

Insights on Antitumor Activity and Mechanism of Natural Benzophenanthridine Alkaloids

Abstract
:1. Introduction
2. Antitumor Activities of Benzophenanthridine Alkaloids from Natural Products
2.1. Antitumor Activities of Benzophenanthridine Alkaloids from Papaveraceae Plants
2.1.1. Antitumor Activities of Benzophenanthridine Alkaloids from Papaver SPP
2.1.2. Antitumor Activities of Benzophenanthridine Alkaloids from Corydalis saxicola Bunting
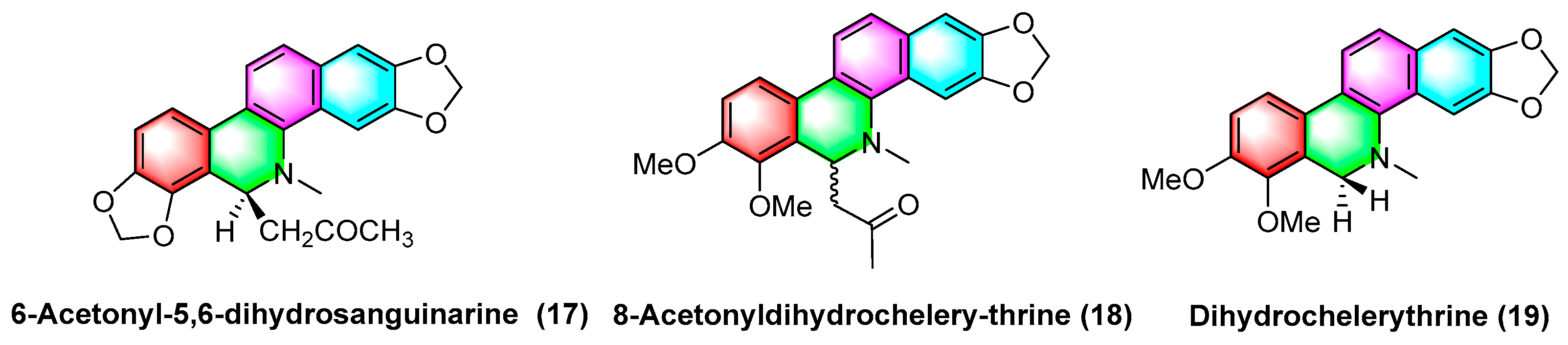
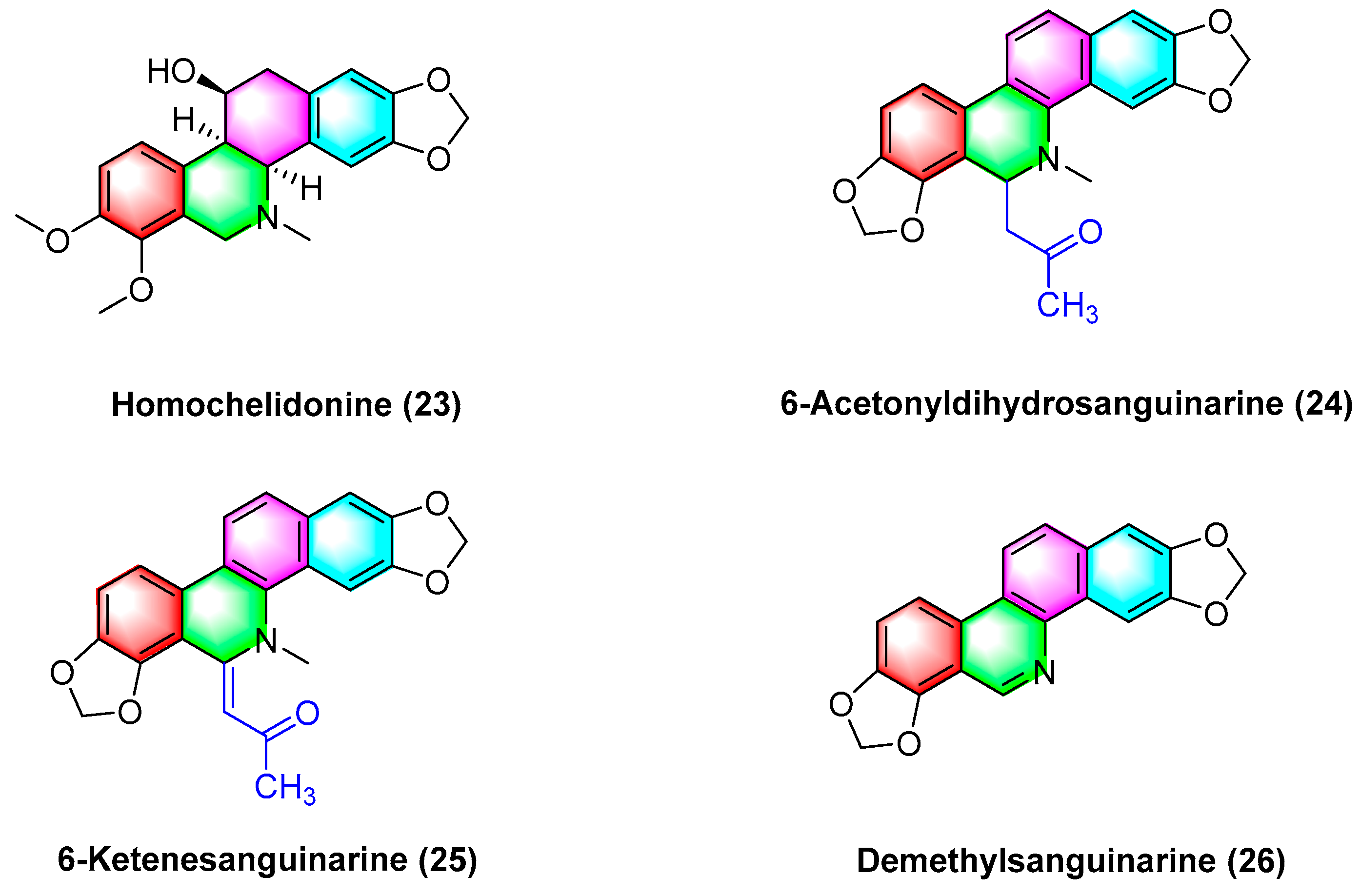
2.1.3. Antitumor Activities of Benzophenanthridine Alkaloids from Chelidonium
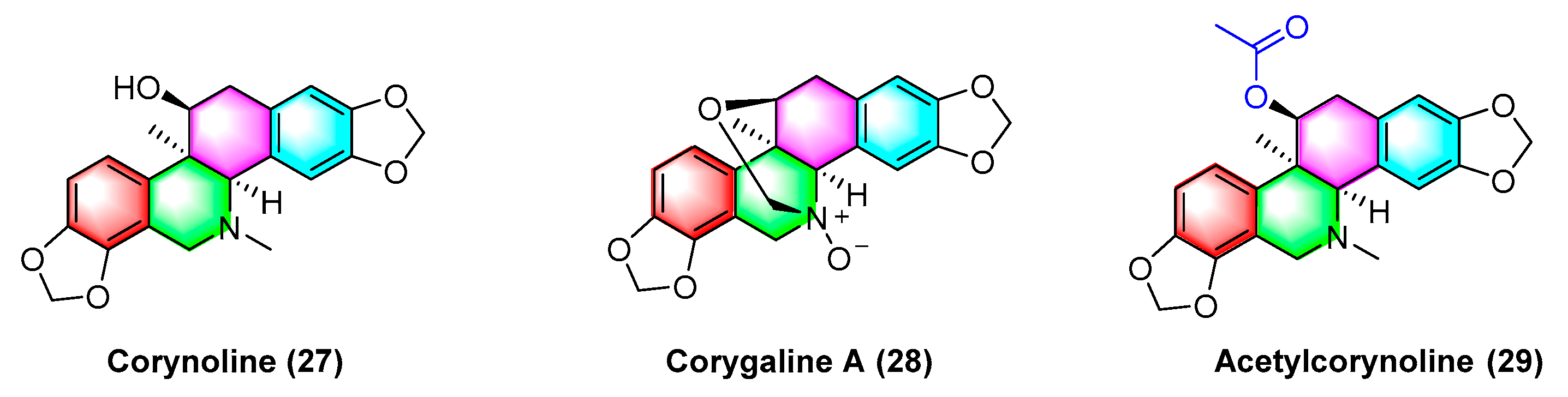
2.1.4. Antitumor Activities of Benzophenanthridine Alkaloids from Corydalis
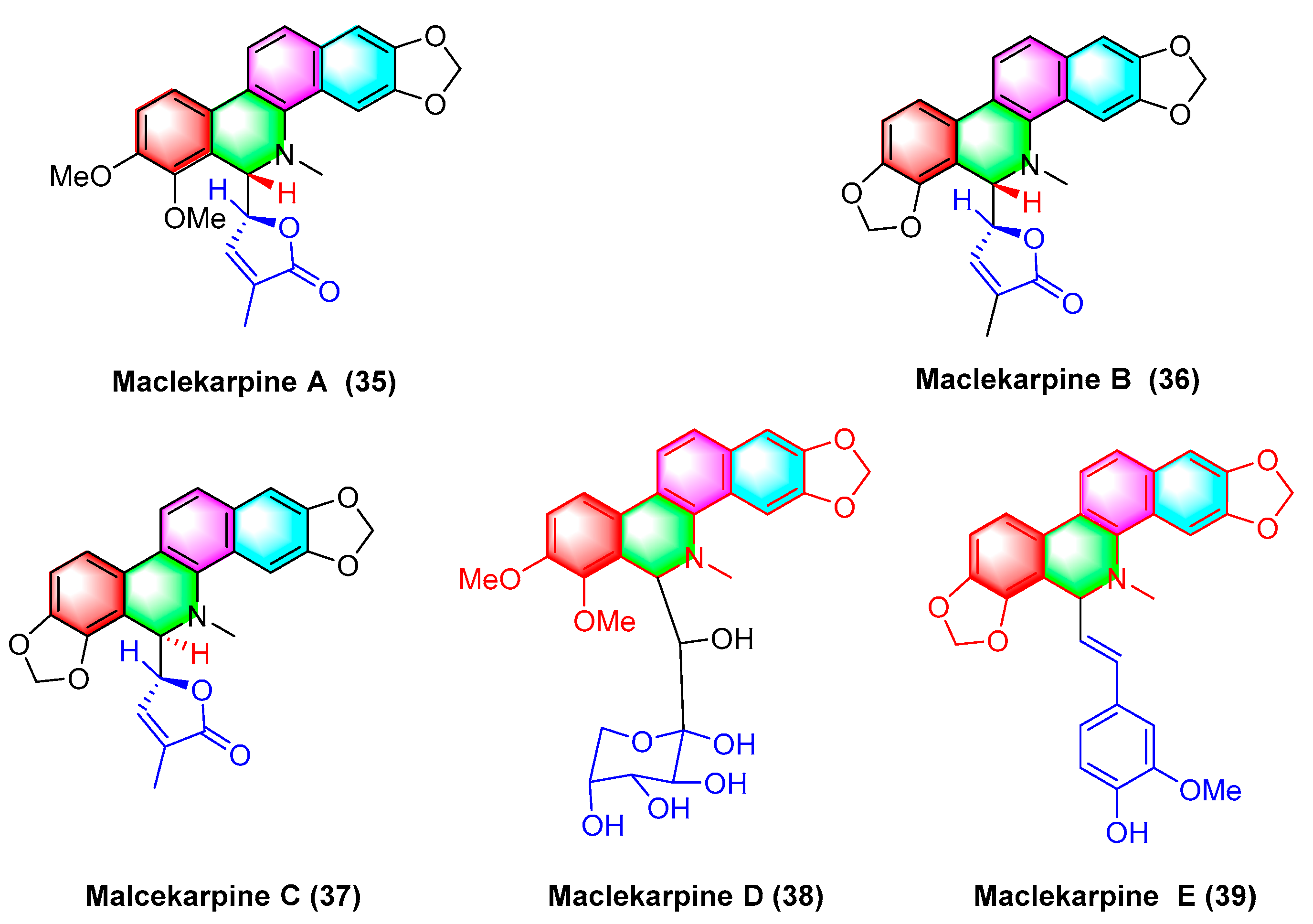
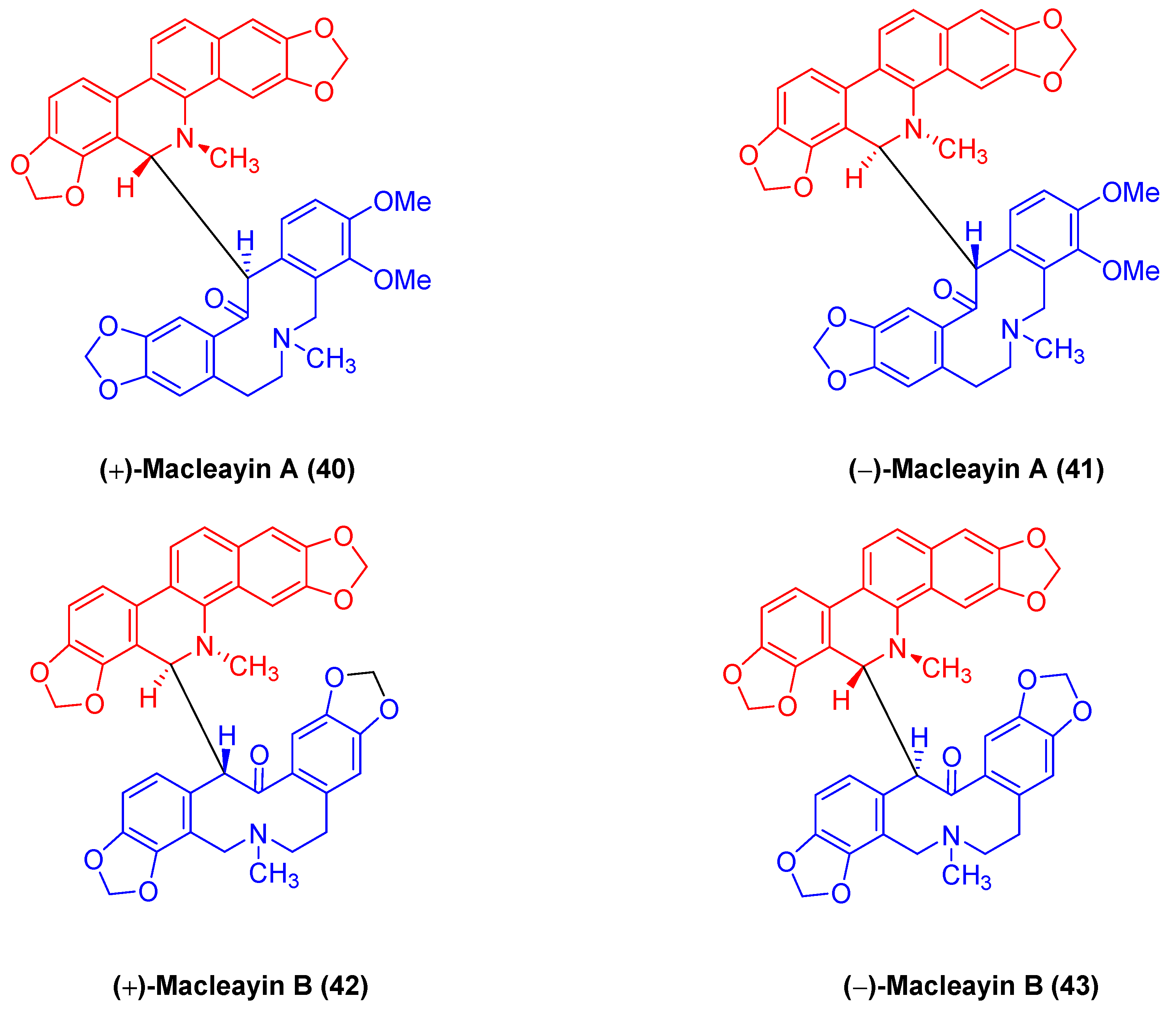
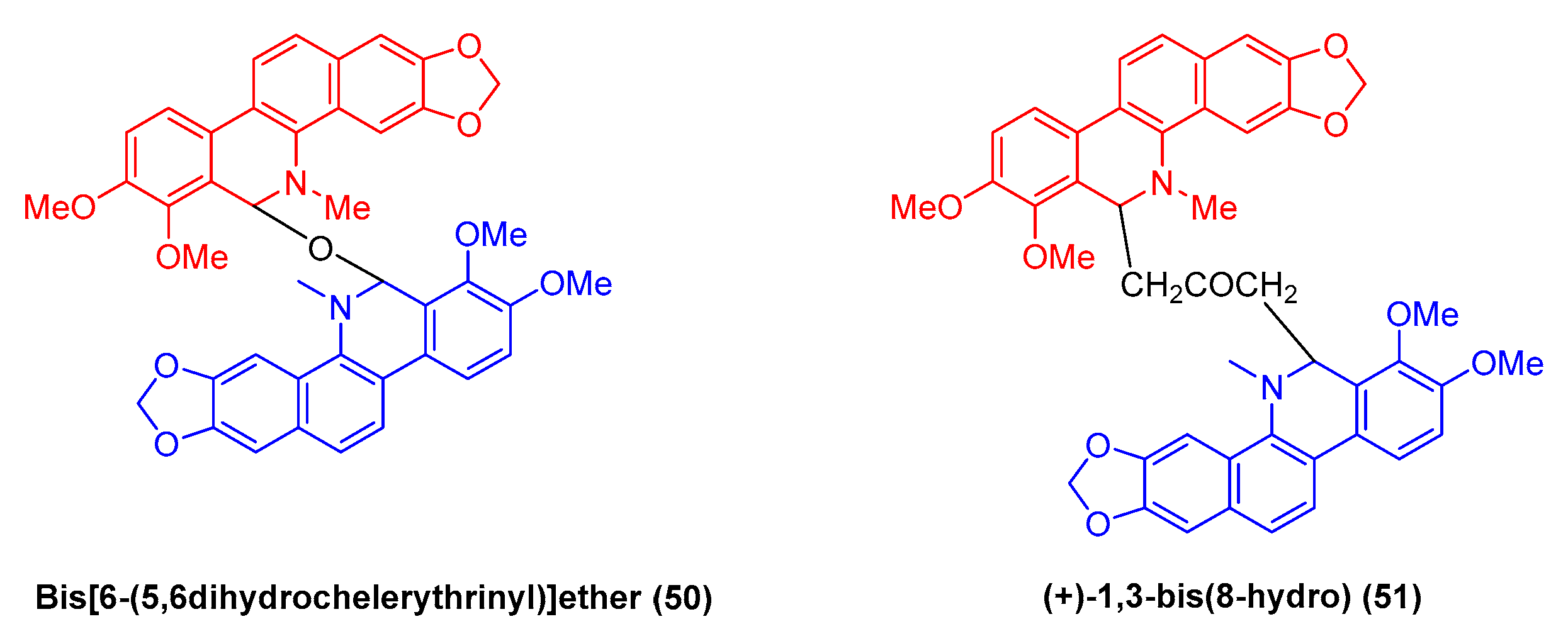
2.1.5. Antitumor Activities of Benzophenanthridine Alkaloids from Macleaya cordata
2.2. Antitumor Activities of Benzophenanthridine Alkaloids from Plants of the Genus Tanneraceae
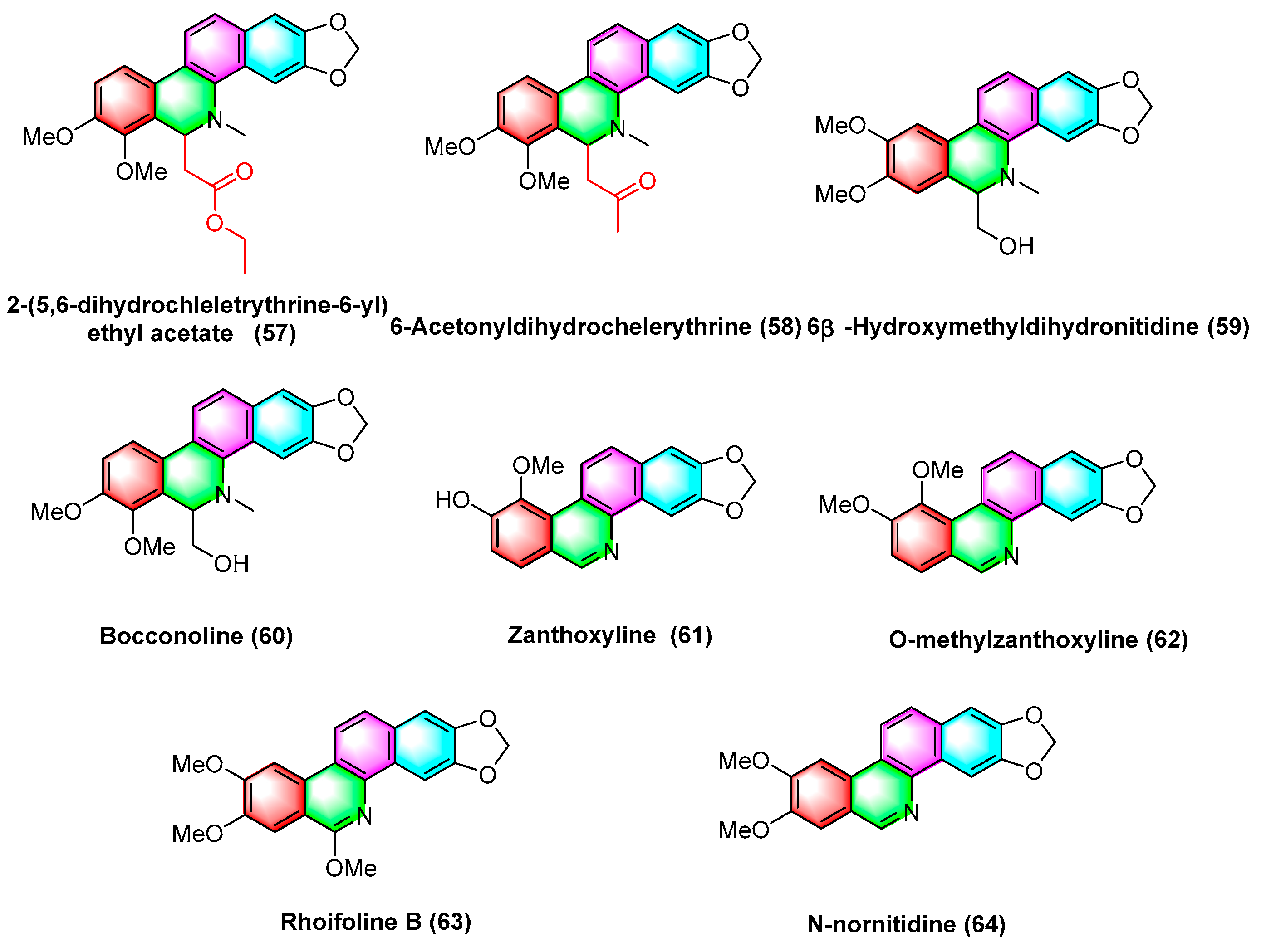
2.3. Antitumor Activities of Benzophenanthridine Alkaloids from Rutaceae
2.3.1. Antitumor Activities of Benzophenanthridine Alkaloids from Zanthoxylum rhoifolium
2.3.2. Antitumor Activities of Benzophenanthridine Alkaloids from Fagaropsis
2.3.3. Antitumor Activities of Benzopheridine Alkaloid from Toddalia
3. Conclusions and Perspectives
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Feng, S.; Chen, X.M.; Wang, J.F.; Xu, X.Q. 4032-4040-Th17 Cells Associated Cytokines and Cancer. Eur. Rev. Med. Pharmacol. Sci. 2016, 20, 4032–4040. [Google Scholar] [PubMed]
- Galadari, S.; Rahman, A.; Pallichankandy, S.; Thayyullathil, F. Molecular Targets and Anticancer Potential of Sanguinarine—A Benzophenanthridine Alkaloid. Phytomedicine 2017, 34, 143–153. [Google Scholar] [CrossRef] [PubMed]
- Singh, A.; Menéndez-Perdomo, I.M.; Facchini, P.J. Benzylisoquinoline Alkaloid Biosynthesis in Opium Poppy: An Update. Phytochem. Rev. 2019, 18, 1457–1482. [Google Scholar] [CrossRef]
- Fu, C.; Guan, G.; Wang, H. The Anticancer Effect of Sanguinarine: A Review. Curr. Pharm. Des. 2018, 24, 2760–2764. [Google Scholar] [CrossRef] [PubMed]
- Han, N.; Yang, Z.; Liu, Z.; Liu, H.; Yin, J. Research Progress on Natural Benzophenanthridine Alkaloids and Their Pharmacological Functions: A Review. Nat. Prod. Commun. 2016, 11, 1181–1188. [Google Scholar] [CrossRef]
- Alakkal, A.; Thayyullathil, F.; Pallichankandy, S.; Subburayan, K.; Cheratta, A.R.; Galadari, S. Sanguinarine Induces H2O2-Dependent Apoptosis and Ferroptosis in Human Cervical Cancer. Biomedicines 2022, 10, 1795. [Google Scholar] [CrossRef]
- Yang, J.; Fang, Z.; Wu, J.; Yin, X.; Fang, Y.; Zhao, F.; Zhu, S.; Li, Y. Construction and Application of a Lung Cancer Stem Cell Model: Antitumor Drug Screening and Molecular Mechanism of the Inhibitory Effects of Sanguinarine. Tumor Biol. 2016, 37, 13871–13883. [Google Scholar] [CrossRef]
- Singh, N.; Sharma, B. Toxicological Effects of Berberine and Sanguinarine. Front. Mol. Biosci. 2018, 5, 21. [Google Scholar] [CrossRef] [PubMed]
- Khan, A.Q.; Rashid, K.; AlAmodi, A.A.; Agha, M.V.; Akhtar, S.; Hakeem, I.; Raza, S.S.; Uddin, S. Reactive Oxygen Species (ROS) in Cancer Pathogenesis and Therapy: An Update on the Role of ROS in Anticancer Action of Benzophenanthridine Alkaloids. Biomed. Pharmacother. 2021, 143, 112142. [Google Scholar] [CrossRef]
- Wang, J.; Su, Q.; Wu, Q.; Chen, K.; Ullah, A.; Ghauri, M.A.; Zhang, Y. Sanguinarine Impairs Lysosomal Function and Induces ROS-Dependent Mitophagy and Apoptosis in Human Hepatocellular Carcinoma Cells. Arch. Pharm. Res. 2021, 44, 1025–1036. [Google Scholar] [CrossRef]
- Choi, W.Y.; Kim, G.-Y.; Lee, W.H.; Choi, Y.H. Sanguinarine, a Benzophenanthridine Alkaloid, Induces Apoptosis in MDA-MB-231 Human Breast Carcinoma Cells through a Reactive Oxygen Species-Mediated Mitochondrial Pathway. Chemotherapy 2008, 54, 279–287. [Google Scholar] [CrossRef] [PubMed]
- Su, Q.; Wang, J.; Wu, Q.; Ullah, A.; Ghauri, M.A.; Sarwar, A.; Chen, L.; Liu, F.; Zhang, Y. Sanguinarine Combats Hypoxia-Induced Activation of EphB4 and HIF-1α Pathways in Breast Cancer. Phytomedicine 2021, 84, 153503. [Google Scholar] [CrossRef] [PubMed]
- Ghauri, M.A.; Su, Q.; Ullah, A.; Wang, J.; Sarwar, A.; Wu, Q.; Zhang, D.; Zhang, Y. Sanguinarine Impedes Metastasis and Causes Inversion of Epithelial to Mesenchymal Transition in Breast Cancer. Phytomedicine 2021, 84, 153500. [Google Scholar] [CrossRef] [PubMed]
- Malla, R.R.; Bhamidipati, P.; Adem, M. Insights into the Potential of Sanguinarine as a Promising Therapeutic Option for Breast Cancer. Biochem. Pharmacol. 2023, 212, 115565. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Zhang, J.; Venkat, P.S.; Gu, C.; Meng, Y. Sanguinarine Exhibits Potent Efficacy against Cervical Cancer Cells through Inhibiting the STAT3 Pathway in Vitro and in Vivo. Cancer Manag. Res. 2019, 11, 7557–7566. [Google Scholar] [CrossRef] [PubMed]
- Ray, W.J.; Post, C.B.; Liu, Y.; Rhyu, G.I. Structural Changes at the Metal Ion Binding Site during the Phosphoglucomutase Reaction. Biochemistry 1993, 32, 48–57. [Google Scholar] [CrossRef]
- Saeed, M.E.M.; Mahmoud, N.; Sugimoto, Y.; Efferth, T.; Abdel-Aziz, H. Molecular Determinants of Sensitivity or Resistance of Cancer Cells Toward Sanguinarine. Front. Pharmacol. 2018, 9, 136. [Google Scholar] [CrossRef]
- Hałas-Wiśniewska, M.; Zielińska, W.; Izdebska, M.; Grzanka, A. The Synergistic Effect of Piperlongumine and Sanguinarine on the Non-Small Lung Cancer. Molecules 2020, 25, 3045. [Google Scholar] [CrossRef]
- Dong, X.-Z.; Song, Y.; Lu, Y.-P.; Hu, Y.; Liu, P.; Zhang, L. Sanguinarine Inhibits the Proliferation of BGC-823 Gastric Cancer Cells via Regulating MiR-96-5p/MiR-29c-3p and the MAPK/JNK Signaling Pathway. J. Nat. Med. 2019, 73, 777–788. [Google Scholar] [CrossRef]
- Rahman, A.; Pallichankandy, S.; Thayyullathil, F.; Galadari, S. Critical Role of H2O2 in Mediating Sanguinarine-Induced Apoptosis in Prostate Cancer Cells via Facilitating Ceramide Generation, ERK1/2 Phosphorylation, and Par-4 Cleavage. Free Radic. Biol. Med. 2019, 134, 527–544. [Google Scholar] [CrossRef]
- Yan, S.; Lin, S.; Chen, K.; Yin, S.; Peng, H.; Cai, N.; Ma, W.; Songyang, Z.; Huang, Y. Natural Product Library Screens Identify Sanguinarine Chloride as a Potent Inhibitor of Telomerase Expression and Activity. Cells 2022, 11, 1485. [Google Scholar] [CrossRef]
- Lee, J.S.; Jung, W.-K.; Jeong, M.H.; Yoon, T.R.; Kim, H.K. Sanguinarine Induces Apoptosis of HT-29 Human Colon Cancer Cells via the Regulation of Bax/Bcl-2 Ratio and Caspase-9-Dependent Pathway. Int. J. Toxicol. 2012, 31, 70–77. [Google Scholar] [CrossRef]
- Zhang, S.; Leng, T.; Zhang, Q.; Zhao, Q.; Nie, X.; Yang, L. Sanguinarine Inhibits Epithelial Ovarian Cancer Development via Regulating Long Non-Coding RNA CASC2-EIF4A3 Axis and/or Inhibiting NF-ΚB Signaling or PI3K/AKT/MTOR Pathway. Biomed. Pharmacother. 2018, 102, 302–308. [Google Scholar] [CrossRef]
- Sarkhosh-Inanlou, R.; Molaparast, M.; Mohammadzadeh, A.; Shafiei-Irannejad, V. Sanguinarine Enhances Cisplatin Sensitivity via Glutathione Depletion in Cisplatin-resistant Ovarian Cancer (A2780) Cells. Chem. Biol. Drug Des. 2020, 95, 215–223. [Google Scholar] [CrossRef] [PubMed]
- Och, A.; Zalewski, D.; Komsta, Ł.; Kołodziej, P.; Kocki, J.; Bogucka-Kocka, A. Cytotoxic and Proapoptotic Activity of Sanguinarine, Berberine, and Extracts of Chelidonium majus L. and Berberis thunbergii DC. toward Hematopoietic Cancer Cell Lines. Toxins 2019, 11, 485. [Google Scholar] [CrossRef]
- Wen, Y.; Song, Y.; Ma, Y.; Wen, J.; Yang, J. Sanguinarine Targets BRD4 to Suppress Cell Proliferation and Migration in Clear Cell Renal Cell Carcinoma. J. Biochem. Mol. Toxicol. 2023, 2, e23451. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.; Yu, W.; Shrivastava, A.; Alemi, F.; Lankachandra, K.; Srivastava, R.K.; Shankar, S. Sanguinarine Inhibits Pancreatic Cancer Stem Cell Characteristics by Inducing Oxidative Stress and Suppressing Sonic Hedgehog-Gli-Nanog Pathway. Carcinogenesis 2017, 38, 1047–1056. [Google Scholar] [CrossRef] [PubMed]
- Pica, F.; Balestrieri, E.; Serafino, A.; Sorrentino, R.; Gaziano, R.; Moroni, G.; Moroni, N.; Palmieri, G.; Mattei, M.; Garaci, E.; et al. Antitumor Effects of the Benzophenanthridine Alkaloid Sanguinarine in a Rat Syngeneic Model of Colorectal Cancer. Anticancer. Drugs 2012, 23, 32–42. [Google Scholar] [CrossRef] [PubMed]
- Kalogris, C.; Garulli, C.; Pietrella, L.; Gambini, V.; Pucciarelli, S.; Lucci, C.; Tilio, M.; Zabaleta, M.E.; Bartolacci, C.; Andreani, C.; et al. Sanguinarine Suppresses Basal-like Breast Cancer Growth through Dihydrofolate Reductase Inhibition. Biochem. Pharmacol. 2014, 90, 226–234. [Google Scholar] [CrossRef]
- De Stefano, I.; Raspaglio, G.; Zannoni, G.F.; Travaglia, D.; Prisco, M.G.; Mosca, M.; Ferlini, C.; Scambia, G.; Gallo, D. Antiproliferative and Antiangiogenic Effects of the Benzophenanthridine Alkaloid Sanguinarine in Melanoma. Biochem. Pharmacol. 2009, 78, 1374–1381. [Google Scholar] [CrossRef]
- Gaziano, R. Antitumor Effects of the Benzophenanthridine Alkaloid Sanguinarine: Evidence and Perspectives. World J. Gastrointest. Oncol. 2016, 8, 30. [Google Scholar] [CrossRef]
- Tian, Y.-Q.; Hu, D.; Zhang, Y.-L.; Zou, J.; Chen, G.-L.; Guo, M.-Q. Inhibitors Targeting Multiple Janus Kinases from Zanthoxylum Simulans Mediate Inhibition and Apoptosis against Gastric Cancer Cells via the Estrogen Pathway. Front. Chem. 2022, 10, 922110. [Google Scholar] [CrossRef]
- Zhu, M.; Niu, J.; Jiang, J.; Dong, T.; Chen, Y.; Yang, X.; Liu, P. Chelerythrine Inhibits the Progression of Glioblastoma by Suppressing the TGFB1-ERK1/2/Smad2/3-Snail/ZEB1 Signaling Pathway. Life Sci. 2022, 293, 120358. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.; Zheng, S.-Y.; Jiang, R.-L.; Wu, H.-D.; Li, Y.-A.; Lu, J.-L.; Xiong, Y.; Han, B.; Lin, L. Necroptosis Signaling and Mitochondrial Dysfunction Cross-Talking Facilitate Cell Death Mediated by Chelerythrine in Glioma. Free Radic. Biol. Med. 2023, 202, 76–96. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.; Pan, Y.; Zhang, G.; Wu, Y.; Zhong, W.; Chu, C.; Qian, Y.; Zhu, G. Chelerythrine Inhibits Human Hepatocellular Carcinoma Metastasis in Vitro. Biol. Pharm. Bull. 2018, 41, 36–46. [Google Scholar] [CrossRef]
- Wang, J.; Song, Y.; Zhang, N.; Li, N.; Liu, C.; Wang, B. Using Liposomes to Alleviate the Toxicity of Chelerythrine, a Natural PKC Inhibitor, in Treating Non-Small Cell Lung Cancer. Front. Oncol. 2021, 11, 658543. [Google Scholar] [CrossRef]
- Yang, B.; Zhang, D.; Qian, J.; Cheng, Y. Chelerythrine Suppresses Proliferation and Metastasis of Human Prostate Cancer Cells via Modulating MMP/TIMP/NF-ΚB System. Mol. Cell. Biochem. 2020, 474, 199–208. [Google Scholar] [CrossRef]
- Zhang, G.; Zhu, L.; Xue, Y.; Zhao, Z.; Li, H.; Niu, Z.; Wang, X.; Chen, P.; Zhang, J.; Zhang, X. Benzophenanthridine Alkaloids Suppress Lung Adenocarcinoma by Blocking TMEM16A Ca2+-Activated Cl− Channels. Pflüg. Arch. Eur. J. Physiol. 2020, 472, 1457–1467. [Google Scholar] [CrossRef]
- He, H.; Zhuo, R.; Dai, J.; Wang, X.; Huang, X.; Wang, H.; Xu, D. Chelerythrine Induces Apoptosis via ROS-mediated Endoplasmic Reticulum Stress and STAT3 Pathways in Human Renal Cell Carcinoma. J. Cell. Mol. Med. 2020, 24, 50–60. [Google Scholar] [CrossRef]
- Cho, O.; Lee, J.-W.; Kim, H.-S.; Jeong, Y.-J.; Heo, T.-H. Chelerythrine, a Novel Small Molecule Targeting IL-2, Inhibits Melanoma Progression by Blocking the Interaction between IL-2 and Its Receptor. Life Sci. 2023, 320, 121559. [Google Scholar] [CrossRef]
- Liang, D.; Liu, L.; Zheng, Q.; Zhao, M.; Zhang, G.; Tang, S.; Tang, J.; Chen, N. Chelerythrine Chloride Inhibits the Progression of Colorectal Cancer by Targeting Cancer-associated Fibroblasts through Intervention with WNT10B/B-catenin and TGFB2/Smad2/3 Axis. Phytother. Res. 2023. Online ahead of print. [Google Scholar] [CrossRef] [PubMed]
- Heng, W.S.; Cheah, S.-C. Chelerythrine Chloride Downregulates β-Catenin and Inhibits Stem Cell Properties of Non-Small Cell Lung Carcinoma. Molecules 2020, 25, 224. [Google Scholar] [CrossRef]
- Almeida, I.V.; Fernandes, L.M.; Biazi, B.I.; Vicentini, V.E.P. Evaluation of the Anticancer Activities of the Plant Alkaloids Sanguinarine and Chelerythrine in Human Breast Adenocarcinoma Cells. Anticancer. Agents Med. Chem. 2017, 17, 1586–1592. [Google Scholar] [CrossRef] [PubMed]
- Bhat, J.; Chatterjee, S. Skeleton Selectivity in Complexation of Chelerythrine and Chelerythrine-like Natural Plant Alkaloids with the G-Quadruplex Formed at the Promoter of c-MYC Oncogene: In Silico Exploration. RSC Adv. 2016, 6, 36667–36680. [Google Scholar] [CrossRef]
- Romo-Pérez, A.; Miranda, L.D.; Chávez-Blanco, A.D.; Dueñas-González, A.; Camacho-Corona, M.d.R.; Acosta-Huerta, A.; García, A. Mild C(Sp)–H Functionalization of Dihydrosanguinarine and Dihydrochelerythrine for Development of Highly Cytotoxic Derivatives. Eur. J. Med. Chem. 2017, 138, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Tang, Y.; Xu, X.; Li, J.; Deng, L.; Mu, S. Synthesis and Antileukemia Activity Evaluation of Benzophenanthridine Alkaloid Derivatives. Molecules 2022, 27, 3934. [Google Scholar] [CrossRef] [PubMed]
- Slaninová, I.; Pěnčíková, K.; Urbanová, J.; Slanina, J.; Táborská, E. Antitumour Activities of Sanguinarine and Related Alkaloids. Phytochem. Rev. 2014, 13, 51–68. [Google Scholar] [CrossRef]
- Slaninová, I.; Slunská, Z.; Šinkora, J.; Vlková, M.; Táborská, E. Screening of Minor Benzo(c.)Phenanthridine Alkaloids for Antiproliferative and Apoptotic Activities. Pharm. Biol. 2007, 45, 131–139. [Google Scholar] [CrossRef]
- Slunská, Z.; Gelnarová, E.; Hammerová, J.; Táborská, E.; Slaninová, I. Effect of Quaternary Benzo[c]Phenanthridine Alkaloids Sanguilutine and Chelilutine on Normal and Cancer Cells. Toxicol. Vitr. 2010, 24, 697–706. [Google Scholar] [CrossRef]
- Hammerová, J.; Uldrijan, S.; Táborská, E.; Vaculová, A.H.; Slaninová, I. Necroptosis Modulated by Autophagy Is a Predominant Form of Melanoma Cell Death Induced by Sanguilutine. Biol. Chem. 2012, 393, 647–658. [Google Scholar] [CrossRef]
- Guo, Y.; Zhao, L.; Chang, B.; Yu, J.; Bao, J.; Yao, Q.; Luo, J. The Traditional Uses, Phytochemistry, Pharmacokinetics, Pharmacology, Toxicity, and Applications of Corydalis Saxicola Bunting: A Review. Front. Pharmacol. 2022, 13, 822792. [Google Scholar] [CrossRef] [PubMed]
- Qin, F.; Dai, L.; Zhang, B.; Huang, R.; Wang, F.; Qin, J.; Liang, D.; Wang, H. (±)-Corysaxicolaine A: A Pair of Antitumor Enantiomeric Alkaloid Dimers from Corydalis Saxicola. Org. Biomol. Chem. 2022, 20, 1396–1400. [Google Scholar] [CrossRef] [PubMed]
- Kazemi Noureini, S.; Fatemi, L.; Wink, M. Telomere Shortening in Breast Cancer Cells (MCF7) under Treatment with Low Doses of the Benzylisoquinoline Alkaloid Chelidonine. PLoS ONE 2018, 13, e0204901. [Google Scholar] [CrossRef] [PubMed]
- Qian, L.; Chen, K.; Wang, C.; Chen, Z.; Meng, Z.; Wang, P. Targeting NRAS-Mutant Cancers with the Selective STK19 Kinase Inhibitor Chelidonine. Clin. Cancer Res. 2020, 26, 3408–3419. [Google Scholar] [CrossRef]
- Csomós, I.; Nagy, P.; Filep, C.; Rebenku, I.; Nizsalóczki, E.; Kovács, T.; Vámosi, G.; Mátyus, L.; Bodnár, A. Opposing Effects of Chelidonine on Tyrosine and Serine Phosphorylation of STAT3 in Human Uveal Melanoma Cells. Int. J. Mol. Sci. 2021, 22, 12974. [Google Scholar] [CrossRef] [PubMed]
- Herrmann, R.; Roller, J.; Polednik, C.; Schmidt, M. Effect of Chelidonine on Growth, Invasion, Angiogenesis and Gene Expression in Head and Neck Cancer Cell Lines. Oncol. Lett. 2018, 16, 3108–3116. [Google Scholar] [CrossRef]
- Havelek, R.; Seifrtova, M.; Kralovec, K.; Krocova, E.; Tejkalova, V.; Novotny, I.; Cahlikova, L.; Safratova, M.; Opletal, L.; Bilkova, Z.; et al. Comparative Cytotoxicity of Chelidonine and Homochelidonine, the Dimethoxy Analogues Isolated from Chelidonium majus L. (Papaveraceae), against Human Leukemic and Lung Carcinoma Cells. Phytomedicine 2016, 23, 253–266. [Google Scholar] [CrossRef]
- Xie, Y.-J.; Gao, W.-N.; Wu, Q.-B.; Yao, X.-J.; Jiang, Z.-B.; Wang, Y.-W.; Wang, W.-J.; Li, W.; Hussain, S.; Liu, L.; et al. Chelidonine Selectively Inhibits the Growth of Gefitinib-Resistant Non-Small Cell Lung Cancer Cells through the EGFR-AMPK Pathway. Pharmacol. Res. 2020, 159, 104934. [Google Scholar] [CrossRef]
- Lee, Y.-K.; Lee, K.W.; Kim, M.; Lee, Y.; Yoo, J.; Hwangbo, C.; Park, K.H.; Kim, K.D. Chelidonine Induces Caspase-Dependent and Caspase-Independent Cell Death through G 2/M Arrest in the T98G Human Glioblastoma Cell Line. Evid. Based Complement. Alternat. Med. 2019, 2019, 6318179. [Google Scholar] [CrossRef]
- Hou, F.; Guo, L.; Zheng, K.; Song, J.; Wang, Q.; Zheng, Y. Chelidonine Enhances the Antitumor Effect of Lenvatinib on Hepatocellular Carcinoma Cells. OncoTargets Ther. 2019, 12, 6685–6697. [Google Scholar] [CrossRef]
- Havelek, R.; Seifrtova, M.; Kralovec, K.; Habartova, K.; Cahlikova, L.; Rezacova, M. Chelidonine and Homochelidonine Induce Cell Death through Cell Cycle Checkpoints and MAP Kinase Pathways. Nat. Prod. Commun. 2017, 12, 1934578X1701200910. [Google Scholar] [CrossRef]
- Yi, C.; Li, X.; Chen, S.; Liu, M.; Lu, W.; Ye, X. Natural Product Corynoline Suppresses Melanoma Cell Growth through Inducing Oxidative Stress. Phytother. Res. 2020, 34, 2766–2777. [Google Scholar] [CrossRef]
- Chen, J.-J.; Duh, C.-Y.; Chen, I.-S. New Tetrahydroprotoberberine N-Oxide Alkaloids and Cytotoxic Constituents of Corydalis Tashiroi. Planta Med. 1999, 65, 643–647. [Google Scholar] [CrossRef]
- Gao, C.; Gu, X.; Wang, X.; Cao, H.; Lin, B.; Liu, Y.; Di, X. Corygaline A, a Hexahydrobenzophenanthridine Alkaloid with an Unusual Carbon Skeleton from Corydalis Bungeana Turcz. Org. Biomol. Chem. 2018, 16, 8710–8714. [Google Scholar] [CrossRef]
- Li, J.; Yan, Z.; Li, H.; Shi, Q.; Huang, L.; Nimishetti, N.; Allen, T.D.; Yang, D.; Zhang, J. A High-Content Screen for Anti-Mitosis and Polyploidy-Induction Identifies an Unknown Activity of Two Benzophenanthridine Alkaloids from Corydalis longicalcarata. Phytochem. Lett. 2021, 41, 180–185. [Google Scholar] [CrossRef]
- Liu, Z.; Mi, Z.; Wang, P.; Chang, S.; Han, N.; Yin, J. Two New Alkaloids from the Tubers of Corydalis Ambigua Subsp. Amurensis and Their Anti-Proliferative Activity. Nat. Prod. Res. 2020, 34, 3305–3312. [Google Scholar] [CrossRef]
- Zou, H.-L.; Li, H.-Y.; Liu, B.-L.; Zhou, G.-X. A New Cytotoxic Benzophenanthridine Isoquinoline Alkaloid from the Fruits of Macleaya cordata. J. Asian Nat. Prod. Res. 2015, 17, 856–860. [Google Scholar] [CrossRef]
- Si, Y.; Wang, J.; Liu, X.; Zhou, T.; Xiang, Y.; Zhang, T.; Wang, X.; Feng, T.; Xu, L.; Yu, Q.; et al. Ethoxysanguinarine, a Novel Direct Activator of AMP-Activated Protein Kinase, Induces Autophagy and Exhibits Therapeutic Potential in Breast Cancer Cells. Front. Pharmacol. 2020, 10, 1503. [Google Scholar] [CrossRef]
- Ma, L.; Xuan, X.; Chen, X.; Fan, M.; Liu, J.; Huang, G.; Liu, Z. Ethoxysanguinarine Induces Apoptosis, Inhibits Metastasis and Sensitizes Cells to Docetaxel in Breast Cancer Cells through Inhibition of Hakai. Chem. Biodivers. 2023, 20, e202200284. [Google Scholar] [CrossRef]
- Deng, A.; Qin, H. Cytotoxic Dihydrobenzophenanthridine Alkaloids from the Roots of Macleaya microcarpa. Phytochemistry 2010, 71, 816–822. [Google Scholar] [CrossRef]
- Sai, C.-M.; Li, D.-H.; Xue, C.-M.; Wang, K.-B.; Hu, P.; Pei, Y.-H.; Bai, J.; Jing, Y.-K.; Li, Z.-L.; Hua, H.-M. Two Pairs of Enantiomeric Alkaloid Dimers from Macleaya Cordata. Org. Lett. 2015, 17, 4102–4105. [Google Scholar] [CrossRef]
- Sai, C.-M.; Li, D.-H.; Li, S.-G.; Han, T.; Guo, Y.-Z.; Pei, Y.-H.; Bai, J.; Jing, Y.-K.; Li, Z.-L.; Hua, H.-M. Racemic Alkaloids from Macleaya cordata: Structural Elucidation, Chiral Resolution, and Cytotoxic, Antibacterial Activities. RSC Adv. 2016, 6, 41173–41180. [Google Scholar] [CrossRef]
- Phillips, S.D.; Castle, R.N. A Review of the Chemistry of the Antitumor Benzo[c]Phenanthridine Alkaloids Nitidine and Fagaronine and of the Related Antitumor Alkaloid Coralyne. J. Heterocycl. Chem. 1981, 18, 223–232. [Google Scholar] [CrossRef]
- Jiang, J.; Ma, H.; Hu, S.; Cheng, L.; Wang, F.; Zhang, G. Benzophenanthridine Alkaloids from the Roots of Thalictrum Microgynum Lecoy.Ex Oliv. Nat. Prod. Res. 2019, 33, 2964–2969. [Google Scholar] [CrossRef]
- Wu, S.; Xu, H.; Wu, X.; Liu, P.; Shi, Y.; Pang, P.; Deng, L.; Zhou, G.; Chen, X. Dihydrosanguinarine Suppresses Pancreatic Cancer Cells via Regulation of Mut-P53/WT-P53 and the Ras/Raf/Mek/Erk Pathway. Phytomedicine 2019, 59, 152895. [Google Scholar] [CrossRef]
- Vrba, J.; Doležel, P.; Vičar, J.; Ulrichová, J. Cytotoxic Activity of Sanguinarine and Dihydrosanguinarine in Human Promyelocytic Leukemia HL-60 Cells. Toxicol. Vitr. 2009, 23, 580–588. [Google Scholar] [CrossRef]
- Qin, S.-Q.; Li, L.-C.; Song, J.-R.; Li, H.-Y.; Li, D.-P. Structurally Simple Phenanthridine Analogues Based on Nitidine and Their Antitumor Activities. Molecules 2019, 24, 437. [Google Scholar] [CrossRef]
- Pan, X.; Han, H.; Wang, L.; Yang, L.; Li, R.; Li, Z.; Liu, J.; Zhao, Q.; Qian, M.; Liu, M.; et al. Nitidine Chloride Inhibits Breast Cancer Cells Migration and Invasion by Suppressing C-Src/FAK Associated Signaling Pathway. Cancer Lett. 2011, 313, 181–191. [Google Scholar] [CrossRef]
- Lin, J.; Shen, A.; Chen, H.; Liao, J.; Xu, T.; Liu, L.; Lin, J.; Peng, J. Nitidine Chloride Inhibits Hepatic Cancer Growth via Modulation of Multiple Signaling Pathways. BMC Cancer 2014, 14, 729. [Google Scholar] [CrossRef]
- Mou, H.; Guo, P.; Li, X.; Zhang, C.; Jiang, J.; Wang, L.; Wang, Q.; Yuan, Z. Nitidine Chloride Inhibited the Expression of S Phase Kinase-Associated Protein 2 in Ovarian Cancer Cells. Cell Cycle 2017, 16, 1366–1375. [Google Scholar] [CrossRef]
- Fang, Z.; Tang, Y.; Jiao, W.; Xing, Z.; Guo, Z.; Wang, W.; Xu, Z.; Liu, Z. Nitidine Chloride Induces Apoptosis and Inhibits Tumor Cell Proliferation via Suppressing ERK Signaling Pathway in Renal Cancer. Food Chem. Toxicol. 2014, 66, 210–216. [Google Scholar] [CrossRef]
- Lian, C.; Huang, Y.; Hu, P.; Cao, Y.; Zhang, Z.; Feng, F.; Zhang, J. Nitidine Chloride Triggers Autophagy and Apoptosis of Ovarian Cancer Cellsthrough Akt/MTOR Signaling Pathway. Curr. Pharm. Des. 2023, 29, 1524–1534. [Google Scholar] [CrossRef]
- Wang, L.; Hou, Y.; Yin, X.; Su, J.; Zhao, Z.; Ye, X.; Zhou, X.; Zhou, L.; Wang, Z. Rottlerin Inhibits Cell Growth and Invasion via Down-Regulation of Cdc20 in Glioma Cells. Oncotarget 2016, 7, 69770–69782. [Google Scholar] [CrossRef]
- Xu, H.; Cao, T.; Zhang, X.; Shi, Y.; Zhang, Q.; Chai, S.; Yu, L.; Jin, G.; Ma, J.; Wang, P.; et al. Nitidine Chloride Inhibits SIN1 Expression in Osteosarcoma Cells. Mol. Ther. Oncolytics 2019, 12, 224–234. [Google Scholar] [CrossRef]
- Zhai, H.; Hu, S.; Liu, T.; Wang, F.; Wang, X.; Wu, G.; Zhang, Y.; Sui, M.; Liu, H.; Jiang, L. Nitidine Chloride Inhibits Proliferation and Induces Apoptosis in Colorectal Cancer Cells by Suppressing the ERK Signaling Pathway. Mol. Med. Rep. 2016, 13, 2536–2542. [Google Scholar] [CrossRef]
- Yang, I.; Jung, W.; Kim, L.; Shin, J.; Cho, N.; Hong, S.D.; Hong, K.; Cho, S. Nitidine Chloride Represses Mcl-1 Protein via Lysosomal Degradation in Oral Squamous Cell Carcinoma. J. Oral Pathol. Med. 2018, 47, 823–829. [Google Scholar] [CrossRef]
- Li, P.; Yan, S.; Dong, X.; Li, Z.; Qiu, Y.; Ji, C.; Zhang, J.; Ji, M.; Li, W.; Wang, H.; et al. Cell Cycle Arrest and Apoptosis Induction Activity of Nitidine Chloride on Acute Myeloid Leukemia Cells. Med. Chem. 2018, 14, 60–66. [Google Scholar] [CrossRef]
- Liu, N.; Li, P.; Zang, S.; Liu, Q.; Ma, D.; Sun, X.; Ji, C. Novel Agent Nitidine Chloride Induces Erythroid Differentiation and Apoptosis in CML Cells through C-Myc-MiRNAs Axis. PLoS ONE 2015, 10, e0116880. [Google Scholar] [CrossRef]
- Wang, C.-F.; Fan, L.; Tian, M.; Du, S.-S.; Deng, Z.-W.; Feng, J.-B.; Wang, Y.-Y.; Su, X. Cytotoxicity of Benzophenanthridine Alkaloids from the Roots of Zanthoxylum Nitidum (Roxb.) DC. Var. Fastuosum How Ex Huang. Nat. Prod. Res. 2015, 29, 1380–1383. [Google Scholar] [CrossRef]
- Jia, M.; Wang, Y.; Guo, Y.; Yu, P.; Sun, Y.; Song, Y.; Zhao, L. Nitidine Chloride Suppresses Epithelial-mesenchymal Transition and Stem Cell-like Properties in Glioblastoma by Regulating JAK2/STAT3 Signaling. Cancer Med. 2021, 10, 3113–3128. [Google Scholar] [CrossRef]
- Li, D.; Zhang, Q.; Zhou, Y.; Zhu, H.; Li, T.; Du, F. A Novel Nitidine Chloride Nanoparticle Overcomes the Stemness of CD133+EPCAM+ Huh7 Hepatocellular Carcinoma Cells for Liver Cancer Therapy. BMC Pharmacol. Toxicol. 2022, 23, 48. [Google Scholar] [CrossRef]
- Sun, M.; Zhang, N.; Wang, X.; Li, Y.; Qi, W.; Zhang, H.; Li, Z.; Yang, Q. Hedgehog Pathway Is Involved in Nitidine Chloride Induced Inhibition of Epithelial-Mesenchymal Transition and Cancer Stem Cells-like Properties in Breast Cancer Cells. Cell Biosci. 2016, 6, 44. [Google Scholar] [CrossRef]
- Wang, S.; Zhai, G.; Huang, Z.; Luo, J.; He, J.; Huang, J.; Yang, L.; Xiao, C.; Li, S.; Chen, K.; et al. Nitidine Chloride Regulates Cell Function of Bladder Cancer in Vitro through Downregulating Lymphocyte Antigen 75. Naunyn. Schmiedebergs Arch. Pharmacol. 2023, 396, 2071–2085. [Google Scholar] [CrossRef]
- Sandjo, L.P.; Kuete, V.; Tchangna, R.S.; Efferth, T.; Ngadjui, B.T. Cytotoxic Benzophenanthridine and Furoquinoline Alkaloids from Zanthoxylum buesgenii (Rutaceae). Chem. Cent. J. 2014, 8, 61. [Google Scholar] [CrossRef]
- Deng, Y.; Ding, T.; Deng, L.; Hao, X.; Mu, S. Active Constituents of Zanthoxylum nitidium from Yunnan Province against Leukaemia Cells in Vitro. BMC Chem. 2021, 15, 44. [Google Scholar] [CrossRef]
- Yang, H.; Wang, F.-T.; Wu, M.; Wang, W.; Agama, K.; Pommier, Y.; An, L.-K. Synthesis of 11-Aminoalkoxy Substituted Benzophenanthridine Derivatives as Tyrosyl-DNA Phosphodiesterase 1 Inhibitors and Their Anticancer Activity. Bioorganic Chem. 2022, 123, 105789. [Google Scholar] [CrossRef]
- Kaigongi, M.M.; Lukhoba, C.W.; Yaouba, S.; Makunga, N.P.; Githiomi, J.; Yenesew, A. In Vitro Antimicrobial and Antiproliferative Activities of the Root Bark Extract and Isolated Chemical Constituents of Zanthoxylum paracanthum Kokwaro (Rutaceae). Plants 2020, 9, 920. [Google Scholar] [CrossRef]
- Nguyen, T.H.V.; Tran, T.T.; Cam, T.I.; Pham, M.Q.; Pham, Q.L.; Vu, D.H.; Nguyen, X.N.; Chau, V.M.; Van, K.P. Alkaloids From Zanthoxylum Nitidum and Their Cytotoxic Activity. Nat. Prod. Commun. 2019, 14, 1934578X1984413. [Google Scholar] [CrossRef]
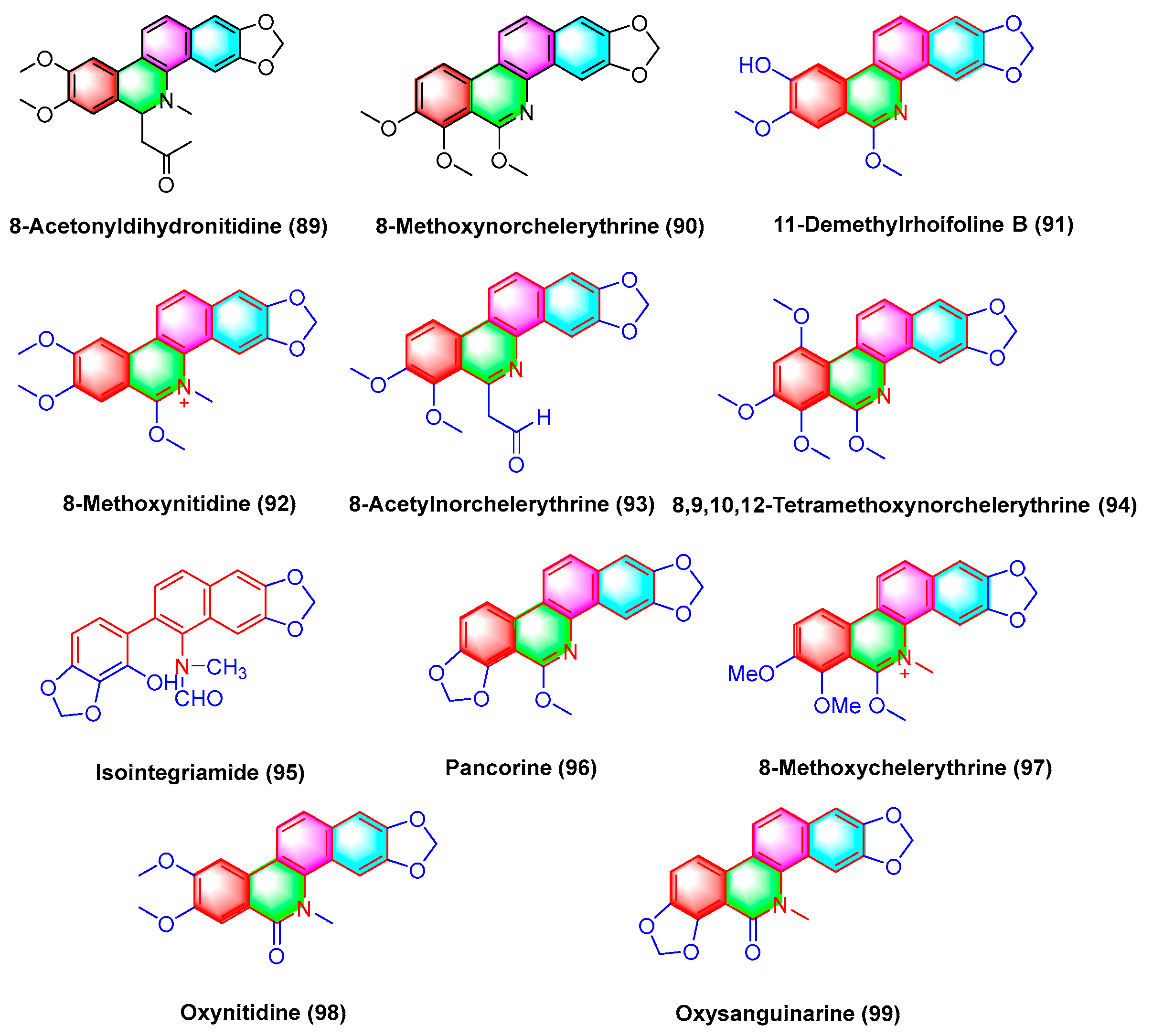
- Iwasaki, H.; Oku, H.; Takara, R.; Miyahira, H.; Hanashiro, K.; Yoshida, Y.; Kamada, Y.; Toyokawa, T.; Takara, K.; Inafuku, M. The Tumor Specific Cytotoxicity of Dihydronitidine from Toddalia asiatica Lam. Cancer Chemother. Pharmacol. 2006, 58, 451–459. [Google Scholar] [CrossRef]
- Chen, J.-J.; Fang, H.-Y.; Duh, C.-Y.; Chen, I.-S. New Indolopyridoquinazoline, Benzo[c]phenanthridines and Cytotoxic Constituents from Zanthoxylum integrifoliolum. Planta Med. 2005, 71, 470–475. [Google Scholar] [CrossRef]
- Phetkul, U.; Hayiawae, N.; Khunthong, S.; Daus, M.; Voravuthikunchai, S.P.; Tamvapee, P.; Watanapokasin, R.; Chakthong, S. Zanthoisobutylamides A–C: Rare Dimeric C-6 Substituent Dihydrobenzophenanthridine Alkaloids from the Roots of Zanthoxylum Nitidum. Nat. Prod. Res. 2021, 37, 1249–1257. [Google Scholar] [CrossRef] [PubMed]
- Wangensteen, H.; Ho, G.T.T.; Tadesse, M.; Miles, C.O.; Moussavi, N.; Mikolo, B.; Malterud, K.E. A New Benzophenanthridine Alkaloid and Other Bioactive Constituents from the Stem Bark of Zanthoxylum heitzii. Fitoterapia 2016, 109, 196–200. [Google Scholar] [CrossRef]
- Mbaveng, A.T.; Damen, F.; Çelik, İ.; Tane, P.; Kuete, V.; Efferth, T. Cytotoxicity of the Crude Extract and Constituents of the Bark of Fagara tessmannii towards Multi-Factorial Drug Resistant Cancer Cells. J. Ethnopharmacol. 2019, 235, 28–37. [Google Scholar] [CrossRef]
- Nakanishi, T.; Masuda, A.; Suwa, M.; Akiyama, Y.; Hoshino-Abe, N.; Suzuki, M. Synthesis of Derivatives of NK109, 7-OH Benzo[c]Phenanthridine Alkaloid, and Evaluation of Their Cytotoxicities and Reduction-Resistant Properties. Bioorg. Med. Chem. Lett. 2000, 10, 2321–2323. [Google Scholar] [CrossRef] [PubMed]
- Nakanishi, T.; Suzuki, M.; Saimoto, A.; Kabasawa, T. Structural Considerations of NK109, an Antitumor Benzo[c]Phenanthridine Alkaloid. J. Nat. Prod. 1999, 62, 864–867. [Google Scholar] [CrossRef]
- Kanzawa, F.; Nishio, K.; Ishida, T.; Fukuda, M.; Kurokawa, H.; Fukumoto, H.; Nomoto, Y.; Fukuoka, K.; Bojanowski, K.; Saijo, N. Anti-Tumour Activities of a New Benzo[c]Phenanthridine Agent, 2,3-(Methylenedioxy)-5-Methyl-7-Hydroxy-8-Methoxybenzo[c]Phenanthridinium Hydrogensulphate Dihydrate (NK109), against Several Drug-Resistant Human Tumour Cell Lines. Br. J. Cancer 1997, 76, 571–581. [Google Scholar] [CrossRef]
- Cho, W.-J.; Yoo, S.-J.; Chung, B.-H.; Choi, B.-G.; Cheon, S.H.; Whang, S.-H.; Kim, S.-K.; Kang, B.-H.; Lee, C.-O. Synthesis of Benzo[c]Phenanthridine Derivatives and Theirin Vitro Antitumor Activities. Arch. Pharm. Res. 1996, 19, 321–325. [Google Scholar] [CrossRef]
- Zhou, J.; Li, Z.; Zhang, J.; Wang, H.; Yin, S.; Du, J. 8-Acetonyldihydronitidine Inhibits the Proliferation of Human Colorectal Cancer Cells via Activation of P53. Eur. J. Pharmacol. 2019, 854, 256–264. [Google Scholar] [CrossRef]
- Hu, J.; Shi, X.; Chen, J.; Mao, X.; Zhu, L.; Yu, L.; Shi, J. Alkaloids from Toddalia asiatica and Their Cytotoxic, Antimicrobial and Antifungal Activities. Food Chem. 2014, 148, 437–444. [Google Scholar] [CrossRef]
- Tang, W.-L.; Zhang, Y.; Hu, D.-X.; Yang, H.; Yu, Q.; Chen, J.-W.; Agama, K.; Pommier, Y.; An, L.-K. Synthesis and Biological Evaluation of 5-Aminoethyl Benzophenanthridone Derivatives as DNA Topoisomerase IB Inhibitors. Eur. J. Med. Chem. 2019, 178, 81–92. [Google Scholar] [CrossRef]
- Cosialls, E.; El Hage, R.; Dos Santos, L.; Gong, C.; Mehrpour, M.; Hamaï, A. Ferroptosis: Cancer Stem Cells Rely on Iron until “to Die for” It. Cells 2021, 10, 2981. [Google Scholar] [CrossRef] [PubMed]






























{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
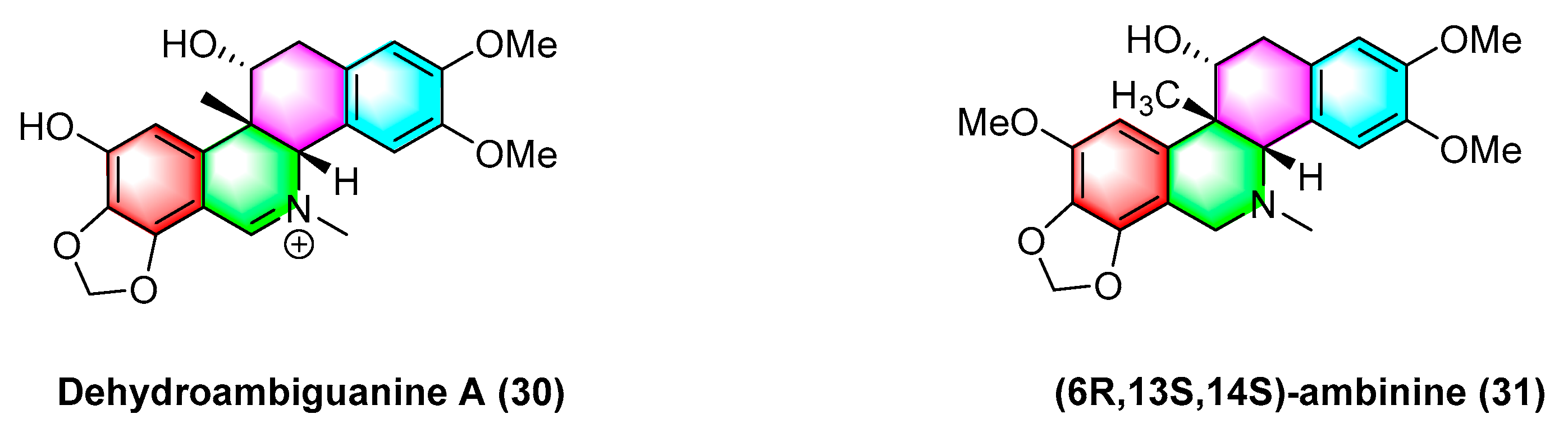{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound | R | IC50 Value | |
|---|---|---|---|
| Jurkat Clone e6-1 | THP-1 | ||
| Sanguinarine | / | 1.56 ± 0.09 | 1.60 ± 0.13 |
| Compound 8 |  | 1.30 ± 0.05 | 1.46 ± 0.06 |
| Compound 9 |  | 0.52 ± 0.03 | 0.48 ± 0.03 |
| Compound 10 |  | 1.23 ± 0.08 | 1.38 ± 0.04 |
| Compound 11 |  | 0.91 ± 0.04 | 1.17 ± 0.13 |
| Compound 12 |  | 0.53 ± 0.05 | 0.18 ± 0.03 |
| Compound | R | IC50 Value | |
|---|---|---|---|
| Jurkat Clone e6-1 | THP-1 | ||
| Chelerythrine | / | 5.58 ± 0.13 | 4.70 ± 0.07 |
| Compound 13 |  | 7.94 ± 0.10 | 5.78 ± 0.23 |
| Compound 14 |  | 2.61 ± 0.19 | 1.87 ± 0.22 |
| Compound 15 |  | 2.48 ± 0.13 | 4.45 ± 0.34 |
| Compound 16 |  | 5.64 ± 0.20 | 5.88 ± 0.07 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Peng, R.; Xu, M.; Xie, B.; Min, Q.; Hui, S.; Du, Z.; Liu, Y.; Yu, W.; Wang, S.; Chen, X.; et al. Insights on Antitumor Activity and Mechanism of Natural Benzophenanthridine Alkaloids. Molecules 2023, 28, 6588. https://doi.org/10.3390/molecules28186588
Peng R, Xu M, Xie B, Min Q, Hui S, Du Z, Liu Y, Yu W, Wang S, Chen X, et al. Insights on Antitumor Activity and Mechanism of Natural Benzophenanthridine Alkaloids. Molecules. 2023; 28(18):6588. https://doi.org/10.3390/molecules28186588
Chicago/Turabian StylePeng, Rui, Mengwei Xu, Baocheng Xie, Qing Min, Siwen Hui, Ziwei Du, Yan Liu, Wei Yu, Shi Wang, Xin Chen, and et al. 2023. "Insights on Antitumor Activity and Mechanism of Natural Benzophenanthridine Alkaloids" Molecules 28, no. 18: 6588. https://doi.org/10.3390/molecules28186588