

Traditional Uses, Pharmacology and Phytochemistry of the Medicinal Plant Flueggea virosa (Roxb. ex Willd.) Royle

1
CNRS, Inserm, CHU Lille, UMR9020-U1277-CANTHER—Cancer Heterogeneity Plasticity and Resistance to Therapies, OncoLille Institut, University of Lille, F-59000 Lille, France
2
Institute of Pharmaceutical Chemistry Albert Lespagnol (ICPAL), Faculty of Pharmacy, University of Lille, F-59006 Lille, France
3
OncoWitan, Scientific Consulting Office, F-59290 Lille, France
Future Pharmacol. 2024, 4(1), 77-102; https://doi.org/10.3390/futurepharmacol4010007
Submission received: 4 November 2023
/
Revised: 15 December 2023
/
Accepted: 16 January 2024
/
Published: 18 January 2024
Abstract
:The white berry bush, officially Flueggea virosa (Roxb. ex Willd.) Royle is a medicinal plant distributed throughout tropical areas and traditionally used in Africa, India and China. Root decoctions are used to treat abdominal pain, whereas extracts from the aerial parts serve to treat liver and urinary diseases, inflammatory pathologies and diabetes, among other pathologies. Plant extracts have revealed antiparasitic, antimicrobial, antiepilepsy, antidiabetic, anticancer and analgesic effects. Three main categories of phytochemicals were isolated from F. virosa: polyphenols, with the lead product bergenin; terpenoids, such as the flueggenoids and related podocarpane-type diterpenoids; and many alkaloids derived from securinine and norsecurinine. A remarkable feature of S. virosa is the production of norsecurinine oligomers, including macromolecular tetramers and pentamers, such as fluevirosinines. The most potent anticancer alkaloid in the family is the dimeric indolizidine flueggine B, which was identified as a potential binder to α/β-tubulin dimer, which is a known target for securinine. This review highlights the diversity of phytochemicals identified from S. virosa and the potential therapeutic benefits of dimeric alkaloids. Studies are encouraged to further investigate the therapeutic properties of the lead compounds but also define and finesse the nutritional profile of the edible fruit.
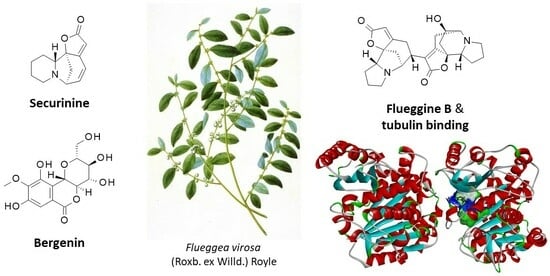
1. Introduction
The plant genus Flueggea (family Phyllanthaceae, previously called Euphorbiaceae) includes 16 species with a botanically accepted name (Table 1). These plants have a world-wide distribution but they are particularly abundant in tropical areas, ranging from Africa to Asia and Australia [1]. They are mostly shrubs and trees and are usually referred to as bushweeds. The plant genus Flueggea was named in honor of the botanist Johannes Flüggé (1775–1816), who was a native of Hamburg (Germany) and the author of the famous monograph on Paspalum plants (Graminum Monographiae published in 1810).
Several Flueggea species are used in traditional medicine. This is the case for F. suffruticosa (Pall.) Baill., which is commonly used in traditional Chinese medicine for the treatment of inflammatory ailments, such as rheumatism and lumbago [2], and for F. leucopyrus Willd., which is used by folk medical practitioners in Sri Lanka as a decoction to treat cancer [3,4]. Another largely used species is Flueggea virosa (Roxb. ex Willd.) Royle (Figure 1; hereafter F. virosa) as a leaf decoction to cure lactation problems and sick babies at birth and as a root decoction to cure abdominal pain and other applications. The plant is used to treat various diseases and symptoms in Asia, notably as a folk Chinese medicine for the treatment of eczema and rheumatoid arthritis [5,6]. It is also an important medicinal plant in tropical Africa, used alone or in combination with other plants, for a variety of purposes, including liver, kidney, urinary and venereal diseases, bile deficiency, testicular inflammation, frigidity, sterility, heavy menstruation, rheumatism and arthritis [7]. The plant has been around for a long time. There is evidence of the use of a digging stick of F. virosa dated to ∼39,000 years before present by the Stone Age inhabitants in South Africa [8].
F. virosa is known as the white berry bush or the Chinese waterberry in English. The given common names vary considerably from one country to another, with more than 50 trivial names identified in >35 countries (Table 2). The diversity of names reflects the common use of the plant in traditional medicine. The complete plastome of the plant is known. It includes 130 genes (for a total of 154,961 bp) with 80 unique protein-coding genes. Phylogenetically, F. virosa is close to another medicinal plant, namely, Phyllanthus emblica Linn. (amla) [9,10]. F. virosa is a dioecious (male and female flowers on separate plants), multi-stemmed, fast-growing, bushy shrub or small tree with small, rounded leaves and tiny, greenish flowers, along with greenish-white fruit (Figure 1). The plant is commonly found in the drier parts of the African continent, the Arabian Peninsula, the Indian subcontinent, China (Guangxi, Guangdong, Guizhou, Hubei, Hunan provinces), southeast Asia and northern Australia. It is listed as an invasive species in Florida, USA. F. virosa usually grows well in a wide variety of habitats, including forest edges, bushland, grassland, woodland and thickets. In drier areas, it occurs mainly along water courses and in swampy habitats, sometimes on termite mounds and rocky slopes; it is also common in disturbed localities and fallow land from sea level up to 2300 m altitude. F. virosa usually grows up to 4 m tall, with many erect or arching branches. It is generally harvested from the wild, but it is also cultivated for its various useful medicinal values and as an ornamental hedge [11].
The ripened fruit of F. virosa are edible small white berries that are about 5 mm in diameter. At maturity, they are claimed to be juicy and sweet, though with a slightly bitter flavor. They are eaten by people, animals and birds when ripe. They are consumed fresh or boiled to make sauces or cooked together with the powder of cultivated crops to make porridge by the Kara and Kwego people in Ethiopia [18]. The pulp of the fruit is used to cure itching by rubbing on the skin [63]. The plant is recognized for its important nutritional properties with high palatability, and therefore, it is largely used as cattle fodder by breeders in Benin [64]. It has also been used as an antivenin medication for the treatment of snakebites, notably in Uganda [21,63,65].
A large diversity of bioactive natural products was identified from F. virosa and different types of plant extracts have revealed marked pharmacological activities. The present analysis offers an overview of the properties of F. virosa and its phytoconstituents. The traditional uses of the plant were analyzed based on an extensive review of the published information on Flueggea virosa and its different synonyms, notably Securinega virosa, which is also a commonly used name (Table 3). The objective was to highlight the benefits of the plant and to promote its use and new research activities. A comprehensive literature search was performed up to November 2023 using multiple databases and keywords. This review offers an exhaustive analysis of the scientific literature on F. virosa, including publications, reports and patents (mainly in the English language). This topic has not been addressed before and the phytochemical richness of the plant, notably the presence of unusual oligomeric alkaloids, piqued my interest.
2. Pharmacological Effects of F. virosa Extracts
Extracts prepared from the aerial parts of the plant or the roots have been largely used in traditional medicine to treat human diseases. The main properties of these extracts are discussed in turn (Figure 2).
2.1. Antiparasitic Activities
The plant is used against malaria in Kenyan folk medicine [66] and in Ghana [20] and other African countries to treat parasitic diseases. A methanol extract made from fresh leaves of F. virosa (collected in Comoros) was shown to inhibit the growth of the malaria parasite Plasmodium falciparum in vitro (IC50 = 2.28 and 3.64 µg/mL, with strains D6 and W2, respectively), with no significant effects on control non-infected Vero E6 cells (CC50 = 683 µg/mL). In vivo, the same extract was found to reduce the growth of Plasmodium berghei in infected mice by more than 70% when administered at 100 mg/kg/day. The effect was modest but noticeable with a mean survival time of 11 days versus 8 days for the vehicle-treated control group. No apparent sign of toxicity was observed [24,25,26]. Moderate antimalaria activity was confirmed using another chloroquine-resistant strain of P. falciparum, namely, strain K1 (IC50 = 7.6 µg/mL) [13]. A marked antiplasmodial activity was obtained with a methanol/water leaf extract and with a root decoction (IC50 = 3 µg/mL) [67]. But the best evidence of activity comes from a study that employed an ethyl acetate fraction of F. virosa leaves, which revealed dose-dependent activity in P. berghei-infected mice, with a nearly complete (86%) suppression of the animal parasitemia and a mean survival time of 17.2 days versus 10.8 days for the control animals [68]. The antimalaria activity was attributed to the presence of the polyphenol bergenin (see below). The plant is not a universal treatment because a similar methanolic extract was found to be inactive against the chloroquine-sensitive strain of P. falciparum PoW (IC50 > 50 µg/mL) [69]. Nevertheless, F. virosa is recognized among major African medicinal plants to treat malaria [70].
The alcoholic plant extract may be used also to combat other parasites. A leaf extract of F. virosa (collected in Mizoram, India) was found to be active against the intestinal cestode parasite Raillietina echinobothrida, which infects domestic fowl. The extract significantly reduced the alkaline phosphatase activity and induced protein catabolism in treated parasites [71]. The anthelmintic activity has been associated with the induction of significant destruction of the parasite tegument with intense vacuolization and swellings of the basal lamina leading to deformities in the cell organelles [72]. Antitrypanosomal activity was also reported with a petroleum ether extract of F. virosa, which was found to reduce the growth of Trypanosoma brucei rhodesiense (IC50 = 0.5 µg/mL), whereas the maximum tolerated concentration was about 20-fold higher (MTC = 9 µg/mL) [73]. Here, again, the antitrypanosomal activity of the extracts can be attributed to the presence of bergenin, which was shown to inhibit the growth of the bloodstream form of Trypanosoma brucei (IC50 = 1.0 µM) [74]. This natural product is a model compound for designing semi-synthetic products that are active against T. brucei [75]. The marked activity observed with the plant extract explains why traditional healers in Uganda use a powder from pounded roots of F. virosa (one spoonful orally) to treat sleeping sickness [73].
The root extract of F. virosa may be used to combat bilharziasis (schistosomiasis). A traditional recipe used in Somalia recommended the use of fresh roots, crushed and boiled with water to prepare a drinkable solution active against the parasite [42]. But no information about the efficacy of the preparation is available. Similarly, the use of a root decoction to treat urinary schistosomiasis is cited in a review on plants used to treat schistosomiasis in Niono District in Mali, but without further details [31]. Experimental data to support the antischistosomal activity are needed because an ancient study only revealed a minor effect of a leaf extract of F. virosa on cercariae and miracidia of Schistosoma mansoni [76].
2.2. Antimicrobial Effects
The traditional use of F. virosa for the treatment of dermal infections and wounds in Ghana has motivated studies aimed at characterizing the potential activity of various polar/apolar extracts of the plant against several organisms. Among the different medicinal plants tested, F. virosa was the most active, notably the chloroform extract derived for the root bark, which was found to be active against 13 microorganisms tested, but with a variable degree of activity. The CHCl3 extract was most potent against the pathogenic Gram-negative species Salmonella abony (MIC = 15.6 µg/mL) and mildly active against Staphylococcus aureus (MIC = 125 µg/mL). Interestingly, the extract was found to potentiate the activity of the antibiotic norfloxacin against a norfloxacin-resistant strain of S. aureus possessing the efflux pump NorA. Antimicrobial activities against other pathogens, such as Bacillus subtilis, Micrococcus flavus and Pseudomonas aeruginosa were also observed (17, 19 and 12 mm of growth inhibition when using an agar well diffusion technique) [77]. A recent study underlined the activity of F. virosa leaf extracts against methicillin-resistant Staphylococcus aureus (MRSA) [78], in agreement with other antimicrobial studies [79,80].
2.3. Antiepilepsy and Antipsychotic Activities
The testing of different Malian plants used traditionally to treat epilepsy and convulsions has revealed a marked activity with a leaf extract of F. virosa (with a plant collected in Bougouni District, Southern Mali). The extract inhibited the spontaneous discharge (SED) in a mouse cortical wedge preparation (IC50 = 0.2 mg/mL) and was efficient in the [3H]-flumazenil-binding assay (IC50 = 0.45 mg/mL), suggesting the presence of GABAergic compounds. The extract showed no effect during anticonvulsive testing in a model of pentylenetetrazol-induced seizure in mice [81]. A similar methanolic extract, which was also prepared from leaves of F. virosa, was shown to decrease aporphine-induced stereotypic climbing behavior, reduce swim-induced grooming in mice and increase the mean duration of ketamine-induced sleep in a murine model at a dose of 50 mg/kg [82,83]. The extract facilitated sleep induction, as was also shown in another study with a model of diazepam-induced sleep. In this case, an extract at 100 mg/kg was shown to prolong the duration of diazepam-induced sleep without affecting the exploratory behavior or the motor coordination of mice [84]. These observations using experimental models are consistent with the traditional use of plant extract as a sedative in children and for mental illnesses.
Sedative effects were also observed when using root bark extracts of F. virosa, notably with a butanol extract that contained tannins, saponins, alkaloids, flavonoids and cardiac glycosides, as with the methanol extract. A butanol fraction administered to mice (75 mg/kg) was found to reduce the mean onset of sleep and to double the mean duration of sleep. The sedative effects were clearly established [38,85]. A bark extract also displayed an antipsychotic activity, reducing swim-induced grooming activity in mice and the mean climbing score [86]. This latter study was performed using a residual aqueous fraction of methanol root bark extract but it gave results comparable with those obtained with the methanolic leaf extract, suggesting the implication of the same bioactive principles [83]. One of the active principles was clearly identified: the polyphenolic substance bergenin, which is present in the plant roots and aerial parts and was shown to possess a sleep-inducing property. This compound may, at least in part, be responsible for the sedative potential of the plant extracts [87]. Bergenin is found in many plants and is known to display antioxidant and antianxiety activities [88,89].
2.4. Antidiabetic Effects
One of the traditional usages of F. virosa is the treatment of diabetes-related impotence [54]. The chronic hyperglycemia that characterizes type 2 diabetes can lead to alteration of the vascular endothelium and associated tissue damage, notably a marked erectile dysfunction, which is considered the most important sexual dysfunction in men with diabetes mellitus [90]. Traditional healers in the Tanga region of Tanzania (northeastern part) use aqueous extracts of F. virosa to treat impotence and as an aphrodisiac [91]. Experimental data show that the oral administration of an aqueous extract prepared from dried roots of F. virosa reduces the blood glucose level in rabbit, but only during hyperglycemia. The extract has no effect once blood glucose has reached fasting levels. A dose-dependent (0.1–1.0 g/kg body weight) reduction in the area under the oral glucose tolerance curve was observed and no major toxicity was noted [47]. Antidiabetic effects have also been reported with a leaf extract of F. virosa administered intraperitoneally to diabetic rats. In this case, the extract reduced blood glucose levels after 4–24 h of administration, with an efficacy relatively close to that of the reference product insulin (at 24 h, the glucose levels were 328.2, 165.4 and 137.0 mg/dL in the groups treated with saline buffer (control), 100 mg/kg F. virosa leaf extract and 6 i.u./kg insulin, respectively) [92]. Thus, the extract clearly presented an insulin-like effect. Recently, a similar observation was made with a hydro-ethanolic plant extract (200 mg/kg) administered to streptozotocin-induced diabetic rats. The plant treatment reduced hyperglycemia and the progression of diabetic nephropathy [93]. F. virosa is regularly cited as a plant used traditionally in Africa to treat diabetes mellitus [91,94,95], but the natural products at the origin of the antidiabetic effects have not been identified at all. This aspect of the plant warrants further investigation. Among the antidiabetic compounds, it is worth mentioning the trimeric alkaloid fluevirosine A (see below). Its use in the preparation of blood-sugar-reducing medicine was patented in China [96].
2.5. Antidiabetic Effects
In relation to diabetes, we can also refer to the use of F. virosa extracts for the treatment of diabetes-associated pain. In Mali, decoctions made from the root, root bark or leaves are taken orally to relieve painful conditions, including stomach ache, menstrual pain and pain due to diabetes [32]. A methanol root bark extract of F. virosa was shown to display analgesic effects in animal models. The extract inhibited acetic acid-induced abdominal constrictions and attenuated formalin-induced neurogenic pain. The extract efficiently reduced abdominal writhing in mice in a dose-dependent manner. Remarkably, the inhibition of abdominal constriction with the plant extract at 25 mg/kg was greater than that of the standard non-steroidal analgesic piroxicam. Moreover, in a model of carrageenan-induced paw edema in rats, the methanol root bark extract was shown to efficiently reduce the edema diameter, with an efficacy close to that of the reference product ketoprofen [97]. In these in vivo tests, the most active doses were 25–100 mg/kg, which was largely inferior to the intraperitoneal median lethal dose (LD50 = 1.26 g/kg), suggesting that the extract is relatively safe at the analgesic doses [98]. Analgesic effects were also observed with an aqueous root extract (100–400 mg/kg) in a model of thermally induced pain in rats [99]. The aqueous root extract was apparently less efficient than the methanol root extract. Experiments were also performed with aqueous extracts prepared from the leaves and stems of F. virosa using a model of acetic acid-induced pain in mice. Interestingly, the stem extract displayed a marked analgesic effect (65% pain inhibition) coupled with a significant anti-inflammatory activity (59% inhibition of carrageenan-induced inflammation in mice) [100]. Collectively, these different studies support the traditional use of F. virosa to manage pain associated with different diseases and conditions, such as diabetes, but also benign prostatic hyperplasia (BPH).
2.6. Anticancer Effects
Decoctions from the roots of F. virosa are traditionally used to treat cancer patients. Notably, the Embu and Mbeere peoples in Kenya use root decoctions to treat prostate and breast cancers and kidney problems [27,101,102]. An antiproliferative action was also observed with a methanol extract and the human cancer cell lines RD (rhabdomysarcoma) and Hep-2C (laryngeal carcinoma) (IC50 = 11.3 and 7.2 μg/mL, respectively) [103]. The plant contains numerous alkaloids that can inhibit the proliferation of cancer cells. These different products are discussed below, notably a series of antiproliferative indolizidine alkaloids. There are also anticancer studies with the plant extracts, notably a study revealing the capacity of a methanolic leaf extract to block the proliferation of MCF7 breast cancer cells and NCI-H460 lung cancer cells (GI50 = 42.2 and 78.3 µg/mL, respectively). The root bark extract was a little less active than the leaf extract, and the chloroform fraction was more active than the aqueous fraction (essentially inactive) [104]. Interesting data have also been reported for a root bark extract, which was found to inhibit the proliferation of U-1242 glioblastoma multiforme tumor cells. Here, again, the chloroform fraction was the most active, and the crude methanol root bark extract was shown to modulate the epidermal growth factor receptor (EGFR) pathway [105].
2.7. Antioxidant Effects
Unsurprisingly, several studies referred to the antioxidant activity of F. virosa extracts, which is a property commonly observed with plant extracts containing flavonoids and phenolic compounds [106]. The best activity was obtained with ethanolic leaf extracts, whereas hexane extracts were much less active [107]. The leaf methanol extract presented higher antioxidant effects than the stem bark methanol extract [108,109]. The leaf extract was found to be as efficient as the reference product ascorbic acid (IC50 = 25 μg/mL in a DPPH assay) [88]. An extract made from the aerial parts of F. virosa revealed an antioxidant action that was a little inferior to that of ascorbic acid (IC50 = 0.01 versus 0.008 mg/mL, respectively), coupled with antifungal and antiproliferation effects [103]. Diverse antioxidant products were isolated from the leaves, such as bergenin, but also kaempferol 3-O-(4-galloyl)-β-D-glucopyranoside, 11-O-caffeoylbergenin and glucogallin acting as hydroxyl radical scavengers [77,110]. Root extracts also displayed a marked antioxidant profile [79]. The main component bergenin contributes significantly to the antioxidant effects. This phenolic compound alleviates hydrogen peroxide-induced oxidative stress in cells [111,112]. The antioxidant properties of F. virosa have been amply documented. Among 47 plant species (27 families) used in traditional medicine in Burkina Faso, a total extract of F. virosa revealed the most important antioxidant capacity [113].
2.8. Other Activities
Occasionally, other properties were observed when using extracts of F. virosa, such as antiarrhythmic effects attributed to bergenin [114]. The capacity of bergenin to protect from myocardial ischemia–reperfusion injury is well documented [115]. Recently, an ethanolic extract made from the aerial parts of F. virosa has demonstrated antisickle cell activity via a capacity to normalize the shape of the circulating abnormal erythrocytes, which is the so-called antifalcemic or antisickling activity [116]. The same type of activity was demonstrated previously with an aqueous methanolic leave extract, which inhibited sodium metabisulphite-induced sickling of hemoglobin sickle-shaped (Hbss) red blood cells in a concentration-dependent manner [117]. The antisickling is attributed to the presence of phenolic compounds capable of modulating the inflammatory response and reducing the vasocclusive crisis, but also to amino acids and flavonoids targeting Hbss polymerization and reducing endothelial dysfunctions [116].
There are not many studies that investigated the activity of F. virosa extracts against pathogenic fungi. Minor activities against Candida albicans [66] and the dermatophyte Trichophyton interdigitale (MIC = 125 µg/mL) [77] were noted. Similarly, modest activity was observed with a root bark extract tested against Candida albicans, C. glabrata and C. tropicalis [118]. The plant extracts are not very active against fungi and different alkaloids from the plant tested, as antifungal agents revealed little or no activity. For example, the alkaloid virosecurinine was found to be inactive against various fungi, such as Aspergillus niger, Penicillium viridicatum and Fusarium monoliforme [119].
Other pharmacological activities have occasionally been reported, such as an antidiarrheal activity with F. virosa methanolic extract [120,121] or the treatment of skin rashes and HIV infection [122]. The plant contains monomeric and dimeric alkaloids acting as inhibitors of HIV replication, such as virosinine A and flueggenine D (see below) [6,123,124]. A leaf powder of F. virosa can also be used as an insecticidal agent, for example, to control the development of the house fly Musca domestica and the transmission of diseases to humans [125].
3. Phytochemical Analyses of F. virosa Extracts
Over the past 30 years, the chemical constituents of the plant have been investigated using different geographic sources and different parts of the plant. Three main product categories can be defined: (i) polyphenols, which are typified by the lead product bergenin and flavonoids, (ii) multiple terpenoids and (iii) alkaloids, in addition to a few other products (saponins, polyketides, steroids, cyano glucosides, etc.). More than 80 natural products were identified belonging to the three main categories, which are discussed in turn below.
3.1. Polyphenols and Flavonoids
The major active component of F. virosa is certainly the polyphenolic isocoumarin bergenin, which is a C-glycoside of 4-O-methyl gallic acid and can be mainly found in the aerial parts of the plant, but also the roots, notably the root bark [87] (Figure 3a). Bergenin was discovered more than 140 years ago and is distributed in more than 11 plant species. This common compound displays a wide range of pharmacological properties (antiparasitic; antimicrobial; antinociceptive; antiarthritic; antidiabetic; antiarrhythmic; anticancer; and cardio-, hepato- and neuroprotective activities) [126,127]. It is likely at the origin of many of the effects observed with F. virosa extracts, notably the antioxidant, antianxiety, antiarrhythmic [114] and trypanocidal activities [74].
Its mechanism of action is multifactorial. A recent molecular modeling study postulated a direct interaction of bergenin with the ligand-binding domain of the estrogen receptor 1 (ESR-1) [128]. The hydroxyl groups on the C ring of the drug engage in contacts with the residues His-524 and Thr-347 of ESR-1 (Figure 3b). But other protein targets are implicated in the mechanism of action. Previous studies also suggested the binding of bergenin to the proteins NLRP3 (NOD-like receptor family-pyrin domain containing 3) inflammasome [129], antiapoptotic protein B-Cell Lymphoma 2 (Bcl-2) [130], enzyme galectin-3 [131] and several other proteins [132,133,134,135].
Derivatives were also found, such as norbergenin, 11-O-acetyl bergenin and 11-O-caffeoylbergenin isolated from the leaves of F. virosa, together with flavonoid glycosides (e.g., kaempferol 3-O-(4-galloyl)-β-d-glucopyranoside) and phenolic derivatives [110,136,137]. Among these products, the demethoxylated form of bergenin, which is known as norbergenin (Figure 3a), is a potent inhibitor of oxidative stress, acting as an efficient scavenger for oxygen-based radicals [138] and a potent anti-inflammatory agent [139].
Analytical methods were implemented to quantify the contents of products in the plant [140]. It was determined that the aerial parts of F. virosa contain 15.2% (w/w) of bergenin [141,142]. A methanolic extraction process can be applied to extract the product from the leaves [114]. This abundant polyphenolic compound is used as a template for the design of novel semi-synthetic antiparasitic drugs [75]. The naturally occurring derivative 11-O-caffeoylbergenin (Figure 3a) displays marked antioxidant and antiarthritic activities [143,144]. Bergenin itself is an anticancer agent that acts as a suppressor of glycolysis and, therefore, is useful to overcome chemo- and radio-resistance [145,146]. Its mechanism of action notably implicates the targeting of the antiapoptotic protein Mcl-1 (Myeloid leukemia 1), which is frequently overexpressed in human malignancies [147]. Bergenin presents a limited bioavailability, but there are options to overcome this limitation, notably through the design of semisynthetic derivatives and the use of micro-/nano-delivery systems for a controlled release of the product [133,148,149]. The abundance of bergenin in F. virosa is a major element of the bioactivity of the plant. Nevertheless, the pharmacological use of this compound (or the plant) must be limited because, at high doses, the compound can impair the physiology of different organs (liver, kidneys, intestine) and some of its metabolites are suspected to be carcinogenic [150].
Other polyphenolic compounds can be found in F. virosa, notably the common dietary flavonol rutin [151]; the ellagitannins corilagin, acalyphidin M and geraniin; the gallic acid derivative glucogallin [109]; and (2S,3R)-4E-dehydrochebulic acid trimethyl ester [152]. These natural products, which are often present in natural medicines, are potent antioxidant compounds [153]. Corilagin is well recognized for its anticancer effects [154,155] and its anti-inflammatory effects [153].
3.2. Terpenoids
A recent study characterized a series of polyoxygenated terpenoids designated podovirosanes A–F from the roots of F. virosa (Figure 4). They are tricyclic molecules with a mild anti-inflammatory activity, at least for some of them, but their bioactivity remains little known at present [156]. They bear a structural analogy with flueggenoids A–E isolated from the twigs and leaves of F. virosa (Figure 4). In this series, only flueggenoid C revealed a modest inhibitory activity against hepatitis C virus (HCV) infection (IC50 = 23.3 µM) and no cytotoxicity against human hepatocellular carcinoma Huh7.5 cells (IC50 = 92.8 µM). The other compounds were inactive. Several 13-methyl-ent-podocarpanes were co-isolated during the process to obtain the flueggenoids, but these compounds also revealed a very modest anti-HCV activity [137]. Another series of terpenoids was isolated from the roots of F. virosa and evaluated for their potential anti-HCV activity. The series of 18 compounds included many ent-podocarpanes, which were more or less rearranged. Two compounds, including compounds 4 and 8 (Figure 4), exhibited a modest inhibitory activity against HCV infection (IC50 = 14.4 and 17.8 µM, respectively) [157]. The tricyclic podocarpatriene 8 warrants further investigations as an antinociceptive agent because it bears a structural analogy with the podocarpatrienone that is active in models of hyperalgesia [158]. Another compound, namely, 9(10− > 20)-abeo-ent-podocarpane, was found in a twigs and leaves extract and characterized as an inhibitor of HCV infection in Huh7.5.1 cell cultures (EC50 =27.4 µM). Better activity was obtained with the derivative 4 (Figure 4), with an EC50 of 7.7 µM and a satisfactory selectivity index (SI = 9.2) [152]. Podocarpane-type diterpenoids are not extremely frequent in nature [159] and their bioactivities are generally little known.
In the same group of podocarpane derivatives, flueggrenes A and B (Figure 4) are two trinorditerpenes that were also isolated from the roots of F. virosa. They display a weak anti-HCV infection activity (IC50 = 13.0 and 26.3 µM, respectively) and they are both able to inhibit elastase release by human neutrophils in response to cytochalasin B (IC50 = 4.3 and 9.6 µM, respectively) [160]. In the same vein, the authors also described a series of dinorditerpenes found in F. virosa roots, including a few potent compounds. An interesting molecule is the podocarpatriene 11 (7α,20-epoxy-3α-hydroxy-12-methoxy-13-methyl-ent-podocarp-8,11,13-triene) with an intramolecular ether linkage. This compound was found to reduce HCV infection (IC50 = 6.6 µM) while being non-cytotoxic to Huh7.5 cancer cells (IC50 > 400 µM) [161]. Its high therapeutic index (TI = 56) encouraged further studies, as well as the total synthesis of the compound renamed (+)-elevenol (Figure 4) and that of a close analog named (+)-przewalskin [162]. There is now a convenient synthetic procedure to access these antiviral compounds. In fact, podovirosane B is a 6-hydroxy derivative of (+)-elevenol.
Diverse triterpenoids were isolated from the leaves and twigs of F. virosa and evaluated as anticancer agents. Friedelin, heptanolide, epifriedelanol, stigmasterol and betulinic acid were identified and the later pentacyclic triterpene was found to be the most active against cancer cell proliferation in vitro [163]. Nothing is surprising here, as betulinic acid is well known to display marked anticancer activities [164,165]. Other common triterpenoids were identified from extracts of the stem bark and leaves of the plant, such as the saponins oleanic acid (C15) and ursolic acid (C16), both of which are very abundant in the stem bark (88–94 mg/g), but also asiatic acid, maslinic acid, corosolic acid and other minor terpenoids (1–7 mg/g) [109,166]. Ursolic acid is a potent anti-inflammatory and antioxidant compound that is of interest in combating multiple diseases, notably urogenital cancer [167,168].
3.3. Alkaloids
Flueggea (or Securinega) alkaloids have been studied for more than half a century. It started in the mid-1950s with the isolation of securinine from Securinega suffruticosa (Pall.) Rehd. in Russia [169], followed by the discovery of virosecurinine from F. virosa in 1963 [170] (Figure 5). Over the past sixty years, many alkaloids have been identified from all parts of plants collected in different territories. The plant generates a multitude of alkaloids, including oligomers biosynthesized from (-)-norsecurinine as a monomeric precursor. Dimers, trimers, tetramers and pentamers were identified, such as flueggenines C and D (dimers), fluevirosine D (trimer), fluevirosinine A (tetramer) and fluevirosinines G–J (pentamers), all from F. virosa. Many alkaloid-related studies were performed with the species S. suffruticosa, with a wide range of structures [171,172,173]. Here, we focus on selected alkaloids from F. virosa only.
Securinine can be considered the leading alkaloid from F. virosa; it is also largely present in other Flueggea species [174]. Among the diverse pharmacological effects of the compound, the anticancer properties deserve special interest owing to the potency of the compound and its mechanism of action that implicates several signaling pathways, notably the AKT/mTOR/S6K pathway, JAK/STAT pathway and MAPK (MEK/ERK) pathway, leading to mitochondria-mediated apoptosis within cancer cells [175,176]. Different molecular targets were postulated, notably a direct interaction between securinine and tubulin (Kd = 9.7 μM) [177]. Securinine has been largely used as a prototypic scaffold for the design of analogs developed as antitumor agents and/or neuroprotective compounds [178,179,180].
The structural diversity within the securinine-type family of alkaloids is large, with about 80 natural products identified from Flueggea species [172,179]. A specific search for alkaloids identified from F. virosa led to the inventory of 64 compounds (Table 4), which were mostly discovered from the aerial parts and occasionally from the roots or the fruit of the plant. There are also a few distinct alkaloids, such as the small molecule hordenine isolated from the roots and likely serving as a biosynthetic precursor for more complex structures [181]. But the vast majority of alkaloids in F. virosa belong to the securinine family. There are many monomeric alkaloids, such as bubbialine, virosine A and niruroidine, but also dimeric (flueggines A and B), trimeric (Flueggether D), tetrameric (fluevirosinine D) and even pentameric (fluevirosinines G–J) alkaloids (Figure 5). Oligomerization of plant alkaloids is not extremely frequent, but it is a feature of the Flueggea species, notably F. virosa [182,183]. A biosynthetic pathway was proposed, starting from (-)-norsecurinine (monomer) and leading to flueggenines C and D (dimers), and then fluevirosine D (trimer) and fluevirosinine A (tetramer) [184] (Figure 5). One must admit that the many product names are a little confusing. It is not easy to navigate the diverse range of alkaloid names and their properties.
Apart from the largely studied securinine, the pharmacological effects of all other alkaloids in the series were superficially investigated. Usually, one or two isolated tests were performed, and no comparison can be made between the series. Nevertheless, one molecule emerged from our analysis, namely, the dimeric indolizidine compound flueggine B, which revealed potent antiproliferative activities against two breast cancer cell lines (MCF-7 and MDA-MB-231 cells with IC50 values of 135 and 147 nM, respectively), whereas the analog flueggine A was more than 400-fold less active (IC50 values of 60 and 68 µM, respectively). Flueggine B is considerably more potent than other compounds that were also tested against the same cell lines, such as dimeric indole fluevirines E and F, both of which were almost inactive (41% growth inhibition with fluevirine F at 40 µM) [186] (Figure 6). Another indolizidine alkaloid dimer, namely, flueggedine (Figure 5), showed no activity against the two identical cell lines MCF-7 and MDA-MB-231 [183]. Other related products were isolated from the fruit of the plant, notably the dimers norsecurinamine A and B (Figure 7), but their pharmacological properties are not known at present [202]. The indole alkaloids named flueindolines A–C were also obtained from the ripe fruit of F. virosa. Flueindolines A and B are tricyclic indoles, whereas flueindoline C is a spirooxindole derivative; they possibly derive from tryptophane [187] (Figure 7).
Flueggine B emerged as an atypical compound and perhaps the most interesting in the family (Figure 6). The situation is reminiscent of that previously observed with other dimeric indole alkaloids, such as vinblastine and vincristine derived from the Madagascar periwinkle (Catharanthus roseus, formerly Vinca rosea). In this case, the dimeric indole alkaloids were much more potent than their monomeric precursor molecules vindoline and catharanthine [207]. It would make sense to evaluate the interaction of flueggine B with tubulin and the effects on the microtubule dynamics. A preliminary docking analysis suggests that flueggine B can form very stable complexes with the α/β-tubulin dimer. The molecular modeling pointed to a binding cavity at the interface of the protein dimer that is suitable to accommodate the monomer securinine or the dimer flueggine B. The calculated empirical energies of interaction (ΔE) were −44.60 and −63.40 kcal/mol for securinine and flueggine B, respectively, and the free energies of hydration (ΔG) were −17.10 and −24.60 kcal/mol, respectively. The binding cavity (297.1 Å3) is large enough to accommodate the dimeric compound, and its interaction with tubulin is stabilized via the formation of two key hydrogen bonds (with residues Leu242 and Lys352), in addition to an array of 16 van der Waals contacts and alkyl/π-alkyl interactions. This is twice the number of H-bonds and contacts observed with securinine under identical conditions (Figure 8). The dimeric compound reveals a prominent capacity to interact with tubulin. We are currently investigating the binding process further. Other alkaloids can be found in F. virosa, such as the pyrrolidone donaxanine (Figure 6), and a few indole alkaloids identified from the fruit, such as N-methyltryptamine and strychnocarpine [187]. This latter product is a β-carbolinone alkaloid with a low affinity for the major brain receptors (serotonin, benzodiazepine, tryptamine, opiate and GABA receptors), whereas N-methyltryptamine is a mild psychoactive product (Figure 7) [208,209].
3.4. Other Compounds
A few other bioactive molecules were isolated from F. virosa, such as the cyano glycoside menisdaurin found in the aerial parts of the plant [139,140] and its analog amiroside [212] (Figure 6). Menisdaurin displays anti-inflammatory properties and possibly functions as an inhibitor of cyclooxygenase-2 [213,214]. Recently, two polyketides were identified from the roots of the plant, namely, (R)-8-methoxymellein and (3R)-5-hydroxy-8-O-methylmellein, but the authors suspect that they originated from root symbiotic fungi [156]. Other compounds could be cited, including a few steroids and fatty acids [215].
4. Discussion and Perspectives
The traditional use of the plant F. virosa is widespread, notably in Africa, where it is a common folk medicine used to treat many pathologies, such as parasitic infections, inflammatory diseases and diabetes. Decoctions from the plant leaves and twigs are largely used and the fruit are consumed locally. It is a prominent medicinal and nutritional plant. The phytochemical interest of F. virosa resides in its large number of bioactive molecules, with two abundant and major products: the polyphenol bergenin and the alkaloid securinine. This latter compound is the leader of the so-called Securinega alkaloid family, which includes more than 80 products, with about 65 identified in F. virosa. A major phytochemical peculiarity of F. virosa is the generation of alkaloid oligomers, with the production of large structures (up to tetra and pentamers) rarely seen in other plants. Alkaloid dimers are relatively frequent (notably among Vinca alkaloids, for example), but tetra and pentamers are much rarer. A few trimeric indole alkaloids have been reported [216,217,218], but higher-order structures are extremely rare. For example, a tetramer and hexamer of the alkaloid terguride were isolated, but the oligomerization is limited and strongly reduces the high affinity of terguride for 5-HT2A receptors [219,220]. In the case of F. virosa, the oligomerization of securinine leads to multiple series of compounds, such as flueggenines, fluevirosinines and flueggedines. The dimers are particularly interesting compounds endowed with anticancer activities. The pentamers are unique. The compounds fluevirosinines G–J represent C-60 macro-structures (fluevirosinine G = C60H65N5O10), which are rarely observed. F. virosa is a model plant to study alkaloid oligomerization, the corresponding biosynthetic pathways and the pharmacology of macro-alkaloids.
The strong capacity of flueggine B to interact with tubulin looks promising and should encourage the design and synthesis of novel synthetic dimeric derivatives of securinine. Dimeric securinine analogs endowed with neuritogenic and/or antileukemic activities have been reported [221,222]. One of the synthetic dimers, which was designated SN3-L6, was shown to function as an activator of translation and neuronal differentiation in neural progenitor cells [223]. The same compound also potently and selectively inhibited the proliferation of HL-60 leukemia cells, triggering their apoptosis, but with no effects on megakaryocytes and granulocytes. An investigation into the mechanism of action of this dimeric compound pointed out the mitotic kinases Aurora, with an inhibition of the expression phospho-Aurora kinase A and B expression in HL60 cells treated with this SN3-L6 [222]. This is interesting because Aurora kinases play a major role in mitosis and are implicated in tubulin complexes. This is coherent with a possible modulation of tubulin polymerization via tubulin binding. Both securinine and SN3-L6 are mitotic blockers. These experimental data, coupled with our preliminary docking information, encourage us to investigate the binding of all flueggines to tubulin. Flueggines A and B are present in F. virosa [188], but there are also 20 related products, including 9 flueggenines (A–I), 10 fluevirosinines (A–J) and flueggedine found in diverse Flueggea or Securinega species [210]. Their tubulin capacities will be compared using the same in silico approach as was used with flueggine B and other tubulin-binding natural products [43,211]. The identification of tubulin as a potential molecular target for securinine and flueggine B should help to design new molecules using guided structure-binding relationships. This is an area of major drug design interest.
The plant F. virosa warrants further investigation, in particular the fruit, which are abundant and edible. The white berries are often consumed by the local population but they are not largely exploited and apparently not commercialized in any form, despite their nutritional and medicinal value. In South Africa, the Mapulana people (an indigenous community of Ehlanzeni district in Mpumalanga province, South Africa) consume the fruit, locally called ditlhalabu or ditlhakawume [43]. The berries are consumed in other African countries (Ethiopia, Tanzania) and also in India, notably in the area of the Warud tahsil village, Amravati district in Maharashtra, for their sweet taste and nutritional value [22]. In Ghana, the pulp of the fruit is used for healing wounds and anti-itching, as well as an extract ointment prepared from the leaves of the plant [63,224]. With no doubt, this fruit deserves further studies, with a proper evaluation of its nutritional intake and safety profile. A few alkaloids were isolated from the fruit, but at present, there is no specific study of their pharmacological effects. The biochemical and nutritional composition of F. virosa fruit shall be investigated.
In conclusion, this study underlined the pharmacological benefits of the plant F. virosa, which is largely used in traditional medicine in a large part of the world. This Flueggea species is a phytochemical reservoir, with a large number of flavonoids and alkaloids isolated from different parts of the plant. The two emblematic compounds, namely, bergenin and securinine, along with many derivatives and analogs, are at the origin of the various pharmacological activities, notably anti-inflammatory, antioxidant, antiparasitic, antidiabetic, anticancer and analgesic effects. A remarkable feature of F. virosa is the presence of norsecurinine oligomers, such as the anticancer dimer flueggine B, which possibly acts as a tubulin binder. The nutritional and pharmacological properties of the plant, notably the white berries consumed locally, shall be investigated further. In today’s rapidly changing world, with a major need to provide healthy food to a growing human population, it is important to accentuate access to a potential new source of food. The fruit of F. virosa (and other Flueggea species) represents an option to consider but within a frame of ecological protection.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Data are contained within the article.
Acknowledgments
The OncoLille Institute has been supported by a grant from Contrat de Plan Etat-Region CPER Cancer 2015–2020. The author thanks Gérard Vergoten (University of Lille, France), who kindly performed the modeling of flueggine B bound to tubulin presented in Figure 8.
Conflicts of Interest
The author declares no conflicts of interest. Christian Bailly is the founder of OncoWitan. This company had no role in the design of the study; in the collection, analysis, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
References
- Barker, C.; van Welzen, P.C. Flueggea (Euphorbiaceae s. l. or Phyllanthaceae) in Malesia. System Bot. 2010, 35, 541–551. [Google Scholar]
- Wu, Z.L.; Huang, X.J.; Xu, M.T.; Ma, X.; Li, L.; Shi, L.; Wang, W.J.; Jiang, R.W.; Ye, W.C.; Wang, Y. Flueggeacosines A-C, Dimeric Securinine-Type Alkaloid Analogues with Neuronal Differentiation Activity from Flueggea suffruticosa. Org. Lett. 2018, 20, 7703–7707. [Google Scholar] [PubMed]
- Soysa, P.; De Silva, I.S.; Wijayabandara, J. Evaluation of antioxidant and antiproliferative activity of Flueggea leucopyrus Willd (katupila). BMC Complement. Altern. Med. 2014, 14, 274. [Google Scholar]
- Mendis, A.S.; Thabrew, I.; Samarakoon, S.R.; Tennekoon, K.H. Modulation of expression of heat shock proteins and apoptosis by Flueggea leucopyrus (Willd) decoction in three breast cancer phenotypes. BMC Complement. Altern. Med. 2015, 15, 404. [Google Scholar] [CrossRef] [PubMed]
- Li, B.; Gilbert, M.G.; Fischer, G.; Meyer, C.A. Flora of China; Science Press: Beijing, China; Missouri Botanical Garden Press: St. Louis, MO, USA, 2008; Volume 11, p. 178. [Google Scholar]
- Zhang, H.; Zhu, K.K.; Han, Y.S.; Luo, C.; Wainberg, M.A.; Yue, J.M. Flueggether A and Virosinine A, Anti-HIV Alkaloids from Flueggea virosa. Org. Lett. 2015, 17, 6274–6277. [Google Scholar] [CrossRef] [PubMed]
- Tabuti, J.R.S. Flueggea virosa (Roxb. ex Willd.) Voigt. In Plant Resources of Tropical Africa 11(1): Medicinal Plants 1; Schmelzer, G.H., Gurib-Fakim, A., Eds.; Prota Foundation: Wageningen, The Netherlands, 2008; pp. 305–308. Available online: http://database.prota.org/PROTAhtml/Flueggea%20virosa_En.html (accessed on 15 December 2023).
- d’Errico, F.; Backwell, L.; Villa, P.; Degano, I.; Lucejko, J.J.; Bamford, M.K.; Higham, T.F.; Colombini, M.P.; Beaumont, P.B. Early evidence of San material culture represented by organic artifacts from Border Cave, South Africa. Proc. Natl. Acad. Sci. USA 2012, 109, 13214–13219. [Google Scholar] [CrossRef]
- Wang, H.T.; Wang, H.X.; Zhu, Z.X.; Wang, H.F. Complete plastome sequence of Flueggea virosa (Roxburgh ex Willdenow) Voigt (Phyllanthaceae): A medicinal plant. Mitochondrial DNA B Resour. 2020, 5, 2650–2651. [Google Scholar]
- Saini, R.; Sharma, N.; Oladeji, O.S.; Sourirajan, A.; Dev, K.; Zengin, G.; El-Shazly, M.; Kumar, V. Traditional uses, bioactive composition, pharmacology, and toxicology of Phyllanthus emblica fruits: A comprehensive review. J. Ethnopharmacol. 2022, 282, 114570. [Google Scholar]
- Available online: https://tropical.theferns.info/viewtropical.php?id=Flueggea+virosa (accessed on 15 December 2023).
- Fah, L.; Klotoé, J.R.; Dougnon, V.; Koudokpon, H.; Fanou, V.B.A.; Dandjesso, C.; Loko, F. Étude ethnobotanique des plantes utilisées dans le traitement du diabète chez les femmes enceintes à Cotonou et Abomey-Calavi (Bénin). J. Animal Plant Sci. 2013, 18, 2647–2658. [Google Scholar]
- Sanon, S.; Gansane, A.; Ouattara, L.P.; Traore, A.; Ouedraogo, I.N.; Tiono, A.; Taramelli, D.; Basilico, N.; Sirima, S.B. In vitro antiplasmodial and cytotoxic properties of some medicinal plants from western Burkina Faso. Afr. J. Lab. Med. 2013, 2, 81. [Google Scholar]
- Ouôba, P.; Lykke, A.M.; Boussim, J.; Guinko, S. La flore médicinale de la Forêt Classée de Niangoloko (Burkina Faso). Etudes Flor. Vég. Burkina Faso 2006, 10, 5–16. [Google Scholar]
- Malzy, P. Quelques plantes du Nord Cameroun et leurs utilisations. J. D’agriculture Trop. Bot. Appliquée 1954, 1, 148–179. [Google Scholar]
- Boulesteix, M.; Guinko, S. Plantes Médicinales Utilisées par les Gbayas dans la Région de Bouar (Empire Centrafricain); Quatrième colloque du Conseil Africain de Malgache pour L’enseignement Supérieur (C.A.M.E.S.): Libreville, Gabon, 1979; pp. 23–52. [Google Scholar]
- Terashima, H.; Ichikawa, M. A comparative ethnobotany of the Mbuti and Efe hunter-gatherers in Itury forest, Democratic Republic of Congo. Afr. Study Monogr. 2003, 24, 1–168. [Google Scholar]
- Teklehaymanot, T.; Giday, M. Ethnobotanical study of wild edible plants of Kara and Kwego semi-pastoralist people in Lower Omo River Valley, Debub Omo Zone, SNNPR, Ethiopia. J. Ethnobiol. Ethnomed. 2010, 6, 23. [Google Scholar]
- Wondimu, T.; Asfaw, Z.; Kelbessa, E. Ethnobotanical study of medicinal plants around ‘Dheeraa’ town, Arsi Zone, Ethiopia. J. Ethnopharmacol. 2007, 112, 152–161. [Google Scholar] [PubMed]
- Asase, A.; Akwetey, G.A.; Achel, D.G. Ethnopharmacological use of herbal remedies for the treatment of malaria in the Dangme West District of Ghana. J Ethnopharmacol 2010, 129, 367–376. [Google Scholar]
- Renu, S.N.; Rawat, A.; Kaur, J.; Kumar, S.; Fatima, N. Taxonomy, phytochemistry, pharmacology and traditional uses of Flueggea virosa (Roxb. ex Willd.) Royle: A Review. Int. J. Life Sci. 2018, 6, 579–585. [Google Scholar]
- Satpute, S.V.; Sinkar, S.R.; Sarode, A.M. Wild edible fruit plants and their use by tribal people and local villagers: A survey-based study, Int. Res. J. Sci. Eng. 2021, A11, 256–262. [Google Scholar]
- Ambe, G.A.; Malaisse, F. Diversité des plantes médicinales et ethnotaxonomie en pays Malinké de Côte d’Ivoire. In Des Sources du Savoir aux Médicaments du Futur: Actes du 4e Congrès Européen D’ethnopharmacologie = From the Sources of Knowledge to the Medicines of the Future: Proceedings of the 4th European Congress on Ethnopharmocology; Fleurentin, J., Pelt, J.M., Mazars, G., Eds.; Lejosne J.C. (trad.), Cabalion Pierre (collab.); Paris (FRA); Metz: IRD; SFE; Congrès Européen d’Ethnopharmacologie, 4; FRA: Metz, France, 2000; pp. 331–338. ISBN 2-7099-1504-9. [Google Scholar]
- Muthaura, C.N.; Rukunga, G.M.; Chhabra, S.C.; Mungai, G.M.; Njagi, E.N. Traditional antimalarial phytotherapy remedies used by the Kwale community of the Kenyan Coast. J. Ethnopharmacol. 2007, 114, 377–386. [Google Scholar] [CrossRef] [PubMed]
- Muthaura, C.N.; Rukunga, G.M.; Chhabra, S.C.; Omar, S.A.; Guantai, A.N.; Gathirwa, J.W.; Tolo, F.M.; Mwitari, P.G.; Keter, L.K.; Kirira, P.G.; et al. Antimalarial activity of some plants traditionally used in treatment of malaria in Kwale district of Kenya. J. Ethnopharmacol. 2007, 112, 545–551. [Google Scholar] [CrossRef] [PubMed]
- Muthaura, C.N.; Keriko, J.M.; Mutai, C.; Yenesew, A.; Gathirwa, J.W.; Irungu, B.N.; Nyangacha, R.; Mungai, G.M.; Derese, S. Antiplasmodial potential of traditional antimalarial phytotherapy remedies used by the Kwale community of the Kenyan Coast. J. Ethnopharmacol. 2015, 170, 148–157. [Google Scholar] [CrossRef] [PubMed]
- Kareru, P.G.; Kenji, G.M.; Gachanja, A.N.; Keriko, J.M.; Mungai, G. Traditional medicines among the Embu and Mbeere peoples of Kenya. Afr. J. Tradit. Complement. Altern. Med. 2007, 4, 75–86. [Google Scholar]
- Wanzala, W.; Takken, W.; Mukabana, W.R.; Pala, A.O.; Hassanali, A. Ethnoknowledge of Bukusu community on livestock tick prevention and control in Bungoma district, western Kenya. J. Ethnopharmacol. 2012, 140, 298–324. [Google Scholar] [PubMed]
- Gathirwa, J.W.; Rukunga, G.M.; Mwitari, P.G.; Mwikwabe, N.M.; Kimani, C.W.; Muthaura, C.N.; Kiboi, D.M.; Nyangacha, R.M.; Omar, S.A. Traditional herbal antimalarial therapy in Kilifi district, Kenya. J. Ethnopharmacol. 2011, 134, 434–442. [Google Scholar] [CrossRef] [PubMed]
- Nicolas, J.P. Plantes médicinales du Nord de Madagascar. In Ethnobotanique Antakarana et Informations Scientifiques; du Monde, J., Ed.; Jardins du Monde: Brasparts, France, 2012; 296p. [Google Scholar]
- Bah, S.; Diallo, D.; Dembélé, S.; Paulsen, B.S. Ethnopharmacological survey of plants used for the treatment of schistosomiasis in Niono District, Mali. J. Ethnopharmacol. 2006, 105, 387–399. [Google Scholar] [PubMed]
- Danton, O.; Somboro, A.; Fofana, B.; Diallo, D.; Sidibés, L.; Rubat-Coudert, C.; Marchand, F.; Eschalierd, A.; Ducki, S.; Chalard, P. Ethnopharmacological survey of plants used in the traditional treatment of pain conditions in Mali. J. Herbal. Med. 2019, 17–18, 100271. [Google Scholar]
- Van der Steur, L. Plantes médicinales utilisées par les Peul du Sénégal Oriental. Rev. Med. Pharm. Afr. 1994, 8, 189–200. [Google Scholar]
- Inngjerdingen, K.; Nergård, C.S.; Diallo, D.; Mounkoro, P.P.; Paulsen, B.S. An ethnopharmacological survey of plants used for wound healing in Dogonland, Mali, West Africa. J. Ethnopharmacol. 2004, 92, 233–244. [Google Scholar] [PubMed]
- Sanogo, R. Medicinal plants traditionally used in Mali for dysmenorrhea. Afr. J. Tradit. Complement. Altern. Med. 2011, 8, 90–96. [Google Scholar] [CrossRef]
- Ribeiro, A.; Romeiras, M.M.; Tavares, J.; Faria, M.T. Ethnobotanical survey in Canhane village, district of Massingir, Mozambique: Medicinal plants and traditional knowledge. J. Ethnobiol. Ethnomed. 2010, 6, 33. [Google Scholar] [CrossRef] [PubMed]
- Adam, J.G.; Echard, N.; Lescot, M. Plantes médicinales Hausa de l’Ader (République du Niger). J. Agric. Trop. Bot. Appl. (JATBA) 1972, 19, 259–399. [Google Scholar] [CrossRef]
- Magaji, M.G.; Yaro, A.H.; Musa, A.M.; Anuka, J.A.; Abdu-Aguye, I.; Hussaini, I.M. Central depressant activity of butanol fraction of Securinega virosa root bark in mice. J. Ethnopharmacol. 2012, 141, 128–133. [Google Scholar] [PubMed]
- Soladoye, M.O.; Ikotun, T.; Chukwuma, E.C.; Ariwaodo, J.O.; Ibhanesebor, G.A.; Agbo-Adediran, O.A.; Owolabi, S.M. Our plants, our heritage: Preliminary survey of some medicinal plant species of Southwestern University Nigeria Campus, Ogun State, Nigeria. Ann. Biol. Res. 2013, 4, 27–34. [Google Scholar]
- Kerharo, J. Senegalese pharmacopoeia: Catalog of medicinal and toxic plants of Wolof and Serer, augmented by the mention of common and vernacular names, of properties and uses, generally recognized in traditional medicine. Ann. Pharm. Fr. 1967, 25, 385–438. [Google Scholar]
- Kerharo, J.; Adam, J.G. Plantes médicinales et toxiques des Peuls et des Toucouleurs du Sénégal. J. Agric. Trop. Bot. Appl. (JATBA) 1964, 11, 384–444+543–599. [Google Scholar]
- Samuelsson, G.; Farah, M.H.; Claeson, P.; Hagos, M.; Thulin, M.; Hedberg, O.; Warfa, A.M.; Hassan, A.O.; Elmi, A.H.; Abdurahman, A.D.; et al. Inventory of plants used in traditional medicine in Somalia. II. Plants of the families Combretaceae to Labiatae. J. Ethnopharmacol. 1992, 37, 47–70. [Google Scholar] [CrossRef] [PubMed]
- Mashile, S.P.; Tshisikhawe, M.P.; Masevhe, N.A. Indigenous fruit plants species of the Mapulana of Ehlanzeni district in Mpumalanga province, South Africa. S. Afr. J. Bot. 2019, 122, 180–183. [Google Scholar] [CrossRef]
- deWet, H.; Ngubane, S.C. Traditional herbal remedies used by women in a rural community in northern Maputaland (South Africa) for the treatment of gynaecology and obstetric complaints. S. Afr. J. Bot. 2014, 94, 129–139. [Google Scholar] [CrossRef]
- Mongalo, N.I.; Makhafola, T.J. Ethnobotanical knowledge of the lay people of Blouberg area (Pedi tribe), Limpopo Province, South Africa. J. Ethnobiol. Ethnomed. 2018, 14, 46. [Google Scholar] [CrossRef]
- Rasethe, M.T.; Semenya, S.S.; Potgieter, M.J.; Maroyi, A. The utilization and management of plant resources in rural areas of the Limpopo Province, South Africa. J. Ethnobiol. Ethnomed. 2013, 9, 27. [Google Scholar] [CrossRef]
- Moshi, M.J.; Uiso, F.C.; Mahunnah, R.L.A.; Mbwanbo, Z.H.; Kapingu, M.C. A survey of plants used by traditional healers in the management of non-insulin dependent diabetes mellitus. East Cent. Afr. J. Pharm. Sci. 2000, 3, 30–39. [Google Scholar]
- Haerdi, F. Afrikanische Heilpflanzen. Die Eingeborenen-Heilpflanzen des Ulanga- Distriktes Tanganjikas (Ostafrika). Acta Trop. 1964, S8, 1–278. [Google Scholar]
- Moshi, M.J.; Otieno, D.F.; Weisheit, A. Ethnomedicine of the Kagera Region, north western Tanzania. Part 3: Plants used in traditional medicine in Kikuku village, Muleba District. J. Ethnobiol. Ethnomed. 2012, 8, 14. [Google Scholar]
- Hedberg, I.; Hedberg, O.; Madati, P.J.; Mshigeni, K.E.; Mshiu, E.N.; Samuelsson, G. Inventory of plants used in traditional medicine in Tanzania. I. Plants of the families Acanthaceae-Cucurbitaceae. J. Ethnopharmacol. 1982, 6, 29–60. [Google Scholar] [PubMed]
- Amri, E.; Kisangau, D.P. Ethnomedicinal study of plants used in villages around Kimboza forest reserve in Morogoro, Tanzania. J. Ethnobiol. Ethnomed. 2012, 8, 1. [Google Scholar] [CrossRef]
- Chhabra, S.C.; Mahunnah, R.L.; Mshiu, E.N. Plants used in traditional medicine in eastern Tanzania. III. Angiosperms (Euphorbiaceae to Menispermaceae). J. Ethnopharmacol. 1990, 28, 255–283. [Google Scholar] [CrossRef] [PubMed]
- Kpodar, M.S.; Karou, S.D.; Katawa, G.; Anani, K.; Gbekley, H.E.; Adjrah, Y.; Tchacondo, T.; Batawila, K.; Simpore, J. An ethnobotanical study of plants used to treat liver diseases in the Maritime region of Togo. J. Ethnopharmacol. 2016, 181, 263–273. [Google Scholar] [CrossRef]
- Kamatenesi-Mugisha, M.; Oryem-Origa, H. Traditional herbal remedies used in the management of sexual impotence and erectile dysfunction in western Uganda. Afr. Health Sci. 2005, 5, 40–49. [Google Scholar] [PubMed]
- Tabuti, J.R.; Lye, K.A.; Dhillion, S.S. Traditional herbal drugs of Bulamogi, Uganda: Plants, use and administration. J. Ethnopharmacol. 2003, 88, 19–44. [Google Scholar] [PubMed]
- Tabuti, J.R. Herbal medicines used in the treatment of malaria in Budiope county, Uganda. J. Ethnopharmacol. 2008, 116, 33–42. [Google Scholar]
- Ssegawa, P.; Kasenene, J.M. Medicinal plant diversity and uses in the Sango bay area, Southern Uganda. J. Ethnopharmacol. 2007, 113, 521–540. [Google Scholar] [CrossRef]
- Okello, J.; Ssegawa, P. Medicinal plants used by communities of Ngai Subcounty, Apac District, northern Uganda. Afr. J. Ecol. 2007, 45, 76–83. [Google Scholar]
- Tugume, P.; Kakudidi, E.K.; Buyinza, M.; Namaalwa, J.; Kamatenesi, M.; Mucunguzi, P.; Kalema, J. Ethnobotanical survey of medicinal plant species used by communities around Mabira Central Forest Reserve, Uganda. J. Ethnobiol. Ethnomed. 2016, 12, 5. [Google Scholar] [CrossRef] [PubMed]
- Oryema, C.; Bukenya Ziraba, R.; Omagor, N.; Opio, A. Medicinal plants of Erute county, Lira district, Uganda with particular reference to their conservation. Afr. J. Ecol. 2010, 48, 285–298. [Google Scholar] [CrossRef]
- Maroyi, A. An ethnobotanical survey of medicinal plants used by the people in Nhema communal area, Zimbabwe. J. Ethnopharmacol. 2011, 136, 347–354. [Google Scholar] [CrossRef]
- Maroyi, A. Traditional use of medicinal plants in south-central Zimbabwe: Review and perspectives. J. Ethnobiol. Ethnomed. 2013, 9, 31. [Google Scholar] [CrossRef] [PubMed]
- Ajaib, M.; Pullaiah, T.; Shah, S. Phytochemistry and Pharmacology of Flueggea virosa (Roxb. ex Willd.) Royle. In Bioactives and Pharmacology of Medicinal Plants, 1st ed.; Pullaiah, T., Ed.; Apple Academic Press: New York, NY, USA, 2022; Volume 1, Chapter 14; 12p, ISBN 9781003281658. [Google Scholar]
- Ouachinou, J.M.S.; Dassou, G.H.; Azihou, A.F.; Adomou, A.C.; Yédomonhan, H. Breeders’ knowledge on cattle fodder species preference in rangelands of Benin. J. Ethnobiol. Ethnomed. 2018, 14, 66. [Google Scholar]
- Omara, T.; Kagoya, S.; Openy, A.; Omute, T.; Ssebulime, S.; Kiplagat, K.M.; Bongomin, O. Antivenin plants used for treatment of snakebites in Uganda: Ethnobotanical reports and pharmacological evidences. Trop. Med. Health 2020, 48, 6. [Google Scholar] [CrossRef]
- Diallo, D.; Diakité, C.; Mounkoro, P.P.; Sangaré, D.; Graz, B.; Falquet, J.; Giani, S. [Knowledge of traditional healers on malaria in Kendi (Bandiagara) and Finkolo (Sikasso) in Mali]. Mali. Med. 2007, 22, 1–8. [Google Scholar]
- Kaou, A.M.; Mahiou-Leddet, V.; Hutter, S.; Aïnouddine, S.; Hassani, S.; Yahaya, I.; Azas, N.; Ollivier, E. Antimalarial activity of crude extracts from nine African medicinal plants. J. Ethnopharmacol. 2008, 116, 74–83. [Google Scholar]
- Singh, S.V.; Manhas, A.; Kumar, Y.; Mishra, S.; Shanker, K.; Khan, F.; Srivastava, K.; Pal, A. Antimalarial activity and safety assessment of Flueggea virosa leaves and its major constituent with special emphasis on their mode of action. Biomed. Pharmacother. 2017, 89, 761–771. [Google Scholar] [CrossRef] [PubMed]
- Kraft, C.; Jenett-Siems, K.; Siems, K.; Jakupovic, J.; Mavi, S.; Bienzle, U.; Eich, E. In vitro antiplasmodial evaluation of medicinal plants from Zimbabwe. Phytother. Res. 2003, 17, 123–128. [Google Scholar]
- Tajbakhsh, E.; Kwenti, T.E.; Kheyri, P.; Nezaratizade, S.; Lindsay, D.S.; Khamesipour, F. Antiplasmodial, antimalarial activities and toxicity of African medicinal plants: A systematic review of literature. Malar. J. 2021, 20, 349. [Google Scholar] [CrossRef] [PubMed]
- Dasgupta, S.; Roy, B.; Venkataswamy, M.; Giri, B.R. Effects of Acacia oxyphylla and Securinega virosa on functional characteristics of Raillietina echinobothrida (Phylum: Platyhelminthes; Class: Cestoidea), a poultry cestode parasite. J. Parasit. Dis. 2013, 37, 125–130. [Google Scholar] [PubMed]
- Dasgupta, S.; Giri, B.R.; Roy, B. Ultrastructural observations on Raillietina echinobothrida exposed to crude extract and active compound of Securinega virosa. Micron 2013, 50, 62–67. [Google Scholar] [CrossRef]
- Freiburghaus, F.; Ogwal, E.N.; Nkunya, M.H.; Kaminsky, R.; Brun, R. In vitro antitrypanosomal activity of African plants used in traditional medicine in Uganda to treat sleeping sickness. Trop. Med. Int. Health 1996, 1, 765–771. [Google Scholar] [CrossRef]
- Nyasse, B.; Nono, J.; Sonke, B.; Denier, C.; Fontaine, C. Trypanocidal activity of bergenin, the major constituent of Flueggea virosa, on Trypanosoma brucei. Pharmazie 2004, 59, 492–494. [Google Scholar]
- El-Hawary, S.S.; Mohammed, R.; AbouZid, S.; Zaki, M.A.; Ali, Z.Y.; Elwekeel, A.; Elshemy, H.A.H. Antitrypanosomal activity of new semi-synthetic bergenin derivatives. Chem. Biol. Drug Des. 2022, 99, 179–186. [Google Scholar]
- Elsheikh, S.H.; Bashir, A.K.; Suliman, S.M.; El Wassila, M. Toxicity of certain Sudanese Plant Extracts to Cercariae and Miracidia of Schistosoma mansoni. Int. J. Crude Drug Res. 1990, 28, 241–245. [Google Scholar] [CrossRef]
- Dickson, R.A.; Houghton, P.J.; Hylands, P.J.; Gibbons, S. Antimicrobial, resistance-modifying effects, antioxidant and free radical scavenging activities of Mezoneuron benthamianum Baill., Securinega virosa Roxb. &Wlld. and Microglossa pyrifolia Lam. Phytother. Res. 2006, 20, 41–45. [Google Scholar]
- Oghenemaro, E.F.; Oise, I.E.; Cynthia, D. The Effects of Securinega virosa Leaves on Methicillin-Resistant Staphylococcus aureus (MRSA). Int. J. Pharm. Res. Allied. Sci. 2021, 10, 29–34. [Google Scholar] [CrossRef]
- Amenu, J.D.; Neglo, D.; Abaye, D.A. Comparative Study of the Antioxidant and Antimicrobial Activities of Compounds Isolated from Solvent Extracts of the Roots of Securinega virosa. J. Biosci. Med. 2019, 7, 27–41. [Google Scholar]
- Anarado, C.E.; Anarado, C.J.O.; Umedum, N.L.; Chukwubueze, F.M.; Anarado, I.L. Phytochemical and Antimicrobial analysis of leaves of Bridelia micrantha, Cassytha filiformis, Euphorbia hirta and Securinega virosa. J. Pharmacogn. Phytochem. 2020, 9, 581–587. [Google Scholar]
- Pedersen, M.E.; Vestergaard, H.T.; Hansen, S.L.; Bah, S.; Diallo, D.; Jäger, A.K. Pharmacological screening of Malian medicinal plants used against epilepsy and convulsions. J. Ethnopharmacol. 2009, 121, 472–475. [Google Scholar] [PubMed]
- Magaji, M.G.; Anuka, J.A.; Abdu-Aguye, I.; Yaro, A.H.; Hussaini, I.M. Preliminary studies on anti-inflammatory and analgesic activities of Securinega virosa (Euphorbiaceae) in experimental animal models. J. Med. Plants Res. 2008, 2, 39–44. [Google Scholar]
- Magaji, M.G.; Yakubu, Y.; Magaji, R.A.; Musa, A.M.; Yaro, A.H.; Hussaini, I.M. Psychopharmacological potentials of methanol leaf extract of Securinega virosa Roxb (Ex Willd) Baill. in mice. Pak. J. Biol. Sci. 2014, 17, 855–859. [Google Scholar] [CrossRef]
- Aiyelero, A.M.; Abdu-Aguye, S.N.; Yaro, A.H.; Magaji, M.G. Behavioural studies on the methanol leaf extract of Securiga virosa (Euphorbiaceae) in mice. J. Pharmacogn. Phythother 2012, 4, 12–15. [Google Scholar]
- Magaji, M.G.; Yaro, A.H.; Musa, A.M.; Anuka, J.A.; Abdu-Aguye, I.; Hussaini, I.M. Sedative activity of residual aqueous fraction of Securinega virosa (Roxb. exWilld) Baill. Root bark extract in mice. Niger. J. Pharm. Sci. 2011, 10, 34–44. [Google Scholar]
- Magaji, M.G.; Mohammed, M.; Magaji, R.A.; Musa, A.M.; Abdu-Aguye, I.; Hussaini, I.M. Evaluation of the antipsychotic potential of aqueous fraction of Securinega virosa root bark extract in mice. Metab. Brain Dis. 2014, 29, 161–165. [Google Scholar] [CrossRef]
- Magaji, M.G.; Musa, A.M.; Abdullahi, M.I.; Ya’u, J.; Hussaini, I.M. Isolation of bergenin from the root bark of Securinega virosa and evaluation of its potential sleep promoting effect. Avicenna J. Phytomed 2015, 5, 587–596. [Google Scholar] [PubMed]
- Chauke, A.M.; Shai, L.J.; Mphahlele, P.M.; Mogale, M.A. Radical scavenging activity of selected medicinal plants from Limpopo province of South Africa. Afr. J. Tradit. Complement. Altern. Med. 2012, 9, 426–430. [Google Scholar] [CrossRef] [PubMed]
- Singh, J.; Kumar, A.; Sharma, A. Antianxiety activity guided isolation and characterization of bergenin from Caesalpinia digyna Rottler roots. J. Ethnopharmacol. 2017, 195, 182–187. [Google Scholar] [PubMed]
- Defeudis, G.; Mazzilli, R.; Tenuta, M.; Rossini, G.; Zamponi, V.; Olana, S.; Faggiano, A.; Pozzilli, P.; Isidori, A.M.; Gianfrilli, D. Erectile dysfunction and diabetes: A melting pot of circumstances and treatments. Diabetes Metab. Res. Rev. 2022, 38, e3494. [Google Scholar] [PubMed]
- Moshi, M.J.; Kapingu, M.C.; Uiso, F.C.; Mbwambo, Z.H.; Mahunnah, R.L. Some pharmacological properties of an aqueous extract of Securinega virosa roots. Pharm. Biol. 2000, 38, 214–221. [Google Scholar] [PubMed]
- Tanko, Y.; Okasha, M.A.; Magaji, G.M.; Yerima, M.; Yaro, A.H.; Saleh, M.I.A.; Mohammed, A. Anti-diabetic properties of Securinega virosa (Euphorbiaceae) leaf extract. Afr. J. Biotechnol. 2008, 7, 22–24. [Google Scholar]
- Deshmukh, M.V.; Ghole, V.S.; Kodam, K.M. Protective Effect Of Hydro-Ethanolic Extract Of Bushweed Flueggea virosa On Renal Damage In Streptozotocin-Induced Hyperglycemia In Rat. J. Pharm. Neg. Res. 2022, 13, 6675–6687. [Google Scholar]
- Adinortey, M.B.; Agbeko, R.; Boison, D.; Ekloh, W.; Kuatsienu, L.E.; Biney, E.E.; Affum, O.O.; Kwarteng, J.; Nyarko, A.K. Phytomedicines Used for Diabetes Mellitus in Ghana: A Systematic Search and Review of Preclinical and Clinical Evidence. Evid. Based Complement. Alternat Med. 2019, 2019, 6021209. [Google Scholar] [CrossRef] [PubMed]
- Agnihotri, P.; Kashyap, H.; Gupta, S. Natural Phyto-Active Antihyperglycemic Moieties as Preventive Therapeutics for Diabetes Type II. Plant Sci. Today 2019, 6 (Suppl. S1), 1–5. [Google Scholar] [CrossRef]
- Application of Fluevirosines A in Preparation of Blood Sugar Reducing Medicines. Chinese Patent CN103479625B; Qingdao Municipal Hospital, Qingdao, China, 20 May 2015.
- Magaji, M.G.; Anuka, J.A.; Abdu-Aguye, I.; Yaro, A.H.; Hussaini, I.M. Behavioural effects of the methanolic root bark extract of Securinega virosa in rodents. Afr. J. Tradit. Complement. Altern. Med. 2008, 5, 147–153. [Google Scholar] [CrossRef]
- Yerima, M.; Magaji, M.G.; Yaro, A.H.; Tanko, Y.; Mohammed, M.M. Analgesic and anti-inflammatory activities of the methanolic leaves extract of Securinega virosa (Euphorbiaceae). Nigerian J. Pharm. Sci. 2009, 8, 47–53. [Google Scholar]
- Ezeonwumelu, J.O.C.; Matuki, E.K.; Ajayi, A.M.; Okoruwa, A.G.; Tanayen, J.K.; Adiukwu, C.P.; Goji, A.D.T.; Dare, S.; Okonkwo, C.O.; Byarugaba, F. Phytochemical Screening, Acute Toxicity and Analgesic Properties of Aqueous Extract of Flueggea virosa’s Root in Rats. Ibnosina J. Med. Biomed. Sci. 2012, 5, 15–21. [Google Scholar]
- Dénou, A.; Haïdara, M.; Diakité, F.; Doumbia, S.; Dembélé, D.L.; Sanogo, R. Phytochemicals and Biological Activities of Flueggea virosa (Phyllanthaceae) Used in the Traditional Treatment of Benign Prostatic Hyperplasia in Mali. J. Diseases Med. Plants 2021, 7, 119–126. [Google Scholar] [CrossRef]
- Misonge, O.J.; Kamindu, G.N.; Wangui, W.S.; Muita, G.M. An ethnobotanical survey of plants used for the treatment and management of cancer in Embu County, Kenya. J. Med. Plants Stud. 2019, 7, 39–46. [Google Scholar]
- Omara, T.; Odero, M.P.; Obakiro, S.B. Medicinal plants used for treating cancer in Kenya: An ethnopharmacological overview. Bull. Nat. Res. Centre 2022, 46, 148. [Google Scholar]
- Salawu, K.M.; Oyerinde, A.A.; Ajaiyeoba, E.O. Phytochemical, antioxidant, cytotoxicity, antiproliferative and antimicrobial studies of Securinega virosa aerial parts extract. J. Pharm. Allied Sci. 2020, 17, 3296–3305. [Google Scholar]
- Ikpefan, E.O.; Ayinde, B.A.; Mudassar, A.; Farooq, A.D. Securinega virosa leaf and root bark extracts: A comparative anti-cancer study against human breast (MCF-7) and lung (NCI-H460) cancer cell lines. Nigerian J. Bot. 2020, 33, 15–23. [Google Scholar]
- Garba, M.M.; Jamilu, Y.; Muhammad, M.A.; Akpojo, A.J.; Ibrahim, A.A.; Marte, H.I. Securinega virosa (Euphorbiaceae) root bark extract inhibits glioblastoma multiforme cell survival in vitro. Afr. J. Pharm. Pharmacol. 2015, 9, 684–693. [Google Scholar]
- Ushie, O.A.; Longbab, B.D.; Kendeson, A.C.; Aasegh, T.J. Antioxidant activities of Flueggea virosa crude extracts. Trends Sci. Technol. J. 2022, 7, 270–274. [Google Scholar]
- Danlami, U.; David, B.M.; Joyce, O.O.; Olutayo, O.; Thomas, S.A. The Antioxidant Potentials and Phytochemical Properties of the Hexane, Ethyl acetate and Ethanolic Extracts of Securinega virosa (Euphorbiaceae) Leaves. J. Applied Pharm. Sci. 2013, 3, 131–133. [Google Scholar]
- Ajaib, M.; Wahla, S.Q.; Shafi, F.; Zahid, M.T.; Siddiqui, M.F.; Abbas, T. Antimicrobial and Antioxidant Screening of Flueggea virosa. Biosci. Res. 2021, 17, 2791–2798. [Google Scholar]
- Zengin, G.; Dall’Acqua, S.; Sinan, K.I.; Uba, A.I.; Sut, S.; Peron, G.; Etienne, O.K.; Kumar, M.; Cespedes-Acuña, C.L.; Alarcon-Enos, J.; et al. Gathering scientific evidence for a new bioactive natural ingredient: The combination between chemical profiles and biological activities of Flueggea virosa extracts. Food Biosci. 2022, 49, 101967. [Google Scholar] [CrossRef]
- Sanogo, R.; Vassallo, A.; Malafronte, N.; Imparato, S.; Russo, A.; Dal Piaz, F. New phenolic glycosides from Securinega virosa and their antioxidant activity. Nat. Prod. Commun. 2009, 4, 1645–1650. [Google Scholar] [CrossRef]
- de Oliveira, G.A.L.; da Silva Oliveira, G.L.; Nicolau, L.A.D.; Mafud, A.C.; Batista, L.F.; Mascarenhas, Y.P.; de Sousa, L.K.M.; David, J.M.; Pinto, L.S.; Alves, C.Q.; et al. Bergenin from Peltophorum dubium: Isolation, Characterization, and Antioxidant Activities in Non-Biological Systems and Erythrocytes. Med. Chem. 2017, 13, 592–603. [Google Scholar]
- Zhang, G.; Wang, H.; Zhang, Q.; Zhao, Z.; Zhu, W.; Zuo, X. Bergenin alleviates H2 O2 -induced oxidative stress and apoptosis in nucleus pulposus cells: Involvement of the PPAR-γ/NF-κB pathway. Environ. Toxicol. 2021, 36, 2541–2550. [Google Scholar] [CrossRef] [PubMed]
- Thiombiano, H.M.V.; Bangou, M.J.; Nacoulma, A.P.; Ouoba, B.; Sawadogo, M.; Lema, A.; Coulidiati, T.H.; Ouoba, H.Y.; Ouedraogo, G.A. Ethnobotanical survey on medicinal plants used in Burkina Faso in the treatment of breast cancer, phytochemistry and antioxidant activities: Euphorbia poissonii Pax and Flueggea virosa (Willd.) Voigt. (Euphorbiaceae). Afr. J. Biol. Med. Res. 2022, 5, 1–16. [Google Scholar]
- Pu, H.L.; Huang, X.; Zhao, J.H.; Hong, A. Bergenin is the antiarrhythmic principle of Flueggea virosa. Planta Med. 2002, 68, 372–374. [Google Scholar] [CrossRef]
- Liu, Y.; Tan, Y.; Cao, G.; Shi, L.; Song, Y.; Shan, W.; Zhang, M.; Li, P.; Zhou, H.; Zhang, B.; et al. Bergenin alleviates myocardial ischemia-reperfusion injury via SIRT1 signaling. Biomed. Pharmacother. 2023, 158, 114100. [Google Scholar] [CrossRef] [PubMed]
- Souleymane, H.D.; Djibo, A.K.; Seyni, S.H.; Zakaria, O.; Botezatu, A.V.; Dinica, R.M.; Ibrahim Maman Laouali, A.; Kouakou, N.D.V. Phytochemical Characterization and In Vitro Evaluation of the Anti-Sickle Cell Activity of Aqueous and Ethanolic Extracts of Two Medicinal Plants from Niger: Flueggea virosa (Roxb. exWilld.) Royle and Kigelia africana (Lam.) Benth. Plants 2023, 12, 3522. [Google Scholar]
- Abere, T.A.; Egharevba, C.O.; Chukwurah, I.O. Pharmacognostic evaluation and antisickling activity of the leaves of Securinega virosa Roxb. ex Willd. (Euphorbiaceae). Afr. J. Biotechnol. 2014, 13, 4040–4045. [Google Scholar]
- Kouangbé, M.A.; Bahi, C.; Tia, H.; Boga Gogo, L.; Edoh, V.; Djama, A.J.; N’Guessan, J.D. Antifungal Activity of Roots Barks Extract of Securinega virosa (Roxb. ex Willd.) Baill and Anogeissus leiocarpa (DC.) Guill. & Perr, Two Plants Used in the Traditional Treatment of Candidiasis in Northern Côte d’Ivoire. Int. J. Biochem. Res. Rev. 2015, 8, 1–11. [Google Scholar]
- Hill, L.; Holdsworth, D.; Small, R. Pharmacological investigations of virosecurinine. PNG Med. J. 1976, 18, 157–161. [Google Scholar]
- Magaji, M.G.; Yaro, A.H.; Mohammed, A.; Zezi, A.U.; Tanko, Y.; Bala, T.Y. Preliminary Antidiarrhoeal Activity of Methanolic Extracts of Securinega virosa (Euphorbiaceae). Afr. J. Biotechnol. 2007, 6, 2752–2757. [Google Scholar] [CrossRef]
- Njume, C.; Goduka, N.I. Treatment of diarrhoea in rural African communities: An overview of measures to maximise the medicinal potentials of indigenous plants. Int. J. Environ. Res. Public. Health 2012, 9, 3911–3933. [Google Scholar]
- Chinsembu, K.C.; Syakalima, M.; Semenya, S.S. Ethnomedicinal plants used by traditional healers in the management of HIV/AIDS opportunistic diseases in Lusaka, Zambia. S.Afr. J. Bot. 2019, 122, 369–384. [Google Scholar] [CrossRef]
- Zhang, H.; Zhang, C.R.; Han, Y.S.; Wainberg, M.A.; Yue, J.M. New Securinega alkaloids with anti-HIV activity from Flueggea virosa. RSC Adv. 2015, 5, 107045–107053. [Google Scholar] [CrossRef]
- Zhang, H.; Han, Y.-S.; Wainberg, M.A.; Yue, J.-M. Anti-HIV Securinega alkaloid oligomers from Flueggea virosa. Tetrahedron 2015, 71, 3671–3679. [Google Scholar] [CrossRef]
- Olaleye, T.; Muse, W.A.; Imeh-Nathaniel, A.; Nathaniel, T.I. Biological effects of Petiveria alliacea and Flueggae virosa on the life cycle of a disease vector (Musca domestica). Int. J. Pure Appl. Zool. 2017, 5, 45–51. [Google Scholar]
- Bajracharya, G.B. Diversity, pharmacology and synthesis of bergenin and its derivatives: Potential materials for therapeutic usages. Fitoterapia 2015, 101, 133–152. [Google Scholar]
- Salimo, Z.M.; Yakubu, M.N.; da Silva, E.L.; de Almeida, A.C.G.; Chaves, Y.O.; Costa, E.V.; da Silva, F.M.A.; Tavares, J.F.; Monteiro, W.M.; de Melo, G.C.; et al. Chemistry and Pharmacology of Bergenin or Its Derivatives: A Promising Molecule. Biomolecules 2023, 13, 403. [Google Scholar] [CrossRef]
- Dai, W.; Yang, J.; Liu, X.; Mei, Q.; Peng, W.; Hu, X. Anti-colorectal cancer of Ardisia gigantifolia Stapf. and targets prediction via network pharmacology and molecular docking study. BMC Complement. Med. Ther. 2023, 23, 4. [Google Scholar] [CrossRef]
- Lei, D.; Sun, Y.; Liu, J.; Chi, J. Bergenin inhibits palmitic acid-induced pancreatic β-cell inflammatory death via regulating NLRP3 inflammasome activation. Ann. Transl. Med. 2022, 10, 1058. [Google Scholar] [PubMed]
- Venkateswara Rao, B.; Pavan Kumar, P.; Ramalingam, V.; Karthik, G.; Andugulapati, S.B.; Suresh Babu, K. Piperazine tethered bergenin heterocyclic hybrids: Design, synthesis, anticancer activity, and molecular docking studies. RSC Med. Chem. 2022, 13, 978–985. [Google Scholar] [CrossRef] [PubMed]
- Jayakody, R.S.; Wijewardhane, P.; Herath, C.; Perera, S. Bergenin: A computationally proven promising scaffold for novel galectin-3 inhibitors. J. Mol. Model. 2018, 24, 302. [Google Scholar] [CrossRef] [PubMed]
- Barai, P.; Raval, N.; Acharya, S.; Borisa, A.; Bhatt, H.; Acharya, N. Neuroprotective effects of bergenin in Alzheimer’s disease: Investigation through molecular docking, in vitro and in vivo studies. Behav. Brain Res. 2019, 356, 18–40. [Google Scholar] [CrossRef]
- Madaan, R.; Singla, R.K.; Kumar, S.; Dubey, A.K.; Kumar, D.; Sharma, P.; Bala, R.; Singla, S.; Shen, B. Bergenin—A Biologically Active Scaffold: Nanotechnological Perspectives. Curr. Top. Med. Chem. 2022, 22, 132–149. [Google Scholar]
- Sharma, P.; Kumar, D.; Shri, R.; Kumar, S. Mechanistic Insights and Docking Studies of Phytomolecules as Potential Candidates in the Management of Cancer. Curr. Pharm. Des. 2022, 28, 2704–2724. [Google Scholar]
- Li, G.; Fang, Y.; Ma, Y.; Dawa, Y.; Wang, Q.; Gan, J.; Dang, J. Screening and Isolation of Potential Anti-Inflammatory Compounds from Saxifraga atrata via Affinity Ultrafiltration-HPLC and Multi-Target Molecular Docking Analyses. Nutrients 2022, 14, 2405. [Google Scholar] [CrossRef]
- Wang, G.-C.; Liang, J.-P.; Wang, Y.; Li, Q.; Ye, W.-C. Chemical constituents from Flueggea virosa. Chin. J. Nat. Med. 2008, 6, 251–253. [Google Scholar]
- Wang, X.F.; Liu, F.F.; Zhu, Z.; Fang, Q.Q.; Qu, S.J.; Zhu, W.; Yang, L.; Zuo, J.P.; Tan, C.H. Flueggenoids A–E, new dinorditerpenoids from Flueggea virosa. Fitoterapia 2019, 133, 96–101. [Google Scholar]
- Haq, K.U.; Rusdipoetra, R.A.; Siswanto, I.; Suwito, H. Elucidation of reactive oxygen species scavenging pathways of norbergenin utilizing DFT approaches. R. Soc. Open Sci. 2022, 9, 221349. [Google Scholar] [CrossRef]
- Li, W.; Cai, Z.; Schindler, F.; Bahiraii, S.; Brenner, M.; Heiss, E.H.; Weckwerth, W. Norbergenin prevents LPS-induced inflammatory responses in macrophages through inhibiting NFκB, MAPK and STAT3 activation and blocking metabolic reprogramming. Front. Immunol. 2023, 14, 1117638. [Google Scholar]
- Khan, M.S.; Khan, W.; Ahmad, W.; Singh, M.; Ahmad, S. Bergenin determination in different extracts by high-performance thin-layer chromatographic densitometry. J. Pharm. Bioallied Sci. 2015, 7, 272–274. [Google Scholar]
- Siddiqui, N.A.; Alam, P.; Al-Rehaily, A.J.; Al-Oqail, M.M.; Parvez, M.K. Simultaneous quantification of biomarkers bergenin and menisdaurin in the methanol extract of aerial parts of Flueggea virosa by validated HPTLC densitometric method. J. Chrom Sci. 2015, 53, 824–829. [Google Scholar] [CrossRef] [PubMed]
- Hussain, A.; Alam, P.; Siddiqui, N.A.; Alajmi, M.F.; Rehman, M.T.; Kalam, M.A.; Al-Rehaily, A.J. Development and validation of UPLC-PDA method for concurrent analysis of bergenin and menisdaurin in aerial parts of Flueggea virosa (Roxb. ex Willd.). Saudi Pharm. J. 2018, 26, 970–976. [Google Scholar] [CrossRef] [PubMed]
- El-Hawary, S.S.; Mohammed, R.; Abouzid, S.; Ali, Z.Y.; Elwekeel, A. Anti-arthritic activity of 11-O-(4’-O-methyl galloyl)-bergenin and Crassula capitella extract in rats. J. Pharm. Pharmacol. 2016, 68, 834–844. [Google Scholar] [CrossRef] [PubMed]
- Khan, H.; Amin, H.; Ullah, A.; Saba, S.; Rafique, J.; Khan, K.; Ahmad, N.; Badshah, S.L. Antioxidant and Antiplasmodial Activities of Bergenin and 11-O-Galloylbergenin Isolated from Mallotus philippensis. Oxid. Med. Cell Longev. 2016, 2016, 1051925. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Xie, L.; Zhou, L.; Gan, Y.; Han, S.; Zhou, Y.; Qing, X.; Li, W. Bergenin Inhibits Tumor Growth and Overcomes Radioresistance by Targeting Aerobic Glycolysis. Am. J. Chin. Med. 2023, 51, 1905–1925. [Google Scholar]
- Li, X.; Liang, Q.; Zhou, L.; Deng, G.; Xiao, Y.; Gan, Y.; Han, S.; Liao, J.; Wang, R.; Qing, X.; et al. Survivin degradation by bergenin overcomes pemetrexed resistance. Cell. Oncol. 2023, 46, 1837–1853. [Google Scholar] [CrossRef]
- Gan, Y.; Li, X.; Han, S.; Zhou, L.; Li, W. Targeting Mcl-1 Degradation by Bergenin Inhibits Tumorigenesis of Colorectal Cancer Cells. Pharmaceuticals 2023, 16, 241. [Google Scholar] [PubMed]
- Shen, M.; Li, H.; Yuan, M.; Jiang, L.; Zheng, X.; Zhang, S.; Yuan, M. Preparation of bergenin - Poly (lactic acid) polymers and in vitro controlled release studies. Int. J. Biol. Macromol. 2018, 116, 354–363. [Google Scholar]
- Ren, Y.; Shen, M.; Ding, Y.; Yuan, M.; Jiang, L.; Li, H.; Yuan, M. Study on preparation and controlled release in vitro of bergenin-amino polylactic acid polymer. Int. J. Biol. Macromol. 2020, 153, 650–660. [Google Scholar]
- de Sá Hyacienth, B.M.; Tavares Picanço, K.R.; Sánchez-Ortiz, B.L.; Barros Silva, L.; Matias Pereira, A.C.; Machado Góes, L.D.; Sousa Borges, R.; Cardoso Ataíde, R.; Dos Santos, C.B.R.; de Oliveira Carvalho, H.; et al. Hydroethanolic extract from Endopleura uchi (Huber) Cuatrecasas and its marker bergenin: Toxicological and pharmacokinetic studies in silico and in vivo on zebrafish. Toxicol. Rep. 2020, 7, 217–232. [Google Scholar] [CrossRef]
- Siddiqui, N.A.; Mothana, R.A.; Al-Rehaily, A.J.; Alam, P.; Yousaf, M.; Ahmed, S.; Alatar, A. High-performance thin-layer chromatography based concurrent estimation of biomarkers ent-phyllanthidine and rutin in the dried aerial parts of Flueggea virosa. Saudi Pharm. J. 2017, 25, 696–702. [Google Scholar] [CrossRef] [PubMed]
- Chao, C.H.; Lin, Y.J.; Cheng, J.C.; Huang, H.C.; Yeh, Y.J.; Wu, T.S.; Hwang, S.Y.; Wu, Y.C. Chemical Constituents from Flueggea virosa and the Structural Revision of Dehydrochebulic Acid Trimethyl Ester. Molecules 2016, 21, 1239. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Fisher, D.; Pronyuk, K.; Dang, Y.; Zhao, L. Agent in Urgent Need of Clinical Practice: Corilagin. Mini Rev. Med. Chem. 2023, 23, 1642–1652. [Google Scholar] [PubMed]
- Li, X.; Deng, Y.; Zheng, Z.; Huang, W.; Chen, L.; Tong, Q.; Ming, Y. Corilagin, a promising medicinal herbal agent. Biomed. Pharmacother. 2018, 99, 43–50. [Google Scholar] [CrossRef]
- Gupta, A.; Singh, A.K.; Kumar, R.; Ganguly, R.; Rana, H.K.; Pandey, P.K.; Sethi, G.; Bishayee, A.; Pandey, A.K. Corilagin in Cancer: A Critical Evaluation of Anticancer Activities and Molecular Mechanisms. Molecules 2019, 24, 3399. [Google Scholar]
- Cheng, J.C.; Chen, Y.J.; Chuang, C.W.; Chao, Y.H.; Huang, H.C.; Lin, C.C.; Chao, C.H. Polyoxygenated Terpenoids and Polyketides from the Roots of Flueggea virosa and Their Inhibitory Effect against SARS-CoV-2-Induced Inflammation. Molecules 2022, 27, 8548. [Google Scholar] [CrossRef]
- Chao, C.H.; Cheng, J.C.; Shen, D.Y.; Huang, H.C.; Wu, Y.C.; Wu, T.S. Terpenoids from Flueggea virosa and their anti-hepatitis C virus activity. Phytochemistry 2016, 128, 60–70. [Google Scholar] [CrossRef]
- de Souza, M.M.; Chagas, L.G.R.D.; Gonçalves, A.E.; Tomczak, M.; Reichert, S.; Schuquel, I.T.A.; Cechinel-Filho, V.; Meyre-Silva, C. Phytochemical Analysis and Antinociceptive Properties of Hydroalcoholic Extracts of Aleurites moluccanus Bark. Planta Med. 2021, 87, 896–906. [Google Scholar] [CrossRef]
- Qin, H.; Yu, D. 10.11 Podocarpane-type diterpenoids. In Volume 5 Diterpenoids, Triterpenoids, Sesterterpenoids, Tetraterpenoids, and Carotenoids; De Gruyter: Berlin, Germany; Boston, MA, USA, 2021; pp. 196–201. [Google Scholar] [CrossRef]
- Chao, C.H.; Cheng, J.C.; Shen, D.Y.; Wu, T.S. Anti-hepatitis C virus dinorditerpenes from the roots of Flueggea virosa. J. Nat. Prod. 2014, 77, 22–28. [Google Scholar] [CrossRef]
- Chao, C.-H.; Cheng, J.-C.; Hwang, T.-L.; Shen, D.-Y.; Wu, T.-S. Trinorditerpenes from the roots of Flueggea virosa. Bioorg Med. Chemi Lett. 2014, 24, 447–449. [Google Scholar] [CrossRef] [PubMed]
- Fischer, M.; Harms, K.; Koert, U. Total Syntheses of 7,20-Oxa-Bridged Dinorditerpenes: Antihepatitis C Virus Active (+)-Elevenol from Flueggea virosa and (+)-Przewalskin. Org. Lett. 2016, 18, 5692–5695, Erratum in Org. Lett. 2018, 20, 4158. [Google Scholar] [CrossRef] [PubMed]
- Monkodkaew, S.; Loetchutinat, C.; Nuntasaen, N.; Pompimon, W. Identification and Antiproliferative. Activity Evaluation of a Series of Triterpenoids Isolated from Flueggea virosa (Roxb. ex Willd.) Am. J. Appl. Sci. 2009, 6, 1800–1806. [Google Scholar]
- Aswathy, M.; Vijayan, A.; Daimary, U.D.; Girisa, S.; Radhakrishnan, K.V.; Kunnumakkara, A.B. Betulinic acid: A natural promising anticancer drug, current situation, and future perspectives. J. Biochem. Mol. Toxicol. 2022, 36, e23206. [Google Scholar] [CrossRef] [PubMed]
- Mu, H.; Sun, Y.; Yuan, B.; Wang, Y. Betulinic acid in the treatment of breast cancer: Application and mechanism progress. Fitoterapia 2023, 169, 105617. [Google Scholar] [CrossRef] [PubMed]
- Okunola, O.J.; Ali, T.; Mann, A.; Bello, O.M. Pharmacological and chemotaxonomic aspect of terpenes in Securinega virosa. FUDMA J. Sci. 2018, 2, 231–237. [Google Scholar]
- Kornel, A.; Nadile, M.; Retsidou, M.I.; Sakellakis, M.; Gioti, K.; Beloukas, A.; Sze, N.S.K.; Klentrou, P.; Tsiani, E. Ursolic Acid against Prostate and Urogenital Cancers: A Review of In Vitro and In Vivo Studies. Int. J. Mol. Sci. 2023, 24, 7414. [Google Scholar] [CrossRef]
- Zhao, M.; Wu, F.; Tang, Z.; Yang, X.; Liu, Y.; Wang, F.; Chen, B. Anti-inflammatory and antioxidant activity of ursolic acid: A systematic review and meta-analysis. Front. Pharmacol. 2023, 14, 1256946. [Google Scholar] [CrossRef]
- Muraveva, V.I.; Bankovskii, A.I. Chemical study of alkaloids of Securinega suffruticosa. Dokl. Akda Nauk. SSSR 1956, 110, 998–1000. [Google Scholar]
- Nakano, T.; Yang, T.H.; Terao, S. Studies on the alkaloides of Securinega virosa Pax et Hoff. I. Telmmron. Tetrahedron 1963, 19, 609–619. [Google Scholar] [CrossRef] [PubMed]
- Raj, D.; Łuczkiewicz, M. Securinega suffruticosa. Fitoterapia 2008, 79, 419–427. [Google Scholar] [PubMed]
- Chirkin, E.; Atkatlian, W.; Porée, F.H. The Securinega alkaloids. Alkaloids Chem. Biol. 2015, 74, 1–120. [Google Scholar] [PubMed]
- Park, K.J.; Kim, C.S.; Khan, Z.; Oh, J.; Kim, S.Y.; Choi, S.U.; Lee, K.R. Securinega Alkaloids from the Twigs of Securinega suffruticosa and Their Biological Activities. J. Nat. Prod. 2019, 82, 1345–1353. [Google Scholar] [PubMed]
- Hou, W.; Huang, H.; Wu, X.Q.; Lan, J.X. Bioactivities and mechanism of action of securinega alkaloids derivatives reported prior to 2022. Biomed. Pharmacother. 2023, 158, 114190. [Google Scholar]
- Liu, C.J.; Fan, X.D.; Jiang, J.G.; Chen, Q.X.; Zhu, W. Potential anticancer activities of securinine and its molecular targets. Phytomedicine 2022, 106, 154417. [Google Scholar] [CrossRef]
- Zhang, D.; Liu, H.; Yang, B.; Hu, J.; Cheng, Y. L-securinine inhibits cell growth and metastasis of human androgen-independent prostate cancer DU145 cells via regulating mitochondrial and AGTR1/MEK/ERK/STAT3/PAX2 apoptotic pathways. Biosci. Rep. 2019, 39, BSR20190469. [Google Scholar] [CrossRef]
- Ashraf, S.M.; Mahanty, S.; Rathinasamy, K. Securinine induces mitotic block in cancer cells by binding to tubulin and inhibiting microtubule assembly: A possible mechanistic basis for its anticancer activity. Life Sci. 2021, 287, 120105. [Google Scholar] [CrossRef] [PubMed]
- Klochkov, S.; Neganova, M. Unique indolizidine alkaloid securinine is a promising scaffold for the development of neuroprotective and antitumor drugs. RSC Adv. 2021, 11, 19185–19195. [Google Scholar] [CrossRef]
- Park, S.; Kang, G.; Kim, C.; Kim, D.; Han, S. Collective total synthesis of C4-oxygenated securinine-type alkaloids via stereocontrolled diversifications on the piperidine core. Nat. Commun. 2022, 13, 5149. [Google Scholar]
- Kang, G.; Park, S.; Han, S. Synthesis of High-Order and High-Oxidation State Securinega Alkaloids. Acc. Chem. Res. 2023, 56, 140–156. [Google Scholar] [CrossRef]
- Iketubosin, G.O.; Mathieson, D.W. The isolation of hordenine and norsecurinine from Securinega virosa. J. Pharm. Pharmacol. 1963, 15, 810–815. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Wei, W.; Yue, J.-M. From monomer to tetramer and beyond: The intriguing chemistry of Securinega alkaloids from Flueggea virosa. Tetrahedron 2013, 69, 3942–3946. [Google Scholar] [CrossRef]
- Zhao, B.-X.; Wang, Y.; Li, C.; Wang, G.-C.; Huang, X.-J.; Fan, C.-L.; Li, Q.-M.; Zhu, H.-J.; Chen, W.-M.; Ye, W.-C. Flueggedine, a novel axisymmetric indolizidine alkaloid dimer from Flueggea virosa. Tetrahedron Lett. 2013, 54, 4708–4711. [Google Scholar]
- Zhang, H.; Zhang, C.R.; Zhu, K.K.; Gao, A.H.; Luo, C.; Li, J.; Yue, J.M. Fluevirosines A-C: A biogenesis inspired example in the discovery of new bioactive scaffolds from Flueggea virosa. Org. Lett. 2013, 15, 120–123. [Google Scholar] [CrossRef]
- Li, X.-H.; Cao, M.-M.; Zhang, Y.; Li, S.-L.; Di, Y.-T.; Hao, X.-J. Fluevirines A-D, four new securinega-type alkaloids from Flueggea virosa. Tetrahedron Lett. 2014, 55, 6101–6104. [Google Scholar]
- Yang, X.; Liu, J.; Huo, Z.; Yuwen, H.; Li, Y.; Zhang, Y. Fluevirines E and F, two new alkaloids from Flueggea virosa. Nat. Prod. Res. 2020, 34, 2001–2006. [Google Scholar]
- Xie, Q.J.; Zhang, W.Y.; Wu, Z.L.; Xu, M.T.; He, Q.F.; Huang, X.J.; Che, C.T.; Wang, Y.; Ye, W.C. Alkaloid constituents from the fruits of Flueggea virosa. Chin. J. Nat. Med. 2020, 18, 385–392. [Google Scholar]
- Zhao, B.X.; Wang, Y.; Zhang, D.M.; Jiang, R.W.; Wang, G.C.; Shi, J.M.; Huang, X.J.; Chen, W.M.; Che, C.T.; Ye, W.C. Flueggines A and B, two new dimeric indolizidine alkaloids from Flueggea virosa. Org. Lett. 2011, 13, 3888–3891. [Google Scholar] [CrossRef]
- Wei, H.; Qiao, C.; Liu, G.; Yang, Z.; Li, C.C. Stereoselective total syntheses of (-)-flueggine A and (+)-virosaine B. Angew. Chem. Int. Ed. Engl. 2013, 52, 620–624. [Google Scholar] [CrossRef]
- Gan, L.S.; Fan, C.Q.; Yang, S.P.; Wu, Y.; Lin, L.P.; Ding, J.; Yue, J.M. Flueggenines A and B, two novel C,C-linked dimeric indolizidine alkaloids from Flueggea virosa. Org. Lett. 2006, 8, 2285–2288. [Google Scholar]
- Jeon, S.; Lee, J.; Park, S.; Han, S. Total synthesis of dimeric Securinega alkaloids (-)-flueggenines D and I. Chem. Sci. 2020, 11, 10934–10938. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Han, Y.-S.; Wainberg, M.A.; Yue, J.-M. Flueggethers B-D, Securinega alkaloids with rare oligomerizing pattern from Flueggea virosa. Tetrahedron Lett. 2016, 57, 1798–1800. [Google Scholar]
- Luo, X.K.; Cai, J.; Yin, Z.Y.; Luo, P.; Li, C.J.; Ma, H.; Seeram, N.P.; Gu, Q.; Xu, J. Fluvirosaones A and B, Two Indolizidine Alkaloids with a Pentacyclic Skeleton from Flueggea virosa. Org. Lett. 2018, 20, 991–994. [Google Scholar]
- Gan, L.S.; Yue, J.M. Alkaloids from the Root of Flueggea virosa. Nat. Prod. Commun. 2006, 1, 819–823. [Google Scholar] [CrossRef]
- Miyatake-Ondozabal, H.; Bannwart, L.M.; Gademann, K. Enantioselective total synthesis of virosaine A and bubbialidine. Chem. Commun. (Camb) 2013, 49, 1921–1923. [Google Scholar] [CrossRef] [PubMed]
- Luger, P.; Weber, M.; Dung, N.X.; Ky, P.T.; Long, P.K. Securinine, an Alkaloid from Fluggea virosa. Acta Cryst. 1995, C51, 127–129. [Google Scholar]
- Alibés, R.; Bayón, P.; de March, P.; Figueredo, M.; Font, J.; García-García, E.; González-Gálvez, D. An effective enantioselective approach to the securinega alkaloids: Total synthesis of (-)-norsecurinine. Org. Lett. 2005, 7, 5107–5109. [Google Scholar] [CrossRef]
- González-Gálvez, D.; García-García, E.; Alibés, R.; Bayón, P.; de March, P.; Figueredo, M.; Font, J. Enantioselective approach to Securinega alkaloids. Total synthesis of securinine and (-)-norsecurinine. J. Org. Chem. 2009, 74, 6199–6211. [Google Scholar] [CrossRef]
- Zhang, H.; Zhu, K.-K.; Gao, X.-H.; Yue, J.-M. Natural occurrence of all eight stereoisomers of a neosecurinane structure from Flueggea virosa. Tetrahedron 2017, 73, 4692–4697. [Google Scholar] [CrossRef]
- Wehlauch, R.; Grendelmeier, S.M.; Miyatake-Ondozabal, H.; Sandtorv, A.H.; Scherer, M.; Gademann, K. Investigating Biogenetic Hypotheses of the Securinega Alkaloids: Enantioselective Total Syntheses of Secu’amamine E/ent-Virosine A and Bubbialine. Org. Lett. 2017, 19, 548–551. [Google Scholar] [CrossRef]
- Ma, N.; Yao, Y.; Zhao, B.X.; Wang, Y.; Ye, W.C.; Jiang, S. Total synthesis of securinega alkaloids (-)-norsecurinine, (-)-niruroidine and (-)-flueggine A. Chem. Commun. 2014, 50, 9284–9287. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.-Y.; Wang, A.-T.; Zhao, B.-X.; Lei, X.-P.; Zhang, D.-M.; Jiang, R.-W.; Wang, Y.; Ye, W.-C. Norsecurinamines A and B, two norsecurinine-derived alkaloid dimers from the fruits of Flueggea virosa. Tetrahedron Lett. 2016, 57, 3810–3813. [Google Scholar] [CrossRef]
- Dehmlow, E.V.; Guntenhöner, M.; Ree, T.V. A novel alkaloid from Flueggea virosa: 14,15-epoxynorsecurinine. Phytochemistry 1999, 52, 1715–1716. [Google Scholar] [CrossRef]
- Tatematsu, H.; Mori, M.; Yang, T.H.; Chang, J.J.; Lee, T.T.; Lee, K.H. Cytotoxic principles of Securinega virosa: Virosecurinine and viroallosecurinine and related derivatives. J. Pharm. Sci. 1991, 80, 325–327. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.-C.; Wang, Y.; Li, Q.; Liang, J.-P.; Zhang, X.-Q.; Yao, X.-S.; Ye, W.-C. Two new alkaloids from Flueggea virosa. Helvet Chim. Acta 2008, 91, 1124–1129. [Google Scholar] [CrossRef]
- Zhao, B.-X.; Wang, Y.; Zhang, D.-M.; Huang, X.-J.; Bai, L.-L.; Yan, Y.; Chen, J.-M.; Lu, T.-B.; Wang, Y.-T.; Zhang, Q.-W.; et al. Virosaines A and B, two new birdcage-shaped Securinega alkaloids with an unprecedented skeleton from Flueggea virosa. Org. Lett. 2012, 14, 3096–3099. [Google Scholar]
- Sertel, S.; Fu, Y.; Zu, Y.; Rebacz, B.; Konkimalla, B.; Plinkert, P.K.; Krämer, A.; Gertsch, J.; Efferth, T. Molecular docking and pharmacogenomics of vinca alkaloids and their monomeric precursors, vindoline and catharanthine. Biochem. Pharmacol. 2011, 81, 723–735. [Google Scholar] [CrossRef]
- Strömbom, J.; Jokela, R.; Saano, V.; Rolfsen, W. Binding of strychnocarpine and related beta-carbolines to brain receptors in vitro. Eur. J. Pharmacol. 1992, 214, 165–168. [Google Scholar] [CrossRef]
- Schmid, C.L.; Bohn, L.M. Serotonin, but not N-methyltryptamines, activates the serotonin 2A receptor via a ß-arrestin2/Src/Akt signaling complex in vivo. J. Neurosci. 2010, 30, 13513–13524. [Google Scholar] [CrossRef]
- Vergoten, G.; Bailly, C. Molecular Docking of Cryptoconcatones to α-Tubulin and Related Pironetin Analogues. Plants 2023, 12, 296. [Google Scholar] [PubMed]
- Vergoten, G.; Bailly, C. The Plagiochilins from Plagiochila Liverworts: Binding to α-Tubulin and Drug Design Perspectives. AppliedChem 2023, 3, 217–228. [Google Scholar]
- Al-Rehaily, A.J.; Yousaf, M.; Ahmed, M.S.; Samoylenko, V.; Li, X.C.; Muhammad, I.; El Tahir, K.E.H. Chemical and biological study of Flueggea virosa native to Saudi Arabia. Chem. Nat. Compounds 2015, 51, 187–188. [Google Scholar] [CrossRef]
- Muhammad, A.; Sirat, H.M. COX-2 inhibitors from stem bark of Bauhinia rufescens Lam. (Fabaceae). EXCLI J. 2013, 12, 824–830. [Google Scholar] [PubMed]
- Krzyżanowska-Kowalczyk, J.; Kowalczyk, M.; Ponczek, M.B.; Pecio, Ł.; Nowak, P.; Kolodziejczyk-Czepas, J. Pulmonaria obscura and Pulmonaria officinalis Extracts as Mitigators of Peroxynitrite-Induced Oxidative Stress and Cyclooxygenase-2 Inhibitors-In Vitro and In Silico Studies. Molecules 2021, 26, 631. [Google Scholar] [CrossRef]
- Peng, Y.L.; Zeng, N.; Yao, Q.Y.; Peng, C.Y.; Sheng, W.B.; Li, B.; Wang, W. A Review of the Medicinal Uses, Phytochemistry and Pharmacology of Genus Flueggea. Current Chinese Sci. 2023, 3, 213–241. [Google Scholar] [CrossRef]
- Fouotsa, H.; Le Pogam, P.; Mkounga, P.; Lannang, A.M.; Bernadat, G.; Vanheuverzwijn, J.; Zhou, Z.; Leblanc, K.; Rharrabti, S.; Nkengfack, A.E.; et al. Voatriafricanines A and B, Trimeric Vobasine-Aspidosperma-Aspidosperma Alkaloids from Voacanga africana. J. Nat. Prod. 2021, 84, 2755–2761. [Google Scholar]
- Ma, X.; Chen, H.; Zhu, S.; Tu, P.; Jiang, Y. Trimeric and Dimeric Carbazole Alkaloids from Murraya microphylla. Molecules 2021, 26, 5689. [Google Scholar]
- Wang, Y.; Ma, S.G.; Li, L.; Yu, S.S. Indole alkaloids from the bark of Acacia confusa and their potential antinociceptive and anti-inflammatory activities. J. Asian Nat. Prod. Res. 2022, 24, 1109–1127. [Google Scholar] [CrossRef]
- Kren, V.; Fiserová, A.; Weignerová, L.; Stibor, I.; Halada, P.; Prikrylová, V.; Sedmera, P.; Pospísil, M. Clustered ergot alkaloids modulate cell-mediated cytotoxicity. Bioorg Med. Chem. 2002, 10, 415–424. [Google Scholar] [CrossRef]
- Kren, V.; Eich, E.; Pertz, H.H. Pergolide, terguride and N,N’-spacer-linked oligomers of both interact with 5-HT2A receptors of rat tail artery. Physiol. Res. 2004, 53, 35–43. [Google Scholar] [CrossRef]
- Tang, G.; Liu, X.; Ma, N.; Huang, X.; Wu, Z.L.; Zhang, W.; Wang, Y.; Zhao, B.X.; Wang, Z.Y.; Ip, F.C.; et al. Design and Synthesis of Dimeric Securinine Analogues with Neuritogenic Activities. ACS Chem. Neurosci. 2016, 7, 1442–1451. [Google Scholar] [CrossRef] [PubMed]
- Hou, W.; Wang, Z.Y.; Lin, J.; Chen, W.M. Induction of differentiation of the acute myeloid leukemia cell line (HL-60) by a securinine dimer. Cell Death Discov. 2020, 6, 123. [Google Scholar] [CrossRef] [PubMed]
- Liao, Y.; Zhuang, X.; Huang, X.; Peng, Y.; Ma, X.; Huang, Z.X.; Liu, F.; Xu, J.; Wang, Y.; Chen, W.M.; et al. A Bivalent Securinine Compound SN3-L6 Induces Neuronal Differentiation via Translational Upregulation of Neurogenic Transcription Factors. Front. Pharmacol. 2018, 9, 290. [Google Scholar] [CrossRef] [PubMed]
- Dickson, R.A.; Houghton, P.J.; Govindarajan, R. In-vitro and in-vivo wound healing properties of two plants from Ghana. Planta Med. 2007, 73, 465. [Google Scholar]
Figure 1.
The plant Flueggea virosa (Roxb. ex Willd.) Royle, also known as Securinega virosa (Roxb. ex Willd.) Baill. (a–e) Twigs, leaves and fruit of F. virosa. (f) This Flueggea species is mostly distributed in Africa, southeast Asia and Oceania. (g) Drawing of the bush.
Figure 1.
The plant Flueggea virosa (Roxb. ex Willd.) Royle, also known as Securinega virosa (Roxb. ex Willd.) Baill. (a–e) Twigs, leaves and fruit of F. virosa. (f) This Flueggea species is mostly distributed in Africa, southeast Asia and Oceania. (g) Drawing of the bush.

Figure 2.
Pharmacological properties reported with extracts of F. virosa.

Figure 3.
(a) Structures of bergenin, norbergenin and 11-O-caffeoylbergenin isolated from the leaves of F. virosa. (b) Molecular model of bergenin bound to ESR-1 (estrogen receptor 1), with two major hydrogen bonds between the drug and residues Thr347 and His524 of ESR-1 [128].
Figure 3.
(a) Structures of bergenin, norbergenin and 11-O-caffeoylbergenin isolated from the leaves of F. virosa. (b) Molecular model of bergenin bound to ESR-1 (estrogen receptor 1), with two major hydrogen bonds between the drug and residues Thr347 and His524 of ESR-1 [128].

Figure 4.
Podocarpane-type diterpenoids found in F. virosa. Compounds 4 and 8 correspond to 3β,12-dihydroxy-13-methylpodocarpa-6,8,11,13-tetraene [152] and 3α-hydroxy-12-methoxy-13-methyl-ent-podocarpa-8,11,13-triene [157], respectively.

Figure 5.
Monomeric and oligomeric alkaloids isolated from F. virosa.

Figure 6.
Other alkaloids found in F. virosa.

Figure 7.
Alkaloids isolated from the fruit of F. virosa.

Figure 8.
Molecular model of flueggine B (Flg-B) bound to α/β-tubulin dimer (PDB: 5FNV). (a) A surface model of α/β-tubulin with bound Flg-B. The α-helices (in red) and β-sheets (in cyan) are shown. (b) A close-up view of Flg-B bound to the α/β-tubulin interface with the solvent-accessible surface (SAS) surrounding the drug-binding zone (color code indicated). (c) A detailed view of the Flg-B binding site. (d,e) Binding map contacts for securinine and Flg-B bound to α-tubulin (color code indicated). The docking analysis was performed as recently described using the GOLD docking procedure and the following flexible amino acids around the binding cavity: Phe135, Ser135, Phe169, Cys200, Phe202, Ser241, Leu242, Phe255, Cys316 and Lys 352 [210,211].
Figure 8.
Molecular model of flueggine B (Flg-B) bound to α/β-tubulin dimer (PDB: 5FNV). (a) A surface model of α/β-tubulin with bound Flg-B. The α-helices (in red) and β-sheets (in cyan) are shown. (b) A close-up view of Flg-B bound to the α/β-tubulin interface with the solvent-accessible surface (SAS) surrounding the drug-binding zone (color code indicated). (c) A detailed view of the Flg-B binding site. (d,e) Binding map contacts for securinine and Flg-B bound to α-tubulin (color code indicated). The docking analysis was performed as recently described using the GOLD docking procedure and the following flexible amino acids around the binding cavity: Phe135, Ser135, Phe169, Cys200, Phe202, Ser241, Leu242, Phe255, Cys316 and Lys 352 [210,211].


{kind=link}
{kind=link}
{kind=link}
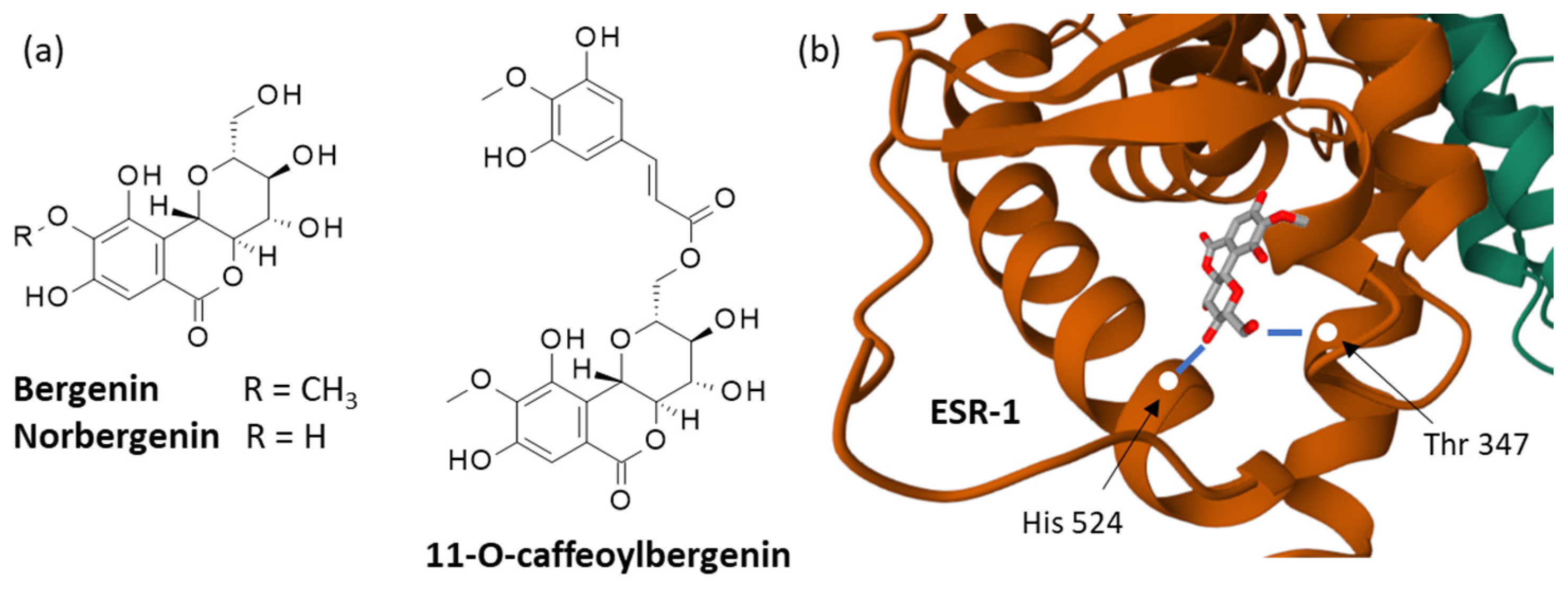{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Accepted names of Flueggea species (Phyllanthaceae) 1.
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
1 According to https://www.worldfloraonline.org/ (accessed on 15 December 2023).
Table 2.
Vernacular names given to Flueggea virosa (and synonyms) a.
| Country (Region or Province) | Vernacular Names (Language) | References |
|---|---|---|
| English names | White berry bush, snowberry tree, Chinese waterberry, simpleleaf bushweed, common bushweed. | (a) |
| Benin (Cotonou, Abomey-Calavi) | Gaagah, tchian tchian, sian sian (Bariba); yéri kalawunfa (Berba); adjaya, gbyihountin, hétré, hounsividjayé, tchaké tchaké (Fon, Goun); koukrinou (Gourmantché); tchamanoudira (Kotokoli); hesré (Watchi); igi agbado, irandjé, wadjidji (Watchi); hesre (Fon) | [12] |
| Burkina Faso (Ouagadougou) (Niangoloko forest) | Sûgdendaaqa (Mossi), fiŋyaŋma (Goin), sï’ngnamâ (Goin) | [13,14] |
| Burundi (Zaïre-Nil peaks region and Western Burundi) | Umubwirrwa, umubwiga, umujisharugi (Kirundi) | (a) |
| Cameroon (northern region) | Tiami (Fulfuldé) | [15] |
| Central African Republic (Gbaya dialect, Bossangoa) | Marro | [16] |
| Comores | M’haniba (Grande Comores) | (a) |
| Democratic Republic of Congo (Itury forest) | Njima (Mbuti pygmies) | [17] |
| Ethiopia (Omo Zone), (Arsi Zone) | Tania, qacaaculee | [18,19] |
| Fiji | Pou | [1] |
| France | Balan des savanes | (a) |
| Ghana (Dangme West district) | Asre (Adangbe) | [20] |
| India | Perimklavu, vellapoolam (Malayalam); pithondi | [21,22] |
| Ivory Cost | Môgôcolo-calaman (Dioula); mokokoana, mokrodoma (Malinké) | [23] |
| Kenya (Bungoma district), (Kilifi district) | Lubwili (Bukusu); mukwamba (Giriama); kiptarpotich, chepochepkai (Pokot); mkwamba (Swahili); mukuluu (Kamba); lkirebuki (Dorobo) (Suiei) | [24,25,26,27,28,29] |
| Madagascar | Kotika (Antakarana); atsikana, fantsikakoholahy, patikakoho, teto, voafotsikotikana (Malagasy) | [30] |
| Mali (Dogonland) (Niono district) | Jene, surukujè (Bambara); nginnin (Malinké); jene (Minyanka); jeme (Sénoufo); sutèmi, sudèrèmi (Bwa); sumenh (Bobo-fing); segele (Dogon); déné, nkoloningé, baram baram, karam karam (Bambara); kolonidiè (Bamanan); ndjene (Bambara); segedere, ségélé, sèsègèrè, balambalam | [31,32,33,34,35] |
| Mozambique (Massingir district) | Nsangasi, sangasi (Changana) | [36] |
| Niger | Fulasco (Hausa), kamal (Peuhl), dagkirto (Béribéri), tsa | [37] |
| Nigeria | Iranje (Yoruba), tsuwawun karee (Hausa), njisinta (igbo) | [38,39] |
| Nigeria, Benin, Togo | Iranje (Yoruba people) | (a) |
| Philippines | Anislag or anislog, (Bikol, Cebu Bisaya, Samar-Leyte Bisaya, Manobo, Mandaya); anislang, hamislag (Bikol); katamangan, malangau (Manobo); tras (Magindanao) | [1] |
| Samoa | Poumuli | [1] |
| Senegal (Saloum Islands) | Sambelgorel, tembelforel (Peul); baram baram, dene, balamanten, tene (Bambara); mbarambaram, faragfarag, maymayin (Sérère); keng (Wolof); fusabel fu nene, e buker, ba herer (Diola); bouroum baran (Socé); tembel gorel (Peul, Toucouleur) | [33,40,41] |
| Solomon Islands | Mamufu’a or Mamufua (Kwara’ae), mewana (Bougainville Island) | [1] |
| Somalia | Xararaay | [42] |
| South Africa (Northern Maputaland), (Vhavenda), (Limpopo province), (Mpumalanga province), (Limpopo province) | Ihlalanyosi (Zulu); mutangauma (Venda); witbessiesbos (Afr.); mutangahuma (Tshivenda); mpfalambati (Tsonga); muhlakaume (Sotho); motlhalabu, motlhakawume, mohlakauma | [43,44,45,46] |
| Tanzania, Kenya, Mozambique (Swahili) | Mkwamba, mkwamba maji, mteja | [47] |
| Tanzania (Katoro Ward, Bukoba District), (Lushoto and Korogwe districts), (Handeni district), (Tanga district), (Kimboza forest reserve in Morogoro), (Muleba district) | Mkwambikwamb, lukwambikwambi (Kibunga); omubwera, omutoruka, muumbiti, muhumba, mkwamba, mtulavuha, msokote, mkalananga, mturuka, eruati, mkwambe (Ndengereko, Swahili, Zaramo) | [48,49,50,51,52] |
| Togo (Maritime region) | Kébantchalé, kébantialé, kégbantilé, akassélem, hesré (Mina, Ewé); tchakatchaka (Tem, Kabiyé, Fè, Adélé); tchacatchaca (Tem); hesre | [53] |
| Uganda (Budiope county), (Sango bay area), (Apac district), (Mabira Central Forest Reserve), (Langi language, Erute county, Lira district) | Lukandwa, olukandwa, iakara, ilakara, omukarara (Runyaruguru); omukalali (Rukonjo); lukandwa (Luganda) | [54,55,56,57,58,59,60] |
| Vanuatu | Mamau (Butu), nemameiu (Hiu) | [1] |
| Wallis and Futuna | Poutea | [1] |
| Zimbabwe (Harare and Nhema area, south-central region) | Muchagawuwe (Shona), umhakawuwe (Ndebele) | [61,62] |
| Zambia (Lusaka) | Lukuswaula | [1] |
a http://www.ethnopharmacologia.org/recherche-dans-prelude/?plant_id=5358 (accessed on 15 December 2023).
Table 3.
Synonyms for Flueggea virosa (Roxb. ex Willd.) Royle (1836) 1.
|
|
|
|
|
|
|
|
|
|
|
|
1 According to https://www.worldfloraonline.org/ (accessed on 15 December 2023).
Table 4.
Alkaloids isolated from F. virosa and their properties.
| Compounds | Plant Parts Used (Location of Plant Collection) | Studies and Properties | References |
|---|---|---|---|
| Fluevirosinines A–D | Twigs and leaves (Guangxi, China) | Isolation, structural characterization, proposed biosynthetic pathway. Dimers, trimer, tetramer. | [182] |
| Fluevirosinines B–J | Twigs and leaves (Guangxi, China) | Isolation, structural characterization, proposed biosynthetic pathway. Inhibitory effect of fluevirosinine B on HIV-1 infected MT4 cells (EC50 = 14.1 μM), without cytotoxicity. Tetramers and pentamers. | [124] |
| Fluevirosines A–C | Twigs and leaves (China) | Isolation, structural characterization, biosynthetic pathway. No antiproliferative activity, but a weak inhibition (35% at 20 µM) of the splicing activity of XBP1 mRNA. | [184] |
| Fluevirosine D | Twigs and leaves (Guangxi, China) | [182] | |
| Fluevirines A–D | Twigs and leaves (China) | Isolation, structural characterization. Activity against Staphylococcus aureus. Fluevirine A showed activity against (MIC = 15.3 µg/mL). | [185] |
| Fluevirines E and F | Twigs and leaves (Xishuangbanna, Yunnan, China) | Very modest inhibition of cancer cell proliferation (<40% inhibition with fluevirine F at 40 µM). | [186] |
| Flueindolines A–C | Ripe fruit (Guangxi, China) | Isolation, structural characterization. Tested for neuroprotective activities on Neuro-2a cells and antivirus effects against the respiratory syncytial virus (RSV), but was found inactive. | [187] |
| Flueggines A and B | Twigs and leaves (China) | Isolation, structural characterization. Potent inhibition of cancer cell proliferation. Flueggine B strongly inhibited growth of MCF-7 and MDA-MB-231 cells (IC50 = 135 and 147 nM, respectively), whereas flueggine B was much less active. Total synthesis of flueggine A. | [188,189] |
| Flueggenines A and B | Roots (China) | Weak antiproliferative activity of flueggenine A against P388 leukemia cells (IC50 = 51.5 µM). | [190] |
| Flueggenines C and D | Twigs and leaves (Guangxi, China) | Isolation, structural characterization. Flueggenine D inhibited HIV-1 infection of MT4 cells (EC50 = 7.8 μM) and was weakly cytotoxic (selectivity index = 12.6). | [123,182] |
| Flueggenines E–I | Twigs and leaves (Xishuangbanna, Yunnan, China) | Isolation, structural characterization. Mild inhibition of MT4 cell infection by HIV-1 (EC50 = 42.6 μM) with flueggenine E. Enantioselective synthesis of flueggenines D and I. | [123,191] |
| Flueggedine | Twigs and leaves (China) | Isolation, structural characterization, proposed biosynthetic pathway from virosecurinine. No antiproliferative activity. | [183] |
| Flueggether A Virosinine A | Stems and leaves (China) | Isolation, structural characterization. Mild inhibitory effect on HIV-1-infected MT4 cells (EC50 = 120 and 45 μM, respectively) without cytotoxicity. | [6] |
| Flueggethers B–D | Twigs and leaves (Guangxi, China) | Isolation, structural characterization. Dimers (B–D), trimer (D). Inactive against tyrosine kinases c-Met and FGFR1. Flueggether D showed mild anti-HIV activity (CC50 = 43.1 μM) on MT-4 cells and was not cytotoxic (IC50 > 100 μM). | [192] |
| Fluvirosaones A and B | Twigs and leaves (China) | Isolation, structural characterization, biosynthetic pathway. Lipid-lowering effects in 3T3-L1 cells: weaker inhibition of triglyceride cellular accumulation compared with virosecurinine. | [193] |
| Bubbialine, Bubbialidine | Roots (Hainan, China) | Isolation. Total synthesis of bubbialidine. | [194,195] |
| Securinine, Norsecurinine | All plant parts | Total synthesis. Crystalline structure. | [196,197,198] |
| Securinol A Episecurinol A Secu’amamine E | Twigs and leaves (Guangxi, China) | Isolation, structural characterization. Total synthesis of secu’amamine E (and bubbialine). | [199,200] |
| Norsecurinine Norsecurinic acid Niruroidine | Roots (Hainan, China) | Isolation, structural characterization. (-)-Norsecurinine was found inactive against cancer cells and several bacteria. Total synthesis. | [194,201] |
| Norsecurinamines A and B | Fruit (China) | Isolation, structural characterization, proposed biosynthetic pathway from tryptophane. | [202] |
| Epoxynorsecurinine | Bark (Venda, South Africa) | Isolation, structural characterization. | [203] |
| ent-phyllanthidine ((+)-phyllanthidine) | Aerial parts (Saudi Arabia) | Quantification from a methanolic extract (biomarker). | [149] |
| Virosecurinine Viroallosecurinine | Leaves (Taiwan) | Isolation, structural characterization. Cytotoxic activity against cancer cell lines (EC50 = 3–8 μM). | [204] |
| Virosines A and B | Twigs and leaves (Guangdong, China) | Isolation, structural characterization. The two products and their stereoisomers. | [199,205] |
| Virosaines A and B | Twigs and leaves (China) | Isolation, structural characterization. Inactive against different cancer cell lines. Total synthesis of virosaines A and B. | [189,195,206] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Bailly, C. Traditional Uses, Pharmacology and Phytochemistry of the Medicinal Plant Flueggea virosa (Roxb. ex Willd.) Royle. Future Pharmacol. 2024, 4, 77-102. https://doi.org/10.3390/futurepharmacol4010007
AMA Style
Bailly C. Traditional Uses, Pharmacology and Phytochemistry of the Medicinal Plant Flueggea virosa (Roxb. ex Willd.) Royle. Future Pharmacology. 2024; 4(1):77-102. https://doi.org/10.3390/futurepharmacol4010007
Chicago/Turabian StyleBailly, Christian. 2024. "Traditional Uses, Pharmacology and Phytochemistry of the Medicinal Plant Flueggea virosa (Roxb. ex Willd.) Royle" Future Pharmacology 4, no. 1: 77-102. https://doi.org/10.3390/futurepharmacol4010007