optimization, comparison and bioactivities. LWT 2022, 154, 112740. [Google Scholar] [CrossRef]
- Jaradat, N.; Al-Maharik, N.; Abdallah, S.; Shawahna, R.; Mousa, A.; Qtishat, A. Nepeta curviflora essential oil: Phytochemical composition, antioxidant, anti-proliferative and anti-migratory efficacy against cervical cancer cells, and α-glucosidase, α-amylase and porcine pancreatic lipase inhibitory activities. Ind. Crops Prod. 2020, 158, 112946. [Google Scholar] [CrossRef]
- Nguyen, T.K.; Tran, L.T.T.; Viet, D.H.; Thai, P.H.; Ha, T.P.; Ty, P.V.; Huu, D.T.T. Xanthine oxidase, α-glucosidase and α-amylase inhibitory activities of the essential oil from Piper lolot: In vitro and in silico studies. Heliyon 2023, 9, e19148. [Google Scholar] [CrossRef]
- Akacha, A.; Badraoui, R.; Rebai, T.; Zourgui, L. Effect of Opuntia Ficus Indica Extract on Methotrexate-Induced Testicular Injury: A Biochemical, Docking and Histological Study. J. Biomol. Struct. Dyn. 2020, 40, 4341–4351. [Google Scholar] [CrossRef]
- Badraoui, R.; Mannai, G.; Siddiqui, A.J.; Pacioglu, O.; Rudayni, H.A.; Boufahja, F.; Essid, N. How toxic is the COVID-19 drug azithromycin in the presence of Posidonia oceanica? Toxicokinetics and experimental approach of meiobenthic nematodes from a metallically pristine area. Environ. Pollut. 2023, 319, 121007. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | Family | Retention Time (min) | Measured m/z | Molecular Formula | Fragment | Fragment Formula | Fragment Ion (m/z) | Δppm | Identification | PM | PW |
|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | Amino acid derivative | 1.30 | 294.1545 | C12H23O7N | [M-H2O + H]+ [M-2H2O + H]+ [M-CHO2 + H]+ | C12H22O6N C12H20O5N C11H22O5N | 276.1443 258.1342 248.1494 | 0.674 2.521 0.527 | N-Fructosyl (iso)leucine | X | X |
| 2 | Amino acid derivative | 2.10 | 166.0862 | C9H11O2N | [M-NH2 + H]+ [M-CHO2 + H]+ | C9H9O2 C8H10N | 149.0594 120.0806 | −1.919 −1.798 | L-Phenylalanine | X | X |
| 3 | Phenolic acid | 7.10 | 355.1019 | C16H18O9 | [M-H2O + H]+ [M-C7H12O6 + H]+ [M-C7H12O6-CO-H2O + H]+ | C16H17O8 C9H7O3 C8H5O | 337.0926 163.0391 117.0335 | 2.362 0.855 −0.097 | Chlorogenic acid | X | X |
| 4 | α-β-Unsaturated γ-lactone | 7.52 | 199.1328 | C11H18O3 | [M-H2O + H]+ [M-2H2O + H]+ | C11H17O2 C11H15O | 181.1221 163.1104 | −1.139 −8.225 | (−)-Hydroxydihydrobovolide | X | X |
| 5 | Phenolic acid | 7.65 | 339.0926 | C16H18O8 | [M-C7H12O6 + H]+ [M-C7H12O6-CO + H]+ | C9H7O2 C8 H7O | 147.0438 119.0484 | −1.469 −6.144 | O-Coumaroylquinic acid | X | X |
| 6 | Phenolic acid | 7.82 | 369.1042 | C17H20O9 | [M-C7H12O6 + H]+ [M-C7H12O6-CH3O + H]+ | C10H9O3 C9 H5O2 | 177.0545 145.0285 | −0.569 0.648 | Feruloylquinic acid | X | |
| 7 | Unsaturated ketone | 8.12 | 169.1222 | C10H16O2 | [M-H2O + H]+ [M-H2O-C3H6 + H]+ | C10H15O C7H9O | 151.1108 109.0658 | −6.496 9.522 | 8-Hydroxycarvotanacetone | X | X |
| 8 | Flavonoid | 8.30 | 465.1020 | C21H20O12 | [M-H2O + H]+ [M-C6H12O6 + H]+ | C21H19O11 C15H11O7 | 447.0923 303.0497 | 0.363 −0.723 | Isoquercetin (quercetin 3-O-β-D-glucoside) | X | |
| 9 | Phenolic acid | 8.80 | 517.1329 | C25H24O12 | [M-H2O + H]+ [M-C9H7O3 + H]+ [M-C9H7O3-C7H12O6 + H]+ | C25H23O11 C16H19O9 C9H7O3 | 499.1230 355.1027 163.0388 | −1.058 0.849 −0.740 | (−)-3,5-Dicaffeoylquinic acid | X | X |
| 10 | Sesquiterpene hydrocarbon | 9.74 | 203.1792 | C15H22 | [M-CH(CH3)2 + H]+ [M-CH(CH3)2-CH3 + H]+ | C12H17 C11H15 | 161.1325 147.1165 | 0.080 −1.883 | Calamenene | X | X |
| 11 | Flavonoid | 11.12 | 317.0652 | C16H12O7 | [M-H2O + H]+ [M-C6H4O2 + H]+ [M-C6H4O2-H2O + H]+ | C16H11O6 C10H9O5 C10H7O4 | 299.0550 209.0448 191.0336 | 0.018 1.771 −1.493 | Rhamnetin | X | X |
| 12 | Flavonoid | 11.35 | 331.0806 | C17H14O7 | [M-CH3 + H]+ [M-CH3-H2O + H]+ | C16H12O7 C16H11O6 | 316.0573 299.0568 | −1.310 6.138 | Quercetin dimethyl ether | X | X |
| 13 | Flavonoid | 11.77 | 287.0912 | C16H14O5 | [M-H2O + H]+ [M-C8H6O + H]+ | C16H13O4 C8H7O4 | 269.0801 167.0338 | −2.584 −0.151 | Sakuranetin | X | |
| 14 | Flavonoid | 11.95 | 301.0705 | C16H12O6 | [M-CH3 + H]+ | C15H10O6 | 286.0784 | 1.324 | Chrysoeriol | X | X |
| 15 | Flavonoid | 12.25 | 315.0864 | C17H14O6 | [M-CH3 + H]+ | C16H12O6 | 300.0630 | 0.501 | Dihydroxy–dimethoxyflavone | X | X |
| 16 | Flavonoid | 12.32 | 345.0969 | C18H16O7 | [M-CH3 + H]+ [M-2CH3 + H]+ | C17H14 O7 C16H11O7 | 330.0733 315.0509 | −0.316 3.177 | Dihydroxy–trimethoxyflavone | X | X |
| 17 | Benzofuran dimer | 13.15 | 451.1380 | C25H22O8 | [M-CH2OH-H2O + H]+ | C24H18O6 | 402.3573 | −1.057 | 1-[9-(6-Acetyl-5-hydroxy-2-benzofuranyl)-6,7,8,9-tetrahydro-2,6-dihydroxy-6-(hydroxymethyl)-3-dibenzofuranyl] ethanone | X |
| A. baumannii | E. coli | L. monocytogenes | P. aeruginosa | S. aureus | |
|---|---|---|---|---|---|
| P. incisa (Ext. H2O) | 35 ± 4 a | 35 ± 3 a | 35 ± 3 a | 33 ± 2 a | 36 ± 2 a |
| P. incisa (Ext. MeOH) | 35 ± 2 a | 30 ± 3 a | 35 ± 4 a | 32 ± 4 a | 32 ± 2 a |
| Tetracycline | 25 ± 2 | 25 ± 2 | 32 ± 1 | 28 ± 1 | 28 ± 1 |
| Percentage of Inhibition Compared to the Control | ||||
|---|---|---|---|---|
| CV (24 h) | P. incisa (Ext. H2O) 10 μg/mL | P. incisa (Ext. H2O) 20 μg/mL | P. incisa (Ext. MeOH) 10 μg/mL | P. incisa (Ext. MeOH) 20 μg/mL |
| A. baumannii | 19.13 **** ± 1.02 | 19.37 **** ± 1.26 | 12.35 *** ± 0.46 | 19.53 **** ± 1.18 |
| E. coli | 4.38 * ± 0.12 | 25.5 **** ± 1.57 | 32.87 **** ± 1.88 | 44.62 **** ± 2.87 |
| L. monocytogenes | 14.93 **** ± 1.02 | 17.21 **** ± 1.16 | 3.32 * ± 0.11 | 18.10 **** ± 1.04 |
| P. aeruginosa | 24.36 **** ± 1.09 | 36.68 **** ± 1.44 | 29.32 **** ± 2.86 | 55.78 **** ± 2.55 |
| S. aureus | 8.26 ** ± 0.11 | 32.54 **** ± 1.43 | 31.31 **** ± 2.97 | 51.27 **** ± 2.07 |
| Percentage of Inhibition Compared to the Control | ||||
|---|---|---|---|---|
| MTT (24 h) | P. incisa (Ext. H2O) 10 μg/mL | P. incisa (Ext. H2O) 20 μg/mL | P. incisa (Ext. MeOH) 10 μg/mL | P. incisa (Ext. MeOH) 20 μg/mL |
| A. baumannii | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 |
| E. coli | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 |
| L. monocytogenes | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 |
| P. aeruginosa | 0.00 ± 0.00 | 19.98 **** ± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 |
| S. aureus | 49.77 **** ± 3.88 | 77.94 **** ± 4.06 | 12.74 **** ± 1.02 | 30.52 **** ± 2.84 |
| IC50 (µg/mL) | ||||
|---|---|---|---|---|
| AChE | BChE | α-Amylase | α-Glucosidase | |
| Pulicaria incisa (aqueous extract) | 1621.5 ± 109.6 b | 1092.5 ± 6.4 b | n.a. | 393.9 ± 39.9 a |
| Pulicaria incisa (methanolic extract) | 292.3 ± 1.5 a | 236.9 ± 15.7 a | n.a. | 1936.5 ± 164.7 b |
| Galantamine | 0.9 ± 0.4 | 4.6 ± 1.5 | – | – |
| Acarbose | – | – | 11 ± 4.2 | 963 ± 0.8 |
| Antioxidant Activity | |||||
|---|---|---|---|---|---|
| TPC mg (GAE)/g | TFC mg (QE)/g | DPPH (IC50) μg/mL | FRAP mmol (Fe2+)/g | ABTS TEAC mg (TE)/g | |
| P. incisa (ext. H2O) | 56.6 a ± 2.1 | 11.24 a ± 0.8 | 64.27 b ± 0.58 | 0.93 a ± 0.09 | 84.42 a ± 7.2 |
| P. incisa (ext. MeOH) | 84.80 b ± 2.8 | 28.30 b ± 1.2 | 23.29 a ± 0.38 | 2.99 b ± 0.14 | 201.86 b ± 7.8 |
| Trolox | – | – | 3.65 ± 0.01 | 10.30 a ± 0.08 | – |
| Compound No. | Binding Affinity (kcal × mol−1) | ||
|---|---|---|---|
| 1JIJ | 2XCT | 2UVO | |
| 1 | −8.0 | −7.4 | −6.8 |
| 2 | −6.3 | −6.1 | −5.4 |
| 3 | −8.6 | −7.5 | −7.1 |
| 4 | −6.9 | −5.7 | −5.4 |
| 5 | −8.1 | −7.8 | −7.0 |
| 6 | −9.0 | −7.0 | −6.6 |
| 7 | −6.1 | −5.3 | −5.1 |
| 8 | −7.7 | −7.8 | −7.3 |
| 9 | −10.4 | −9.1 | −7.9 |
| 10 | −10.1 | −8.4 | −7.8 |
| 11 | −9.6 | −8.0 | −6.7 |
| 12 | −9.1 | −7.8 | −6.8 |
| 13 | −8.6 | −7.5 | −6.8 |
| 14 | −9.7 | −8.0 | −7.2 |
| 15 | −8.4 | −7.6 | −6.7 |
| 16 | −9.7 | −7.8 | −6.9 |
| 17 | −7.7 | −7.0 | −6.8 |
| Receptor | Chemical Compound No. | No. Conventional H-Bonds | Closest Interacting Residues | Closest Residue (Distance, Å) | No. Closest Interacting Residues |
|---|---|---|---|---|---|
| 1JIJ | 9 | 9 | Conventional H-Bond: ASP40, LYS84, LYS84, ARG88, VAL224, TYR36, GLN190, PRO222, PRO222 Carbon H-Bond: ALA39, GLY193 Hydrophobic: LEU70, VAL224 | VAL224:HN (2.070) | 10 |
| 10 | 10 | Conventional H-Bond: LYS84, LYS84, GLY193, GLN196, VAL224, THR75, TYR170, ASP177, ASP195, PRO220 Carbon H-Bond: GLY193, ASP195 | VAL224:HN (1.980) | 9 | |
| 14 | 6 | Conventional H-Bond: LYS84, LYS84, LYS84, ARG88, ASP177, GLN190 Carbon H-Bond: ASP40 Electrostatic: ASP80 | GLN190:OE1 (2.064) | 6 | |
| 16 | 8 | Conventional H-Bond: LYS84, LYS84, LYS84, ARG88, TYR36, GLN190, THR75, ASP177 Carbon H-Bond: ASP40 Electrostatic: ASP80 Hydrophobic: HIS50, LEU70 | LYS84:HZ1 (2.184) | 10 | |
| 2XCT | 9 | 6 | Conventional H-Bond: SER88, THR222, GLY34, VAL12, THR222, GLN54 Carbon H-Bond: ASP86 Electrostatic: ARG51, ASP218 Hydrophobic: ARG51, PRO120 | THR222:OG1 (1.917) | 9 |
| 10 | 6 | Conventional H-Bond: ARG51, SER88, THR222, SER301, SER301, LYS49 Carbon H-Bond: GLY220 Hydrophobic: PHE58, TYR225 | SER301:HG (2.023) | 8 | |
| 11 | 6 | Conventional H-Bond: ARG195, ASP214, SER190, ASP191, GLU193, ALA303 Carbon H-Bond: GLN227 Hydrophobic: ILE213 | ASP214:HN (1.961) | 8 | |
| 14 | 5 | Conventional H-Bond: ARG192, THR221, ASP218, THR221, ASP37 Electrostatic: ASP218 Hydrophobic: ILE305, ALA133 | ASP218:OD2 (2.340) | 6 | |
| 2UVO | 9 | 10 | Conventional H-Bond: SER43, SER43, NDG1173, NAG1174, NDG1173, NAG1174, NDG1173, NAG1174, ASN15, GLN59 Hydrophobic: PHE69, ALA39 | SER43:HG (2.176) | 7 |
| 10 | 8 | Conventional H-Bond: GLY113, NAG1176, ILE155, CYS153, GOL1177, NAG1176, ASP129, CYS153 Electrostatic: ASP129 | NAG1176:H3 (2.057) | 6 | |
| 8 | 10 | Conventional H-Bond: NAG1176, GOL1177, GOL1177, GOL1177, NAG1176, ASP129, GOL1177, GOL1177, GOL1177, ILE155 Hydrophobic: ALA125, ALA125 | NAG1176:H3 (2.092) | 5 | |
| 14 | 4 | Conventional H-Bond: TYR159, GOL1177, ASN101, NAG1176 Electrostatic: GOL1177, ASP129 Carbon H-Bond: GOL1177, GOL1177 Hydrophobic: ALA125, ALA125, ILE155 | TYR159:HH (2.223) | 7 |
| Entry | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 | 13 | 14 | 15 | 16 | 17 |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Lipophilicity and Physicochemical Properties | |||||||||||||||||
| TPSA | 139.48 | 63.32 | 354.31 | 46.53 | 144.52 | 153.75 | 37.30 | 210.51 | 211.28 | 211.28 | 210.36 | 190.36 | 75.99 | 100.13 | 89.13 | 98.36 | 98.36 |
| Log Po/w (iLOGP) | 0.71 | 1.08 | 0.87 | 2.43 | 0.83 | 1.64 | 1.96 | 0.94 | 1.50 | 1.92 | 2.23 | 2.61 | 2.41 | 2.44 | 2.58 | 2.98 | 2.84 |
| Consensus Log Po/w | −1.62 | −0.01 | −0.39 | 2.24 | −0.08 | −0.00 | 1.57 | −0.48 | 0.83 | 0.92 | 1.63 | 2.20 | 2.25 | 2.18 | 2.47 | 2.52 | 2.43 |
| Log S (ESOL) solubility | 0.12 | −0.08 | −1.62 | −2.13 | −1.75 | −1.84 | −1.51 | −3.04 | −3.35 | −3.36 | −3.36 | −4.25 | −3.70 | −4.06 | −4.20 | −4.35 | −4.16 |
| Pharmacokinetics | |||||||||||||||||
| GI absorption | Low | High | Low | High | Low | Low | High | Low | Low | Low | High | High | High Yes | High | High | High | High |
| BBB permeant | No | No | No | Yes | No | No | Yes | No | No | No | No | No | Yes | No | No | No | No |
| P-gp substrate | No | No | No | No | No | No | No | No | Yes | Yes | No | No | No | No | No | No | No |
| CYP1A2 | No | No | No | No | No | No | No | No | No | No | Yes | Yes | Yes | Yes | Yes | Yes | Yes |
| CYP2C19 | No | No | No | No | No | No | No | No | No | No | No | No | Yes | No | No | No | No |
| CYP2C9 | No | No | No | No | No | No | No | No | No | No | No | Yes | No | Yes | Yes | Yes | Yes |
| CYP2D6 | No | No | No | No | No | No | No | No | No | No | Yes | Yes | No | Yes | Yes | Yes | Yes |
| CYP3A4 | No | No | No | No | No | No | No | No | No | No | Yes | Yes | Yes | Yes | Yes | Yes | Yes |
| Log Kp (skin permeation) | −10.24 | −8.39 | −8.76 | −6.01 | −8.41 | −8.62 | −6.55 | −8.88 | −8.37 | −8.37 | −6.90 | −5.99 | −6.02 | −5.93 | −5.86 | −5.97 | −6.17 |
| Druglikeness and Medicinal Chemistry | |||||||||||||||||
| Lipinski | Yes | Yes | Yes | Yes | Yes | Yes | Yes | No | No | No | Yes | Yes | Yes | Yes | Yes | Yes | Yes |
| Ghose | No | Yes | No | Yes | No | No | Yes | No | No | No | Yes | Yes | Yes | Yes | Yes | Yes | Yes |
| Veber | Yes | Yes | No | Yes | No | No | Yes | No | No | No | Yes | Yes | Yes | Yes | Yes | Yes | Yes |
| Ergan | No | Yes | No | Yes | No | No | Yes | No | No | No | Yes | Yes | Yes | Yes | Yes | Yes | Yes |
| Muegge | No | No | No | No | Yes | No | No | No | No | No | Yes | Yes | Yes | Yes | Yes | Yes | Yes |
| Leadlikeness | 0.55 | 0.55 | 0.11 | 0.55 | 0.56 | 0.55 | 0.55 | 0.17 | 0.11 | 0.11 | 0.55 | 0.55 | 0.55 | 0.55 | 0.55 | 0.55 | 0.55 |
| Synthetic accessibility | 3.96 | 1.46 | 4.16 | 3.43 | 4.07 | 4.27 | 3.19 | 5.32 | 4.84 | 4.81 | 3.30 | 3.40 | 3.11 | 3.06 | 3.22 | 3.31 | 3.52 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alreshidi, M.; Abdulhakeem, M.A.; Badraoui, R.; Amato, G.; Caputo, L.; De Martino, L.; Nazzaro, F.; Fratianni, F.; Formisano, C.; De Feo, V.; et al. Pulicaria incisa (Lam.) DC. as a Potential Source of Antioxidant, Antibacterial, and Anti-Enzymatic Bioactive Molecules: Phytochemical Constituents, In Vitro and In Silico Pharmacological Analysis. Molecules 2023, 28, 7439. https://doi.org/10.3390/molecules28217439
Alreshidi M, Abdulhakeem MA, Badraoui R, Amato G, Caputo L, De Martino L, Nazzaro F, Fratianni F, Formisano C, De Feo V, et al. Pulicaria incisa (Lam.) DC. as a Potential Source of Antioxidant, Antibacterial, and Anti-Enzymatic Bioactive Molecules: Phytochemical Constituents, In Vitro and In Silico Pharmacological Analysis. Molecules. 2023; 28(21):7439. https://doi.org/10.3390/molecules28217439
Chicago/Turabian StyleAlreshidi, Mousa, Mohammad A. Abdulhakeem, Riadh Badraoui, Giuseppe Amato, Lucia Caputo, Laura De Martino, Filomena Nazzaro, Florinda Fratianni, Carmen Formisano, Vincenzo De Feo, and et al. 2023. "Pulicaria incisa (Lam.) DC. as a Potential Source of Antioxidant, Antibacterial, and Anti-Enzymatic Bioactive Molecules: Phytochemical Constituents, In Vitro and In Silico Pharmacological Analysis" Molecules 28, no. 21: 7439. https://doi.org/10.3390/molecules28217439