
The Invasive Mechanisms of the Noxious Alien Plant Species Bidens pilosa


1
Department of Applied Biological Science, Faculty of Agriculture, Kagawa University, Miki, Kagawa 761-0795, Japan
2
Department of Agronomy, Faculty of Agriculture, Universitas Padjadjaran, Jalan Raya Bandung Sumedang Km 21, Jatinangor, Sumedang 45363, Jawa Barat, Indonesia
*
Authors to whom correspondence should be addressed.
Plants 2024, 13(3), 356; https://doi.org/10.3390/plants13030356
Submission received: 7 December 2023
/
Revised: 12 January 2024
/
Accepted: 12 January 2024
/
Published: 25 January 2024
(This article belongs to the Special Issue Plant Invasion 2023)
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Bidens pilosa L. is native to tropical America and has widely naturized from tropical to warm temperate regions in Europe, Africa, Asia, Australia, and North and South America. The species has infested a wide range of habitats such as grasslands, forests, wetlands, streamlines, coastal areas, pasture, plantations, agricultural fields, roadsides, and railway sides and has become a noxious invasive weed species. B. pilosa forms thick monospecific stands, quickly expands, and threatens the indigenous plant species and crop production. It is also involved in pathogen transmission as a vector. The species was reported to have (1) a high growth ability, producing several generations in a year; (2) a high achene production rate; (3) different biotypes of cypselae, differently germinating given the time and condition; (4) a high adaptative ability to various environmental conditions; (5) an ability to alter the microbial community, including mutualism with arbuscular mycorrhizal fungi; and (6) defense functions against natural enemies and allelopathy. The species produces several potential allelochemicals such as palmitic acid, p-coumaric acid, caffeic acid, ferulic acid, p-hydroxybenzoic acid, vanillic acid, salycilic acid, quercetin, α-pinene, and limonene and compounds involved in the defense functions such as 1-phenylhepta-1,3,5-trine, 5-phenyl-2-(1-propynyl)-thiophene, 5-actoxy-2-phenylethinyl-thiophene, and icthyothereol acetate. These characteristics of B. pilosa may contribute to the naturalization and invasiveness of the species in the introduced ranges. This is the first review article focusing on the invasive mechanisms of the species.
1. Introduction
Bidens pilosa L., belonging to the Asteraceae family, is an annual (or biennial) herbaceous plant. The species grows 20–180 cm tall, and the stems are quadrangular with hairy straggling branches. It has alternate leaves with 3–5 pinnate leaflets, which are supported by a petiole (10–70 mm long). The leaflets are broadly ovate, serrate, and 30–70 mm long and 12–18 mm wide. Capitula occur at the end of the main stems and lateral branches and expand 5–12 mm in diameter. Capitula consist of 0–8 ray florets and 35–55 disk florets. The corollas of the ray florets are 7–15 mm long and white–yellow. The ray florets have poorly developed pistils and lack stamens. The disk florets have 3–5 mm long yellow corollas, five stamens, and well-developed 2–3 mm long pistils. Its fruits are black liner cypselae with 2–5 stiff awns of 2–4 mm long [1,2,3,4,5]. The species often forms thick monospecific stands [1,2,3] (Figure 1).
B. pilosa has been used as folk medicine in the treatment of various diseases such as fever, diarrhea, hepatitis, snake bite, and wounds [6,7] and as a nutritious vegetable [8,9,10,11]. Recent investigations showed B. pilosa has a wide range of pharmacological activity such as for malaria, cancer, diabetes, inflammation, and hypertension [12,13,14]. The leaves also contain high levels of protein [11,15].
Despite the pharmacological potential, B. pilosa is also known as a noxious invasive weed species. The species is native to tropical America and has naturized in over 60 countries of tropical, subtropical, and warm temperate regions in Europe, Africa, Asia, Australia, and North and South America [1,2,3]. Many of the invasive plant species potentially threaten the native flora and fauna in the introduced ranges [16,17,18]. B. pilosa has infested a wide range of habitats such as grasslands, forest margins, secondary forests, wetlands, streamlines, coastal areas, roadsides, railway sides, disturbed lands, pasture, plantations, and agriculture fields [1,2,19,20,21]. The species has also infested several islands such as Hawaii, Fiji, and the Cook Islands [22]. The ecosystems of these islands are very vulnerable to alien species [23,24]. B. pilosa is listed as a noxious agricultural and environmental weed in more than 40 countries [1,2,19,20,21]. According to the accumulated publications, the species is highly invasive and potentially causes serious negative impacts on natural ecosystems and agricultural crop production. However, there is no review article of the impacts and invasive mechanisms of B. pilosa. This paper provides an overview of the literature describing the impacts and invasive characteristics and mechanisms of the species. The species B. pilosa is divided into some varieties such as B. Pilosa var. minor, var. pilosa, and var. radiata [25,26]. Although some authors distinguished the varieties of B. pilosa for their investigations, many researchers did not distinguish them. Therefore, this paper describes the species as B. pilosa because it is impossible to confirm the variety of the species for their investigations. The literature was searched using a combination of the predominant online search engines, Scopus, ScienceDirect, and Google Scholar, and all possible combinations of B. pilosa with the following words: biology, invasive mechanism, habitat, impact, reproduction, flower, seed, plasticity, adaptation, nutrient, colonization, arbuscular mycorrhizal fugus, rhizobium, allelopathy, allelochemical, allelopathic substance, defense function, natural enemy, insecticidal activity, fungicidal activity, pharmacology, second metabolite, and global warming.
2. Impacts of B. pilosa
B. pilosa has the potential to rapidly grow and to form dense thickets. The species outcompetes crops in agricultural fields and eliminates indigenous plant species in introduced ranges by expanding the margins of its dense thickets [1,2,3,19,20,27]. B. pilosa was reported to replace the indigenous plant species of islands such as Panicum fauriei Hitcc. and Scaevola coriacea Nutt. on Hawaii [28], and Salvia pleberia R.Br. on Iriomote Island, Japan [29] (Figure 2).
B. pilosa infestation suppressed the growth of sugarcane (Saccharum officinarum L.) by 40% on day 60 and 80% on day 120 after sugarcane planting under field conditions in Okinawa, Japan, and caused 80% of the final production losses [30]. The species infestation reduced the production of a bean (Phaseolus vulgaris L.) by 48% in Uganda and by 18–48% in Peru [1]. The growth of this bean showed a significant negative correlation with the density of B. pilosa [31,32]. The species, at a density of 1.85 plants per m2, caused the bean yield to reduce by 18%, while 10 plants per m2 caused a reduction by 48% [33]. When B. pilosa and soybean (Glycine max (L.) Merr.) were grown together under field conditions, B. pilosa showed a higher relative growth rate than soybean and suppressed soybean’s vegetative growth and seed production [34]. A density of one and four plants of B. pilosa per m2, respectively, caused a soybean yield loss of 9.4% and 28% [2]. The contamination of the cypselae of B. pilosa also spoiled the quality of the crop grains [2]. When B. pilosa was germinated and grown with 16 crop plant species under different fertilizer and disturbance gradients under field conditions, B. pilosa showed a high competitive ability against crop plants from the families of Poaceae, Begoniaceae, Solanaceae, Balsaminaceae, Caryophyllaceae, and Convolvulaceae [27]. The stiff awns of its cypselae also irritate people and livestock [1,2]. Observations suggest that the infestation of B. pilosa causes a reduction in crop production and quality.
B. pilosa is also involved in the transmission of pathogens to agricultural crop plants as a vector. Sonchus yellow net virus was transmitted from B. pilosa to Nicotiana glutinosa L., Nicotiana clevelandii A.Gray, Zinnia elegans Jacq., and lettuce (Lactuca sativa L.) through the medium of the aphid (Hyperomyzus lactuae L.) [35,36]. B. pilosa has the potential to serve as a vector for tomato spotted wilt virus, which causes the typical tospovirus symptoms of chlorotic ringspots and necrosis on both young and older leaves of tomato (Lycopersicon esculentum Mill.) [37]. However, the movement of the virus between B. pilosa and tomato remains unknown. The papaya mealybug Paracoccus marginatus Williams & Granara de Willink, which is native to tropical America, was detected in South Asia, and B. pilosa was a host plant of the mealybug and transmitted it to other plant species [38]. The involvement of B. pilosa in the transmission of pathogens and insects may reduce the agricultural crop production.
As described in this section, the impacts of B. pilosa infestation on agricultural crop production were documented in several publications. However, its impacts on native plants and indigenous ecosystem are limited.
3. Fast Growth
B. pilosa established dense stands 50–135 days after its germination. The species starts flowering four–eight weeks after germination and produces mature seeds two–four weeks after flowering [39,40]. Regarding the population of the species in its favorite conditions, B. pilosa flowers throughout the year and makes up to four generations annually [1,2,5]. B. pilosa showed a higher growth rate, leaf area, and biomass than two indigenous congeners, Bidens bripartit (Lour.) Merr. & Sherff and Bidens tripartita L., under field conditions in China [41]. B. pilosa also showed more efficient utilization of water, photosynthetic fixed carbon, and nitrogen than the indigenous Cirsium setosum (Willd.) M. Bieb [42]. B. pilosa was reported to regenerate from remaining stems after cutting the above-ground parts of the plants [29]. These observations suggest that B. pilosa rapidly grows, establishes dense stands soon after germination, and produces several generations in a year.
4. Reproduction
The species produces about 80 capitula on long peduncles arising from the apexes of the stems and the leaf axils. A single capitulum contains ray and disk florets, as explained in the Introduction. The ray florets are sterile but serve as a nectar guide for pollinators. A nectary gland is present at the base of the style of disk florets [5]. The fertilized disk florets produce single-seeded black cypselae within four weeks [1,2,3,4]. Each plant produces 2000–6000 cypselae during its life cycles [1,2,43]. The species produces two types of cypselae within the same capitulum: long cypselae (8–10 mm in length) and short cypselae (3–7 mm). The production rate of the long and short cypselae was 64% and 36%, respectively [5,44]. Relatively young plants produced more long cypselae, and senescent plants produced more short cypselae [44,45]. Both types of cypselae have 2–5 stiff awns 2–4 mm long and can be easily dispersed by the attachment of the awns to animals, birds, and human clothes or by wind and water [5,43]. The seeds in cypselae remain viable for 5–6 years [2] (Figure 3).
Long cypselae have thin seed coats compared with short cypselae. Long cypselae germinated soon after their dispersion under a wide range of conditions, while short cypselae showed dormancy and germinated only under favorable conditions, such as adequate moisture and temperature, for the subsequent growth of seedlings [5,45,46]. The germination of short cypselae was stimulated by red light and gibberellin application [46], which indicates phytochrome involvement in the germination. Long cypselae showed no light sensitivity and germinated in both light and dark conditions [45]. Long cypselae produced three–four generations in a year due to their quick germination after dispersion, while short cypselae germinated in subsequent years due to their dormancy [5]. Long cypselae contribute to the species by continuously increasing the population, while short cypselae contribute to the species by remaining in reserve for the population [5,47].
The B. pilosa population also shows two phenological types of flowering–fruiting events during the growing season. Less than 10% of the population of the species flower one–two months after germination (early type), and the others flower four months after germination (normal type). Normal-type plants grow larger than early-type plants, and early-type plants produce fewer and heavier seeds than normal-type plants. The seeds from early-type plants more quickly germinate than those of normal-type plants, and both seeds do not show dormancy. The different flowering phenology affects seed mass and production and germination. However, the seeds from both types of plants produce early-type and normal-type offspring. Therefore, the flowering–fruiting events of their offspring are not affected by the parental types [48]. Early-type plants may contribute to the species quickly expanding because of their quick growth and flowering–fruiting. Normal-type plants may contribute to the species by stably increasing the population and/or maintaining the population.
The germination behavior of plants affects plant fitness and persistence. Invasive plant species often show earlier and/or rapid germination and asynchronous germination. The production of different biotypes of seeds by a single plant is also one of the characteristics of invasive plant species [49,50,51,52]. Early-germinating species and rapidly germinating species may benefit from less competition with other plant species for resources and niches [53,54]. These species enable the suppression of the germination and establishment of later-germinating species [51,52,55]. However, early and rapid germination is a risky strategy under unpredictable environmental conditions, especially in the early growing season, and often increases seedling mortality in such conditions [51,56,57]. Asynchronous germination may be more beneficial to the rapid expansion of species under relatively stable conditions and to the persistence and survival of species under unpredictable environmental conditions, regarded as a significant characteristic of invasive species to expand their ecological niches and population [50,58].
B. pilosa produced a great number of cypselae during its life cycle, different biotypes of cypselae such as long cypselae and short cypselae [5,43,45,46], and different phenological types of flowering–fruiting events such as the early-type flower and normal-type flower [48]. The asynchronous germination of B. pilosa is a strategy in which one of the seeds’ subsets may successfully germinate and establish itself at different times and under different conditions, providing the chance to survive and colonize in new habitats. Therefore, B. pilosa may be able to survive as a widespread invasive species in different habitats and to expand its distribution in introduced ranges.
In addition, B. pilosa showed different breeding systems at a variety of levels within the species. B. pilosa var. minor and B. pilosa var. pilosa produced 89% and 74% of seeds, respectively, compared to the corresponding open-pollinated capitula, although 73% of the seeds were set in open-pollinated capitula [25,37]. This observation suggested that B. pilosa var. radiata is highly self-incompatible, whereas the other two are self-compatible. Therefore, B. pliosa showed different breeding systems at a variety of levels within the species. The invasiveness of B. pilosa var. radiata was observed to be higher than that of B. pilosa var. minor and var. pilosa. When solar irradiation is high, B. pilosa var. radiata allocates more biomass to axillary shoots than the other two varieties, contributing to the expansion of the varieties [26]. Xenogamy enables the species to increase its genetic diversity, which favors its establishment in heterogenous and variable environments [5].
5. Adaptation
B. pilosa grows best in areas with full sunlight, a mean annual temperature between 25 °C and 38 °C, and annual rainfall between 500 mm and 3500 mm. It can grow in a wide range of soil types including sand and lime soil with a pH ranging from 4 to 9 and salinity up to 100 mM NaCl. The species tolerates frost, and the regeneration of its roots occurs after temperatures as low as −15 °C [1,2]. The species benefits from disturbances such as fire and soil tillage and quickly infests after a disturbance [19,59].
Under resource-limiting conditions, some invasive plant species can outcompete indigenous plant species either by efficient resource uptaking or by a lower resource requirement, called the resource conservative strategy [60]. The carbon (C), nitrogen (N), and phosphorus (P) contents in plant tissues and their ratios such as N:P and C:P reflect the use and adjustment of the available nutrients and the relative growth rate of the plants [61,62,63]. Lower N:P and C:P ratios in plant tissues indicate a higher P content in the whole plants and the stimulation of protein synthesis, resulting in higher growth and reproductive output [61]. A higher C:N ratio indicates higher C assimilation and lower nutrient requirements [63]. B. pilosa under low-nutrient conditions showed a higher C:N ratio in the roots, indicating a lower nutrient requirement and higher C assimilation, adapting a resource conservative strategy. B. pilosa under high-nutrient conditions showed lower N:P and C:P ratios in the roots, indicating higher protein synthesis, growth, and reproductive output [64]. When maize (Zea mays, L.) and B. pilosa grew together under field conditions, the C:N, C:P, C:K, N:P, and N:P ratios in B. pilosa significantly increased, while those ratios in maize changed a little, which indicates that B. pilosa has a distinct survival strategy under nutrient-competitive conditions, and is able to more efficiently use nutrients [65]. Therefore, B. pilosa may apply a competitive strategy in nutrient-rich environments and a resource conservative strategy in resource-poor environments.
B. pilosa showed a similar plant height as the indigenous plant species B. biternata (Lour.) Merr. & Sherff and B. tripartita L. under unfavorable light and moisture conditions. However, B. pilosa showed a greater plant height, biomass, and growth rate than both indigenous species under favorable light and moisture conditions. B. pilosa allocated more resources to the root biomass under a high-light condition and to the leaf biomass under a low-light condition [41]. When B. pilosa and Bidens biternata were grown in the conditions of different light intensities (40% and 10% sunlight) for 64 days, B. pilosa showed a higher leaf mass, a higher total leaf area, and increased photosynthesis than B. biternata [66]. These observations suggest that B. pilosa has a higher adaptative ability and a higher phenotypic plasticity than B. biternata and B. tripartita under low-light and low-moisture conditions, which may contribute to the invasiveness of B. pilosa.
Extreme precipitation often causes short-term waterlogging conditions for terrestrial plants, which cause hypoxia and serious damage to the plants’ root systems [67,68]. Short-term waterlogging stimulated adventitious root generation and higher dehydrogenase activity in the roots of B. pilosa [69,70], which is the adaptive response to the hypoxic condition [67,68]. The reduction in the photosynthesis and growth rate of B. pilosa under waterlogging conditions was significantly less than that of an indigenous species, B. biternata [70]. Therefore, B. pilosa may tolerate waterlogging conditions better than the indigenous species B. biternata.
The seeds of B. pilosa were collected from three different locations in China. Those seeds germinated at a temperature between 10 °C and 30 °C. The seeds lost viability after 8 days of continuous heating at 40 °C or 30 min heating at 50 °C. However, the intraspecific variation for high-temperature tolerance was found among seeds from different collection sites [71]. The germination requirements for temperature and water also significantly differed among seeds from different collection sites [72]. Seeds obtained from nine locations in Brazil showed 20–97% germination rates at a temperature of 10 °C–35 °C. The seed weight and the rates of germination and dormant seeds significantly varied among collection locations [73]. The germination rate of B. pilosa was higher with seeds buried in shallow soil than deep soil, and the seed dormancy was greater with deeper soil than shallow soil [74,75]. Therefore, the population of B. pilosa from different locations showed different tolerance and requirements for germination such as temperature and water and the different ratios of dormant seeds. The variation in the water requirement, temperature tolerance, and dormancy among seeds from different locations may be involved in the adaptation of B. pilosa to local conditions.
These observations suggest that the acclimation ability of B. pilosa to various environmental conditions such as soil fertility, temperature, and solar radiation is high. It also adapts to waterlogging conditions.
6. Genetic Variation
The species was divided into three varieties such as B. pilosa var. minor, var. pilosa, and var. radiata, and these varieties showed different morphological traits and adaptive potentials [25,26]. The genetic variation of five loci such as the chloroplast and nuclear ribosomal DNA of B. pilosa var. minor, var. pilosa, and var. radiata collected in Taiwan was found to be high, and each of these varieties showed different morphological traits and adaptive potentials [76]. However, those genetic variations have not been related to the invasiveness of the species. The genetic variation of plastomes in the Chinese population was very high, to the degree that specimens from the same population were misidentified as different species [77]. The genetic variations of B. pilosa complex were also determined for the identification of the species [78]. However, information on the genetic variation of the species is very limited. In addition, there is no information available for the genetic variation and invasiveness of the species.
7. Effects on Microbial Community
Invasive plants often change the soil condition and alter the soil microbial community, affecting the carbon and nitrogen cyclings, soil organic matter, and decomposition rate of the plant residues [79,80,81,82,83,84,85,86]. The rhizosphere of B. pilosa develops different soil microbial assemblages compared with indigenous plant species [87]. B. pilosa and the indigenous plant species Saussurea deltoidea (D.C.) C.B.Clark were grown in non-sterile and sterile soil obtained from under dominant stands of a native shrub, Dodonaea viscosa (L.) Jacq. B. pilosa showed a greater biomass in non-sterile soil than S. deltoidei, and soil sterilization had a more negative effect on the growth of B. pilosa than S. deltoidei [88]. These observations suggest that the soil microbiota under D. viscosa affects B. pilosa more than S. deltoidei, and B. pilosa may benefit in introduced ranges by using the native soil microbiota associated with D. viscosa. The nitrogen and potassium contents in the soil under B. pilosa were significantly higher than those under Bidens bipinnata L. under the condition of full sunlight [89], indicating that B. pilosa is involved in rapid nutrient mobilization in the full-sunlight condition.
Arbuscular mycorrhizal fungi (AMF) promote their host plant’s fitness by increasing water and nutrient acquisition and the defense performance against pathogen attacks and stress conditions [90,91,92,93,94]. The inoculation of AMF, Septoglomus vicosum, S. constrictum, and Glomus perpusillum, to B. pilosa increased its competitive ability, and AMF-inoculated B. pilosa outcompeted an indigenous plant species, Setaria viridis (L.) P.Beauv. The plant community in areas has finally been replaced by B. pilosa [95]. The inoculation of AMF, Funneliformis mosseas, Diversispora versiformis, and Glomus diaphnum and/or G. etunicatum, increased the biomass and the absorption ability of nitrogen and phosphorus of B. pilosa in a karst environment [96,97]. The inoculation of Glomus mosseae also increased the drought stress tolerance of B. pilosa [98]. Therefore, the mutualism of B. pilosa with AMF may increase its competitive ability and nutrient acquisition, resulting in a greater biomass of B. pilosa and the elimination of native plant species.
These observations suggest that B. pilosa may alter the soil microbial community and/or utilize the native soil microbiota. These microbes and their mutualism with AMF may contribute to increase the ability of B. pilosa regarding competition, nutrient acquisition, and stress tolerance.
8. Allelopathy
The interaction of introduced plants with indigenous plant species is one of the important determinants of the success of their naturalization in introduced ranges [99,100,101,102]. Invasive plants were often reported to have an ability of allelopathy. Allelopathy is the chemical interaction between donor plants and receiver plants, and chemicals involved in allelopathy are defined as allelochemicals [100,103]. Allelochemicals are released from donor plants into neighboring environments through volatilization, rainfall leachates, root exudation, and the decomposition of donor plant residues in the rhizosphere soil. Allelochemicals disturb the germination, growth, and regeneration process of neighboring plant species, and many invasive plants showed allelopathic activity [104,105,106]. Allelopathic plants synthesize and store allelochemicals in certain plant tissues until releasing them into neighboring environments [107,108,109,110,111]. Therefore, several researchers investigated the allelopathic activity and allelochemicals in extracts from different plant tissues and the residues of B. pilosa and its rhizosphere soil.
The chopped leaves of B. pilosa were incorporated into a rice paddy field (2 tons per hectare) in Thailand. The treatments suppressed the total emergence number and weight of 12 weed species by 15% and 18%, respectively, compared with the control, 30 days after the treatments. The major weeds in the area were Brachiaria mutica (Fordk.) Stapf., Commelina diffusa Rurm., Monochoria vaginalis Presl, and Marsilea quadrifolia L. B. pilosa suppressed the emergence total weed number by 4, 0, 35, and 31% of the control, and the total weed weight by 17, 0, 88, and 3% of the control for Brachiaria mutica, Commelina diffusa, Monochoria vaginalis, and Marsilea quadrifolia, respectively [112]. The mulching and incorporation of whole plants of B. pilosa into soil suppressed the weed emergence and density under greenhouse conditions [113]. The residues of the shoots and roots of B. pilosa incorporated into the soil suppressed the growth of Cyperus rotundus L. Even the soil collected under the B. pilosa-infested field also suppressed the growth of C. rotundus [114]. These investigations suggested that certain allelochemicals may be released into the soil during the decomposition process of the leaves, shoots, and roots of B. pilosa and suppress the emergence and growth of these weed species.
The continuous application of the root exudate of B. pilosa by a recirculation system suppressed the seedling growth of lettuce, bean, maize, and Sorghum bicolor (L.) Moench. The dry mass of lettuce, bean, maize, and Sorghum bicolor was reduced by 38%, 55%, 47%, and 58% on day 14, respectively [115]. The root exudate of B. pilosa also inhibited the germination and growth of Leucaena leucocephala (Lam.) de Wit., Echinochloa crus-galli(L.) P. Beauv., Medicago sativa L., and rice (Oryza sativa L.) [116]. When B. pilosa and Cyperus rotundus were grown together under pot conditions, the growth of Cyperus rotundus was suppressed. However, when activated carbon was added into the cultivation soil in the pots, the suppression was significantly reduced [114]. These investigations suggested that certain allelochemicals may be released into the soil as the root exudates of B. pilosa, and suppress the germination and growth of these plant species.
The spore germination, growth, and photosynthesis of the fern species Pteris multifida Poir. were also suppressed by the root exudation of B. pilosa. The root exudation increased the antioxidant enzyme activities in the gametophytes of the ferns such as superoxide dismutase, catalase, glutathione reductase, and glutathione S-transferase, which resulted in the death of the gametophytes. Palmitic acid (hexadecanoic acid; fatty acid) is a major compound in root exudates [117,118]. However, it is not certain if palmitic acid is involved in the inhibitory effect of the root exudate of B. pilosa because the allelopathic activity of this compound has not been proved.
Aqueous extracts of the whole parts of B. pilosa inhibited the germination of Amaranthus dubius Mart. ex. Thell [119] and the root growth of Ageratum conyzoides L. [120]. Fresh-cut B. pilosa (1 cm in length) or its aqueous extract was applied with irrigation water to Echinochloa crus-galli seedlings grown in pots (20 cm d.i.; 5 kg of soil). Both treatments resulted in increasing the mortality of E. crus-galli. When fresh cuts of B. pilosa were also applied to the rice paddy field with irrigation water, the treatment suppressed the weed emergence of the common weeds in rice fields such as E. crus-galli, E. coloa (L.) Link., Leptochloa chinensis (L.) Nees, Ludwigia hyssopifolia (G.Don) Exell, Sphenoclea zaylania Gaerth, Cypens iria L., Fimbristylis dichotoma (L.) Vahl., and Fimbristylis milacea (L.) Vahl. The treatments significantly increased rice production due to this inhibition of weed emergence [113,121]. These observations suggested that aqueous extracts of B. pilosa may contain certain allelochemicals and that these allelochemicals suppress the germination and growth of these plant species. Some allelochemicals may also be released by rainfall leachates because of the allelopathic activity of aqueous extracts and the irrigation water with fresh cuts of B. pilosa.
Boiling water extracts of the leaves, stems, and roots of B. pilosa inhibited the germination and growth of Raphanus sativus L. and Echinocholoa crus-galli. Phenol derivatives such as p-coumaric acid, caffeic acid, ferulic acid, p-hydroxybenzoic acid, vanillic acid, and salycilic acid were identified in the extracts [122]. These compounds were found in several other plant species and showed germination and growth inhibitory activity as allelopathic agents [123,124,125]. The inhibitory activity of those compounds was considered to be due to the disturbance of nutrient uptake, water transport, and photosynthesis [126,127,128]. Salycilic acid is also involved in the plant defense function against pathogen attacks [129].
Quercetin (flavonoid) was identified in the leaf extracts of B. pilosa [130]. Quercetin was reported to inhibit the growth and mitochondrial function of several plant species as an allelopathic agent [131,132,133,134]. Several terpenes were identified in the essential oil of B. pilosa [135]. Terpenoids were reported to be involved in the defense function of plants such as anti-fungal, anti-bacterial, and anti-feeding activities, in the interaction with insects such as for pollination, and the attraction of predators of their natural enemies [136,137,138,139,140,141]. Among the identified terpenoids found in B. pilosa, α-pinene and limonene, which are readily volatile, showed allelopathic activity and inhibited the germination and growth of several plant species [123,142].
These investigations suggested that whole parts of B. pilosa may contain water-extractable allelochemicals, and some of them may be released into its rhizosphere soil through the volatilization, rainfall leachate, root exudation, and decomposition processes. The novel weapons hypothesis indicates that the competitive ability of invasive plants is high due to allelochemicals (weapons) [99,100,103]. Therefore, these allelochemicals may be effective on indigenous plant species to suppress their regeneration process through germination and growth inhibition and contribute to the invasion of B. pilosa in introduced ranges (Figure 4).
9. Nematocidal, Fungicidal, and Insecticidal Activity
The interaction between introduced plants and natural enemies such as herbivore insects and pathogens is also one of the important determinants of the success of naturalization in introduced ranges [143,144,145]. When aqueous extracts of the leaves and roots of B. pilosa were applied into soil that contained the earthworm Eisenia fetida Savigny, the treatments resulted in the suppression of the body mass and respiration of the earthworm and in the enhancement of the oxidative- and DNA-damage biomarkers in the earthworm, in a concentration-dependent manner [146]. The aqueous extracts of the above-ground parts of B. pilosa increased the mortality of the root-knot nematode Meloidogyne incognita Kofoid & White, the pine-wood nematode (Bursaphelenchus xylophilus (Steiner & Buhrer) Nickle), and the pine sawyer (Monochamus alternatus Hope), which is the vector of the pine-wood nematode, with the extract concentration-dependent manner [147,148]. The boiling water extracts of the leaves, stems, and roots of B. pilosa showed the growth-inhibitory activity on the pathogenic fungi Corticium rolfsii (Curzi) C.C. Tu & Kimbr., Fusarium solani (Mart.) Sacc., and F. oxysporum Schlecht. emend.Snyder & Hansen [122].
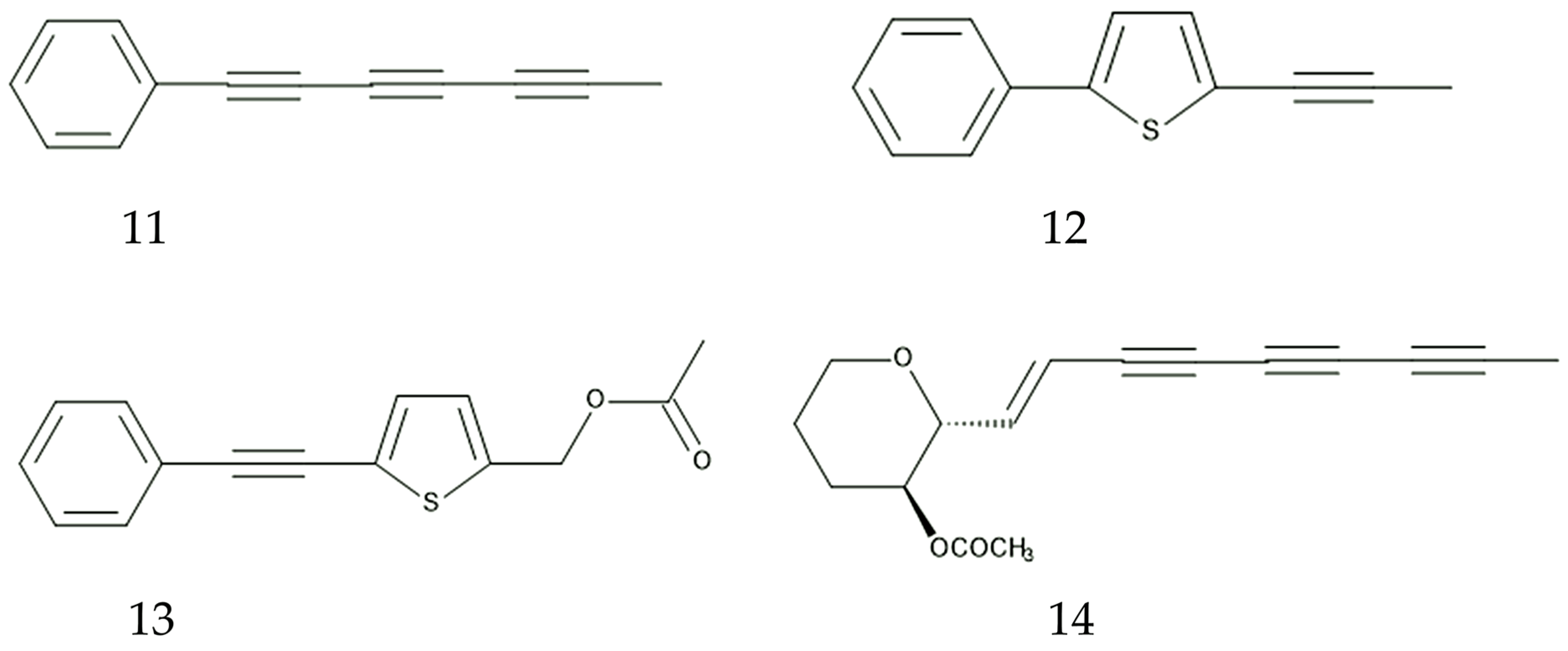
The methanol leaf extracts of B. pilosa showed insecticidal activity against three pests, the rice weevil (Sitophilus oryzae L.), grain beetle (Oryzaephilus surinamensis L.), and bean weevil (Acanthoscelides obtectus Say), with the extract concentration-dependent manner [149]. The spray of the aqueous extracts of B. pilosa reduced pest species such as flower beetles (Epicauta albovittata Gestro and E. limbatipennis Pic), foliage beetles (Ootheca mutabilis Sahlberg, and O. bennigseni Weise), and pod suckers (Clavigralla tomentosicollis Stäl and C. schadabi Dollong) [150]. Four polyacetylenes, 1-phenylhepta-1,3,5-trine, 5-phenyl-2-(1-propynyl)-thiophene, 5-actoxy-2-phenylethinyl-thiophene, and icthyothereol acetate isolated from the above-ground parts of B. pilosa showed insecticidal activity against the larvae of six moth species, Plutella xylostella L., Spodoptera litura Fabricius, Mythimna separata Walker, Spodoptera exigua Hübner, Helicoverpa armigera Hübner, and Ostrinia furnacalis Guenée, with LD50 values ranging between 0.133 and 4.084 μg per larva. However, 1-phenylhepta-1,3,5-trine, among the four polyacetylenes, showed the highest insecticidal activity [151]. 1-phenylhepta-1,3,5-trine (phenylheptatriyne) also showed nematocidal activity [152] and allelopathic activity against the germination and growth of Asclepias syriaca L., Chenopodium album L., Phlem pratense L., and Trifolium pratense L. [153].
These investigations suggested that B. pilosa contains water and/or methanol extractable compounds that have nematocidal, fungicidal, and insecticidal activity. These compounds may contribute to the increase in the fitness of B. pilosa and be involved in its invasiveness in introduced ranges (Figure 5).
10. Secondary Metabolites
Recent pharmacological investigations showed that B. pilosa contains more than 200 natural compounds, in many chemical classes such as fatty acids, polyacetylenes, flavonoids, terpenoids, phenolics, and their glycosides. Some of these compounds showed pharmacological activity such as anti-malaria, anti-cancer, anti-diabetic, anti-allergic, anti-hypertensive, and anti-inflammatory activity [11,14,154,155,156,157,158]. Although most of the identified compounds in B. pilosa have not yet been related to its invasiveness, some of these identified compounds may be involved in allelopathy and defense functions against herbivores and pathogens. As described in Section 7 and Section 8, the extracts, root exudates, residues, and rhizosphere soil of B. pilosa showed allelopathic activity and nematocidal, fungicidal, and insecticidal activity. However, the entirety of the compounds involved in these activities may not be identified. Therefore, some of these compounds and/or unidentified compounds in B. pilosa may be involved in allelopathy and defense functions against herbivores and pathogens. In fact, several secondary metabolites in invasive plants showed multiple functions such as anti-herbivore, anti-fungal, and allelopathic activity [100,102,159,160,161]. Some of these compounds in B. pilosa may contribute to the invasiveness and naturalization of species in introduced ranges.
11. Prospective
The impact of biological invasions with global warming has been identified as a serious threat to biodiversity on the planet [162,163,164]. The increasing mean annual temperature promotes the invasion of alien species originating from warmer regions [165,166]. Warmer conditions enhance the competitive ability of alien plant species [167]. The growth and reproduction of B. pilosa was enhanced in warmer conditions by the shifting of its phenology to early germination and flowering and prolonged vegetative growth and reproduction [168]. Global warming trends may favor the spread of B. pilosa northward in the Northern Hemisphere and southward in the Southern Hemisphere, originating from warmer regions. The species will possibly expand into these areas in the near future and may cause serious threats to the natural ecosystems in the areas.
Several natural enemies of B. pilosa have been recorded in its native ranges such as Cercospora bidentis Tharp, Ralstonia solanacearum Smith, Sclerotinia sclerotiorum (Lib.) de Baary, Bidens mosaic virus, and Sonchus yellow net virus [2,169,170,171]. However, biological control agents for B. pilosa have not yet been sufficiently investigated [2]. The management of B. pilosa can be achieved with several herbicides such as glyphosate, atrazine, 2,4-D glyphosate, imazethapyr, metribuzin, and paraquat [2,75,172,173]. However, several herbicide-resistant biotypes of B. pilosa against glyphosate, atrazine, and imazethapyr were reported [174,175]. The appearance of multiple herbicide-resistant biotypes is a serious issue for weed management. The most effective management strategy for herbicide-resistant weeds is probably to rotate the herbicide’s sites of actions [176,177]. The employment of herbicide mixtures and the rotation of different types of herbicides in different growing seasons may be one of the options to control the herbicide-resistant biotypes of B. pilosa [178,179].
12. Conclusions
An annual or biennial plant species, B. pilosa quickly grows and has several generations during a growing season. Each plant produces 2000–6000 cypselae during its life cycles. There are different biotypes of cypselae: quick germination after dispersion, late germination with dormancy, early flowering–fruiting types, and normal flowering–fruiting types. Asynchronous germination and flowering–fruiting may be beneficial for the persistence and survival of the species under unpredictable environmental conditions and for rapidly increasing the population under relatively stable conditions. B. pilosa showed a highly adaptative ability regarding water, light, and nutrient availability, temperature, and flooding. The populations of B. pilosa from different locations showed different requirements for germination such as temperature and water and different ratios of dormant seeds. B. pilosa altered the microbial community in infested areas, and mutualism with arbuscular mycorrhizal fungi increased the competitive ability of B. pilosa. The species showed allelopathic activity and contains several allelochemicals, which suppress the germination and growth of other plant species. B. pilosa also showed defense functions against pathogen fungi, nematodes, and insects and contains compounds involved in the functions. These characteristics of B. pilosa may contribute to the naturalization and invasiveness of the species. Global warming trends may also favor the spread of this species into additional non-native areas and may increase the threat of the species.
Funding
This research received no external funding.
Acknowledgments
The authors acknowledge the review journal funding: contract No. 1549/UN6.3.1/PT.00/2023, and the publication charge from Universitas Padjadjaran, Bandung, Indonesia.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Global Invasive Species Database, Bidens pilosa. Available online: http://www.iucngisd.org/gisd/speciesname/Bidens+pilosa (accessed on 4 August 2023).
- Invasive Species Compendium, Bidens pilosa. Available online: https://www.cabidigitallibrary.org/doi/full/10.1079/cabicompendium.9148 (accessed on 4 August 2023).
- Plants of the World. Royal Botanical Gardens-Kew, Bidens pilosa. Available online: https://powo.science.kew.org/taxon/urn:lsid:ipni.org:names:32564-2 (accessed on 4 August 2023).
- Huang, Y.L.; Chen, S.-J.; Kao, W.-Y. Floral biology of Bidens pilosa var. radiata, an invasive plant in Taiwan. Bot. Stud. 2012, 53, 501–507. [Google Scholar]
- Budumajji, U.; Raju, A.J.S. Pollination ecology of Bidens pilosa L. (Asteraceae). Taiwania 2018, 63, 89–100. [Google Scholar]
- Duke, J.A. Duke’s Handbook of Medicinal Plants of Latin America; CRC Press: Boca Raton, FL, USA, 2008; pp. 1–960. [Google Scholar]
- Mtenga, D.V.; Ripanda, A.S. A review on the potential of underutilized Blackjack (Biden Pilosa) naturally occurring in sub-Saharan Africa. Heliyon 2022, 8, e09586. [Google Scholar] [CrossRef]
- Yang, R.Y.; Keding, G.B. Nutritional contributions of important African indigenous vegetables. In African Indigenous Vegetables in Urban Agriculture; Shackleton, C.M., Pasquini, M.W., Drescher, A.W., Eds.; Earthcan Publishing: London, UK, 2009; pp. 105–136. [Google Scholar]
- Arthur, G.D.; Naidoo, K.K.; Coopoosamy, R.M. Bidens pilosa L.: Agricultural and pharmaceutical importance. J. Med. Plants Res. 2012, 6, 3282–3287. [Google Scholar] [CrossRef]
- Otang-Mbeng, W.; Mashabela, M.N. A review of beneficial phytochemicals and postharvest studies on some indigenous leafy vegetables from the Mpumalanga Province of South Africa. Med. Plants Int. J. Phytomed. Relat. Ind. 2020, 12, 533–544. [Google Scholar] [CrossRef]
- Kuo, T.F.; Yang, G.; Chen, T.Y.; Wu, Y.C.; Minh, T.R.H.; Chen, L.S.; Chen, W.C.; Huang, M.G.; Liang, Y.C.; Yang, W.C. Bidens pilosa: Nutritional value and benefits for metabolic syndrome. Food Front. 2021, 2, 32–45. [Google Scholar] [CrossRef]
- Connelly, P. Horrible weed or miracle herb?: A review of Bidens pilosa. J. Aust. Tradit.-Med. Soc. 2009, 15, 77–79. [Google Scholar]
- Bartolome, A.P.; Villaseñor, I.M.; Yang, W.C. Bidens pilosa L.(Asteraceae): Botanical properties, traditional uses, phytochemistry, and pharmacology. Evid.-Based Complement. Altern. Med. 2013, 2013, 340215. [Google Scholar] [CrossRef]
- Xuan, T.D.; Khanh, T.D. Chemistry and pharmacology of Bidens pilosa: An overview. J. Pharm. Investig. 2016, 46, 91–132. [Google Scholar] [CrossRef]
- Mzengereza, K.; Msiska, O.V.; Kapute, F.; Kang’ombe, J.; Singini, W.; Kamangira, A. Nutritional value of locally available plants with potential for diets of tilapia Rendalli in pond aquaculture in Nkhata Bay, Malawi. J. Aquac. Res. Dev. 2014, 5, 265. [Google Scholar]
- Kato-Noguchi, H. Allelopathy of knotweeds as invasive plants. Plants 2022, 11, 3. [Google Scholar] [CrossRef]
- Kato-Noguchi, H. Allelopathy and allelochemicals of Imperata cylindrica as an invasive plant species. Plants 2022, 11, 2551. [Google Scholar] [CrossRef]
- Kato-Noguchi, H. Invasive mechanisms of one of the world’s worst alien plant species Mimosa pigra and its management. Plants 2023, 12, 1960. [Google Scholar] [CrossRef]
- Sankaran, K.V.; Suresh, T.A.; FAO Regional Office for Asia and the Pacific. No.2013/06 213, Bangkok, Thailand. Available online: http://www.fao.org/asiapacific/rap/publications/pub-rap-results/en/ (accessed on 4 August 2023).
- Davidse, G.; Sousa-Sánchez, M.; Knapp, S.; Chiang, F.; Ulloa, U.C.; Pruski, J.F. Flora Mesoamericana, Vol. 5, Part 2: Asteraceae; Missouri Botanical Garden Press: St. Louis, MO, USA, 2018; pp. 1–608. [Google Scholar]
- Queensland Government, Weeds of Australia, Biosecurity Queensland Edition. Australia: Queensland Government. Available online: http://keyserver.lucidcentral.org/weeds/data/media/Html/search.html (accessed on 4 August 2023).
- Pacific Islands Ecosystems at Risk. University of Hawaii. Available online: http://www.hear.org/pier/index.html (accessed on 4 August 2023).
- Reaser, J.K.; Meyerson, L.A.; Cronk, Q.; De Poorter, M.A.J.; Eldrege, L.G.; Green, E.; Kairo, M.; Latasi, P.; Mark, R.N.; Mauremootoo, J.; et al. Ecological and socioeconomic impacts of invasive alien species in island ecosystems. Environ. Conser. 2007, 34, 98–111. [Google Scholar] [CrossRef]
- Russell, J.C.; Meyer, J.Y.; Holmes, N.D.; Pagad, S. Invasive alien species on islands: Impacts, distribution, interactions and management. Environ. Conser. 2017, 44, 359–370. [Google Scholar] [CrossRef]
- Huang, Y.L.; Kao, W.Y. Different breeding systems of three varieties of Bidens pilosa in Taiwan. Weed Res. 2013, 54, 162–168. [Google Scholar] [CrossRef]
- Huang, Y.L.; Kao, W.Y. Comparisons of growth, biomass allocation, and morphology of an invasive and two non-invasive varieties of Bidens pilosa in Taiwan. Taiwania 2016, 61, 288–294. [Google Scholar]
- Ng, C.C.; Wu, S.J.; Wang, C.Y.; Tzeng, W.S.; Shyu, Y.T. Emergence and growth of beggarticks (Bidens pilosa var. radiata) in different plant communities under experimental field conditions. J. Agric. Sci. Technol. A 2011, 1, 950–962. [Google Scholar]
- US Fish and Wildlife Service. Panicum fauriei var. carteri. 5-Year Review: Summary and Evaluation; US Fish and Wildlife Service: Washington, DC, USA, 2011; pp. 1–17. [Google Scholar]
- Shimamoto, Y.; Nomura, N.; Takaso, T.; Setoguchi, H. Overcompensation of seed production caused by clipping of Bidens pilosa var. radiata (Compositae): Implications for weed control on Iriomote-Jima Island, Japan. Weed Biol. Manag. 2011, 11, 118–126. [Google Scholar] [CrossRef]
- Ishimine, Y.; Miyazato, K.; Matsumoto, S. Physiological and ecological characteristics of weeds of sugarcane fields in the Ryukyu Islands. 7 Competition between sugarcane and Bidens pilosa L. var. radiata Scherff. at earlier stages of growth. Weed Res. Japan 1986, 31, 287–293. [Google Scholar] [CrossRef]
- Blanco, H.G.; Arévalo, R.A.; Blanco, F.M.G. Injury of Bidens pilosa L. on bean plants. Arq. Do Inst. Biológico 1996, 63, 35–40. [Google Scholar]
- de Carvalho, D.A. Study of specific competition of weeds on bean crop. 1. Competitive effects of Alexander grass (Brachiaria plantaginea (Link) Hitch) and hairy beggarticks (Bidens pilosa L.) in different densities on growth and mineral nutrition of common bean (Phaseolus vulgaris L.). Cienc. E Prat. 1980, 24, 1131–1137. [Google Scholar]
- Cerna, L.; Valdez, V. The influence of populations of the weeds Sorghum halepense (L.) Pers. and Bidens pilosa L. on the yield of beans (Phaseolus vulgaris L.). Pirata 2. Turrialba 1987, 37, 303–309. [Google Scholar]
- Chivinge, O.A.; Schwappenhauser, M.A. Competition of soybean with blackjack (Bidens pilosa L.) and pigweed (Amaranthus hybridus L.). Afr. Crop Sci. J. 1995, 3, 73–82. [Google Scholar] [CrossRef]
- Christie, S.R.; Christie, R.G.; Edwardson, J.R. Transmission of a bacilliform virus of sowthistle and Bidens pilosa. Phytopathology 1974, 64, 840–845. [Google Scholar] [CrossRef]
- Andrew, O.; Dietzgen, R.G.; Goodin, M.M.; Li, Z. Development of model systems for plant rhabdovirus research. Adv. Virus Res. 2018, 202, 23–57. [Google Scholar]
- Huang, C.J.; Liu, Y.; Yu, H.Q.; Liu, B.Z.; Qing, L. Bidens pilosa is a natural host of Tomato spotted wilt virus in Yunnan Province, China. Plant Dis. 2016, 100, 1957. [Google Scholar] [CrossRef]
- Saengyot, S.; Burikam, I. Host plants and natural enemies of papaya mealybug, Paracoccus marginatus (Hemiptera: Pseudococcidae) in Thailand. Thai J. Agric. Sci. 2011, 44, 197–205. [Google Scholar]
- Ishimine, Y.; Miyazato, K.; Matsumoto, S. Physiological and ecological characteristics of weeds of sugarcane fields in the Ryukyu Islands. J. Weed Sci. Technol. 1986, 31, 16–23. [Google Scholar] [CrossRef]
- Asami, K.; Yamato, M.; Hattori, T.; Akamatsu, H.; Takeda, Y. Floristic composition and process of establishment of the Bidens pilosa var. minor—Imperata cylindrica community maintained by cutting on non-arable land in Okinawa Prefecture. Veg. Sci. 1999, 16, 1–11. [Google Scholar]
- Pan, Y.; Tang, S.; Wei, C.; Li, X. Comparison of growth, photosynthesis and phenotypic plasticity between invasive and native Bidens species under different light and water conditions. Biodivers. Sci. 2017, 25, 1257–1266. [Google Scholar]
- Chen, X.W.; Li, H.Y.; Liu, H.M.; Yang, D.L.; Huangfu, C.H. Comparison of gas exchange characteristics between invasive Parthenium hysterophorus and Bidens pilosa and co-occurring native Cirsium setosum (Asteraceae). Acta Ecol. Sin. 2016, 36, 5732–5740. [Google Scholar]
- Rocha, O.J. The effects of achene heteromorphism on the dispersal capacity of Bidens pilosa L. Int. J. Plant Sci. 1996, 157, 316–322. [Google Scholar] [CrossRef]
- Hadač, E.; Hadačová, V. Notes on the ecology and distribution of Bidens pilosa L. in Cuba. Folia Geobot. Phytotax. 1998, 6, 165–173. [Google Scholar]
- Amaral, A.; Takaki, M. Achene dimorphism in Bidens pilosa L.(Asteraceae) as determined by germination test. Braz. Arch. Biol. Technol. 1998, 41, 10–15. [Google Scholar] [CrossRef]
- Forsyth, C.; Brown, N.A.C. Germination of the dimorphic fruits of Bidens pilosa L. New Phytol. 1982, 90, 151–164. [Google Scholar] [CrossRef]
- Imbert, E.; Ronce, O. Phenotypic plasticity for dispersal ability in the seed heteromorphic Crepissancta (Asteraceae). Oikos 2001, 93, 126–134. [Google Scholar] [CrossRef]
- Gurvich, D.E.; Enrico, L.; Funes, G.; Zak, M.R. Seed mass, seed production, germination and seedling traits in two phenological types of Bidens pilosa (Asteraceae). Aust. J. Bot 2004, 52, 647–652. [Google Scholar] [CrossRef]
- Harper, J. Population Biology of Plants; Academic Press: London UK, 1977; pp. 1–892. [Google Scholar]
- García, Y.; Callaway, R.M.; Diaconu, A.; Montesinos, D. Invasive and non-invasive congeners show similar trait shifts between their same native and non-native ranges. PLoS ONE 2013, 8, e82281. [Google Scholar] [CrossRef]
- Gioria, M.; Pyšek, P. The legacy of plant invasions: Changes in the soil seed bank of invaded plant communities. Bioscience 2016, 66, 40–53. [Google Scholar] [CrossRef]
- Gioria, M.; Pyšek, P. Early bird catches the worm: Germination as a critical step in plant invasion. Biol. Invasions 2017, 19, 1055–1080. [Google Scholar] [CrossRef]
- Abraham, J.K.; Corbin, J.D.; D’Antonio, C.M. California native and exotic perennial grasses differ in their response to soil nitrogen, exotic annual grass density, and order of emergence. Plant Ecol. 2009, 201, 445–456. [Google Scholar] [CrossRef]
- Gioria, M.; Osborne, B. Resource competition in plant invasions: Emerging patterns and research needs. Front. Plant Sci. 2014, 5, 501. [Google Scholar] [CrossRef] [PubMed]
- Fenesi, A.; Albert, Á.J.; Ruprecht, E. Fine-tuned ability to predict future competitive environment in Ambrosia artemisiifolia seeds. Weed Res. 2014, 54, 58–69. [Google Scholar] [CrossRef]
- Ruxton, G.D.; Schaefer, H.M. The conservation physiology of seed dispersal. Philos. Trans. R Soc. Lond B Biol. Sci. 2012, 367, 1708–1718. [Google Scholar] [CrossRef]
- Wainwright, C.E.; Wolkovich, E.M.; Cleland, E.E. Seasonal priority effects: Implications for invasion and restoration in a semi-arid system. J. Appl. Ecol. 2012, 49, 234–241. [Google Scholar] [CrossRef]
- Donohue, K.; de Casas, R.R.; Burghardt, L.; Kovach, K.; Willis, C.G. Germination, post-germination adaptation, and species ecological ranges. Annu. Rev. Ecol. Evol. Syst. 2010, 41, 293–319. [Google Scholar] [CrossRef]
- Galinato, M.I.; Moody, K.; Piggin, C.M. Upland Rice Weeds of South and Southeast Asia; International Rice Institute: Los Baños, Philippines, 1999; pp. 1–156. [Google Scholar]
- Funk, J.L.; Vitousek, P.M. Resource-use efficiency and plant invasion in low-resource systems. Nature 2007, 446, 1078–1081. [Google Scholar] [CrossRef]
- Elser, J.J. Biological stoichiometry: A chemical bridge between ecosystem ecology and evolutionary biology. Am. Nat. 2006, 168, 25–35. [Google Scholar] [CrossRef] [PubMed]
- Kerkhoff, A.J.; Fagan, W.F.; Elser, J.J.; Enquist, B.J. Phylogenetic and growth form variation in the scaling of nitrogen and phosphorus in the seed plants. Am. Nat. 2006, 168, 103–122. [Google Scholar] [CrossRef] [PubMed]
- González, A.L.; Kominoski, J.S.; Danger, M.; Ishida, S.; Iwai, N.; Rubach, A. Can ecological stoichiometry help explain patterns of biological invasions. Oikos 2010, 119, 779–790. [Google Scholar] [CrossRef]
- Wang, J.; Chen, W.; Zhu, H. Ecological stoichiometry and invasive strategies of two alien species (Bidens pilosa and Mikania micrantha) in subtropical China. Ecol. Res. 2019, 34, 612–623. [Google Scholar] [CrossRef]
- de Matos, C.C.; Teixeira, R.D.S.; da Silva, I.R.; Costa, M.D.; da Silva, A.A. Interspecific competition changes nutrient: Nutrient ratios of weeds and maize. J. Plant Nutr. Soil Sci. 2019, 182, 286–295. [Google Scholar] [CrossRef]
- Wang, R.L.; Yan, W.B.; Quan, G.M.; Liu, S.W.; Zhang, J.E. Effects of light intensity on morphology and physiology of exotic invasive Bidens pilosa L. and non-invasive congener Bidens bipinnata L. Allelopath. J. 2017, 42, 157–168. [Google Scholar] [CrossRef]
- Drew, M.C. Oxygen deficiency and root metabolism: Injury and acclimation under hypoxia and anoxia. Ann. Rev. Plant Biol. 1997, 48, 223–250. [Google Scholar] [CrossRef] [PubMed]
- Fukao, T.; Bailey-Serres, J. Plant responses to hypoxia–is survival a balancing act? Trends Plant Sci. 2004, 9, 449–456. [Google Scholar] [CrossRef]
- Hsu, H.M.; Kao, W.Y. Vegetative and reproductive growth of an invasive weed Bidens pilosa L. var. radiata and its noninvasive congener Bidens bipinnata in Taiwan. Taiwania 2014, 59, 119–126. [Google Scholar]
- Yue, M.; Shen, H.; Li, W.; Chen, J.; Ye, W.; Tian, X.; Yin, A.; Cheng, S. Waterlogging tolerance of Bidens pilosa translates to increased competitiveness compared to native Bidens biternata. Plant Soil 2019, 437, 301–311. [Google Scholar] [CrossRef]
- Wen, B.; An, X.; Chen, L.; Yin, X.; Mi, L. Variation in Tolerance to High Temperature and Water Stress in Weedy Bidens pilosa Seeds from Yunnan, SW China. Research Square. 2022. Available online: https://www.researchsquare.com/article/rs-1782585/v1 (accessed on 4 August 2023).
- Zhang, R.; Chen, D.; Liu, H.; Guo, C.; Tang, L.; Wang, H.; Chen, Y.; Luo, K. Effect of temperature and water potential on the germination of seeds from three different populations of Bidens pilosa as a potential Cd hyperaccumulator. BMC Plant Biol. 2022, 22, 487. [Google Scholar] [CrossRef] [PubMed]
- de Barros, R.T.; Martins, C.C.; da Silva, G.Z.; Martins, D. Origin and temperature on the germination of beggartick seeds. Rev. Bras. Eng. Agríc. Ambient. 2017, 21, 448–453. [Google Scholar] [CrossRef]
- Sahoo, U.K.; Jha, L.K. Effect of depth and duration of burial on seed viability and dormancy of Bidens pilosa L. and Richardsonia pilosa HBK. Seed Res. 1997, 25, 5–10. [Google Scholar]
- Chauhan, B.S.; Ali, H.H.; Florentine, S. Seed germination ecology of Bidens pilosa and its implications for weed management. Sci. Rep. 2019, 9, 16004. [Google Scholar] [CrossRef] [PubMed]
- Tsai, L.C.; Wang, J.C.; Hsieh, H.M.; Liu, K.L.; Linacre, A.; Lee, J.C.I. Bidens identification using the noncoding regions of chloroplast genome and nuclear ribosomal DNA. Forensic Sci. Int. Genet. 2008, 2, 35–40. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Jiang, M.; Liu, M.; Wang, B.; Yu, Q.; Chen, H.; Wang, L.; Liu, C. Analysis of the complete plastomes of Bidens pilosa L. 1753 (Asteraceae, Coreopsideae) from Beijing, China reveals high genetic diversity and possible misidentifications. Mitochondrial DNA B Resour. 2023, 8, 612–618. [Google Scholar] [CrossRef] [PubMed]
- Grombone-Guaratini, M.T.; Semir, J.; Solferini, V.N. Low allozymic variation in the Bidens pilosa L. complex (Asteraceae). Biochem. Genet. 2005, 43, 335–345. [Google Scholar] [CrossRef]
- van der Putten, W.H. Impacts of soil microbial communities on exotic plant invasions. Trends Ecol. Evol. 2010, 25, 512–519. [Google Scholar]
- Castro-Diez, P.; Godoy, O.; Alonso, A.; Gallardo, A.; Saldana, A. What explains variation in the impacts of exotic plant invasions on the nitrogen cycle? A meta-analysis. Ecol. Lett. 2014, 17, 1–12. [Google Scholar] [CrossRef]
- Callaway, R.M.; Maron, J.L. What have exotic plant invasions taught us over the past 20 years? Trends Ecol. Evol. 2006, 21, 369–374. [Google Scholar] [CrossRef]
- Sun, X.; Gao, C.; Guo, L. Changes in soil microbial community and enzyme activity along an exotic plant Eupatorium adenophorum invasion in a Chinese secondary forest. Chin. Sci. Bull. 2013, 58, 4101–4108. [Google Scholar] [CrossRef]
- Fenner, M. The inhibition of germination of Bidens pilosa seeds by leaf canopy shade in some natural vegetation types. New Phytol. 1980, 84, 95–101. [Google Scholar] [CrossRef]
- Elgersma, K.J.; Ehrenfeld, J.G. Linear and non-linear impacts of a non-native plant invasion on soil microbial community structure and function. Biol. Invasions 2010, 13, 757–768. [Google Scholar] [CrossRef]
- Li, W.; Zhang, C.; Jiang, H.; Xin, G.; Yang, Z. Changes in soil microbial community associated with invasion of the exotic weed, Mikania micrantha H.B.K. Plant Soil 2006, 281, 309–324. [Google Scholar] [CrossRef]
- Hawkes, C.V.; Wren, I.F.; Herman, D.J.; Firestone, M.K. Plant invasion alters nitrogen cycling by modifying the soil nitrifying the soil nitrifying community. Ecol. Lett. 2005, 8, 976–985. [Google Scholar] [CrossRef]
- Wang, L.; Li, Q.; Li, C.; Wu, C.; Chen, F.; Chen, X.; Zhang, F. Nitrate nitrogen and pH correlate with changes in rhizosphere microbial community assemblages during invasion of Ambrosia artemisiifolia and Bidens pilosa. Microbiol. Spectr. 2023, 11, e03649-22. [Google Scholar] [CrossRef] [PubMed]
- Cui, Q.G.; He, W.M. Soil biota, but not soil nutrients, facilitate the invasion of Bidens pilosa relative to a native species Saussurea deltoidea. Weed Res. 2009, 49, 201–206. [Google Scholar] [CrossRef]
- Wei, H.; Yan, W.; Quan, G.; Zhang, J.; Liang, K. Soil microbial carbon utilization, enzyme activities and nutrient availability responses to Bidens pilosa and a non-invasive congener under different irradiances. Sci. Rep. 2017, 7, 11309. [Google Scholar] [CrossRef]
- Smith, S.E.; Read, D.J. Mycorrhizal Symbiosis, 3rd ed.; Academic Press: London, UK, 2008; pp. 1–815. [Google Scholar]
- Diagne, N.; Ngom, M.; Djighaly, P.I.; Fall, D.; Hocher, V.; Svistoonoff, S. Roles of arbuscular mycorrhizal fungi on plant growth and performance: Importance in biotic and abiotic stressed regulation. Diversity 2020, 12, 370. [Google Scholar] [CrossRef]
- Tang, H.; Hassan, M.U.; Feng, L.; Nawaz, M.; Shah, A.N.; Qari, S.H.; Liu, Y.; Miao, J. The critical role of arbuscular mycorrhizal fungi to improve drought tolerance and nitrogen use efficiency in crops. Front. Plant Sci. 2022, 13, 919166. [Google Scholar] [CrossRef]
- Brundrett, M. Mycorrhizas in natural ecosystems. Adv. Ecol. Res. 1991, 21, 171–313. [Google Scholar]
- Vandenkoornhuyse, P.; Ridgway, K.P.; Watson, I.J.; Fitter, A.H. Co-existing grass species have distinctive arbuscular mycorrhizal communities. Mol. Ecol. 2003, 12, 3085–3095. [Google Scholar] [CrossRef] [PubMed]
- Zhang, F.J.; Li, Q.; Yerger, E.H.; Chen, X.; Shi, Q.; Wan, F.H. AM fungi facilitate the competitive growth of two invasive plant species, Ambrosia artemisiifolia and Bidens pilosa. Mycorrhiza 2018, 28, 703–715. [Google Scholar] [CrossRef]
- He, Y.; Jiang, C.; Yang, H.; Wang, Y.; Zhong, Z. Arbuscular mycorrhizal fungal composition affects the growth and nutrient acquisition of two plants from a karst area. Sains Malays. 2017, 46, 1701–1708. [Google Scholar] [CrossRef]
- Shen, K.; He, Y.; Xu, X.; Umer, M.; Liu, X.; Xia, T.; Guo, Y.; Wu, B.; Xu, H.; Zang, L.; et al. Effects of AMF on plant nutrition and growth depend on substrate gravel content and patchiness in the karst species Bidens pilosa L. Front. Plant Sci. 2022, 13, 968719. [Google Scholar] [CrossRef] [PubMed]
- Song, H.; Zhong, Z.; Yang, W.; Chen, Q. Analysis of the activities of protective enzymes in Bidens pilosa L. inoculated with Glomus mosseae under drought stress. Acta Ecol. Sinica 2011, 31, 2471–2477. [Google Scholar]
- Callaway, R.M.; Aschehoug, E.T. Invasive plants versus their new and old neighbors: A mechanism for exotic invasion. Science 2000, 290, 521–523. [Google Scholar] [CrossRef]
- Cappuccino, N.; Arnason, J.T. Novel chemistry of invasive exotic plants. Biol. Lett. 2006, 2, 189–193. [Google Scholar] [CrossRef]
- Wang, C.; Zhu, M.; Chen, X.; Qu, B. Review on allelopathy of exotic invasive plants. Procedia Engin. 2011, 18, 240–246. [Google Scholar]
- Meiners, S.J.; Kong, C.H.; Ladwig, L.M.; Pisula, N.L.; Lang, K.A. Developing an ecological context for allelopathy. Plant. Ecol. 2012, 213, 1861–1867. [Google Scholar] [CrossRef]
- Callaway, R.M.; Ridenour, W.M. Novel weapons: Invasive success and the evolution of increased competitive ability. Front Ecol. Environ. 2004, 2, 436–443. [Google Scholar] [CrossRef]
- Kato-Noguchi, H.; Kato, M. Evolution of the secondary metabolites in invasive plant species Chromolaena odorata for the defense and allelopathic functions. Plants 2023, 12, 521. [Google Scholar] [CrossRef]
- Kato-Noguchi, H.; Kurniadie, D. Allelopathy and allelochemicals of Leucaena leucocephala as an invasive plant species. Plants 2022, 11, 1672. [Google Scholar] [CrossRef]
- Kato-Noguchi, H.; Kato, M. Allelopathy and allelochemicals of Solidago canadensis L. and S. altissima L. for their naturalization. Plants 2022, 11, 3235. [Google Scholar] [CrossRef] [PubMed]
- Rice, E.L. Allelopathy, 2nd ed.; Academic Press: Orlando, FL, USA, 1984; pp. 1–422. [Google Scholar]
- Bais, H.P.; Weir, T.L.; Perry, L.G.; Gilroy, S.; Vivanco, J.M. The role of root exudates in rhizosphere interactions with plants and other organisms. Annu. Rev. Plant Biol. 2006, 57, 233–266. [Google Scholar] [CrossRef]
- Kato-Noguchi, H. Defensive molecules momilactones A and B: Function, biosynthesis, induction and occurrence. Toxins 2023, 15, 241. [Google Scholar] [CrossRef] [PubMed]
- Bonanomi, G.; Sicurezza, M.G.; Caporaso, S.; Esposito, A.; Mazzoleni, S. Phytotoxicity dynamics of decaying plant materials. New Phytol. 2006, 169, 571–578. [Google Scholar] [CrossRef]
- Belz, R.G. Allelopathy in crop/weed interactions—An update. Pest. Manag. Sci. 2007, 63, 308–326. [Google Scholar] [CrossRef]
- Hong, N.H.; Xuan, T.D.; Eiji, T.; Khanh, T.D. Paddy weed control by higher plants from Southeast Asia. Crop Prot. 2004, 23, 255–261. [Google Scholar] [CrossRef]
- Krumsri, R.; Suwunnamek, U.; Homhaul, W.; Laosinwattana, C.; Poonpaiboonpipattana, T. Allelopathic effects of Bidens pilosa var. radiata and its preliminary utilization to control weeds in rice. J. Agric. Technol. 2015, 11, 1875–1886. [Google Scholar]
- Hsueh, M.T.; Fan, C.; Chang, W.L. Allelopathic effects of Bidens pilosa L. var. radiata Sch. Bip. on the tuber sprouting and seedling growth of Cyperus rotundus L. Plants 2020, 9, 742. [Google Scholar] [CrossRef]
- Stevens, G.A., Jr.; Tang, C.S. Inhibition of seedling growth of crop species by recirculating root exudates of Bidens pilosa L. J. Chem. Ecol. 1985, 11, 1411–1425. [Google Scholar] [CrossRef]
- Khanh, T.D.; Cong, L.C.; Xuan, T.D.; Uezato, Y.; Deba, F.; Toyama, T.; Tawata, S. Allelopathic plants: 20. Hairy beggarticks (Bidens pilosa L.). Allelopath. J. 2009, 24, 243–259. [Google Scholar]
- Zhang, K.M.; Shen, Y.; Zhou, X.Q.; Fang, Y.M.; Liu, Y.; Ma, L.Q. Photosynthetic electron-transfer reactions in the gametophyte of Pteris multifida reveal the presence of allelopathic interference from the invasive plant species Bidens pilosa. J. Photochem. Photobiol. B Biol. 2016, 158, 81–88. [Google Scholar] [CrossRef] [PubMed]
- Zhang, K.M.; Shen, Y.; Yang, J.; Miu, X.; Bhowmik, P.C.; Zhou, X.; Fang, Y.M.; Xing, B.S. The defense system for Bidens pilosa root exudate treatments in Pteris multifida gametophyte. Ecotoxicol. Environ. Saf. 2019, 173, 203–213. [Google Scholar] [CrossRef]
- Khasabulli, B.D.; Musyimi, D.M.; George, O.; Gichuhi, M.N. Allelopathic effect of Bidens pilosa on seed germination and growth of Amaranthus dubius. J. Asian Sci. Res. 2018, 8, 103–112. [Google Scholar] [CrossRef]
- Hsu, H.M.; Kao, W.Y. Contrasting effects of aqueous tissue extracts from an invasive plant, Bidens pilosa L. var. radiata, on the performance of its sympatric plant species. Taiwania 2009, 54, 255–260. [Google Scholar]
- Poonpaiboonpipat, T.; Poolkum, S. Utilization of Bidens pilosa var. radiata (Sch. Bip.) Sherff integrated with water irrigation for paddy weed control and rice yield production. Weed Biol. Manag. 2019, 19, 31–38. [Google Scholar] [CrossRef]
- Deba, F.; Xuan, T.D.; Yasuda, M.; Tawata, S. Herbicidal and fungicidal activities and identification of potential phytotoxins from Bidens pilosa L. var. radiata Scherff. Weed Biol. Manag. 2007, 7, 77–83. [Google Scholar] [CrossRef]
- Macías, F.A.; Molinillo, J.M.; Varela, R.M.; Galindo, J.C. Allelopathy—A natural alternative for weed control. Pest Manag. Sci. 2007, 63, 327–348. [Google Scholar] [CrossRef]
- Anh, L.H.; van Quan, N.; Nghia, L.T.; Xuan, T.D. Phenolic allelochemicals: Achievements, limitations, and prospective approaches in weed management. Weed Biol. Manag. 2021, 21, 37–67. [Google Scholar]
- Hussain, M.I.; Danish, S.; Sánchez-Moreiras, A.M.; Vicente, Ó.; Jabran, K.; Chaudhry, U.K.; Branca, F.; Reigosa, M.J. Unraveling sorghum allelopathy in agriculture: Concepts and implications. Plants 2021, 10, 1795. [Google Scholar] [CrossRef]
- Barkosky, R.R.; Einhellig, F.A.; Butler, J.L. Caffeic acid-induced changes in plant-water relationships and photosynthesis in leafy spurge Euphorbia esula. J. Chem. Ecol. 2000, 26, 2095–2109. [Google Scholar] [CrossRef]
- Einhellig, F.A. Mode of action of allelochemical action of phenolic compounds. In Chemistry and Mode of Action of Allelochemicals; Macías, F.A., Galindo, J.C.G., Molino, J.M.G., Cutler, H.G., Eds.; CRC Press: Boca Raton, FL, USA; London, UK; New York, NY, USA; Washington, DC, USA, 2004; pp. 217–238. [Google Scholar]
- Dalton, B.R. The occurrence and behavior of plant phenolic acids in soil environments and their potential involvement in allelochemical interference interactions: Methodological limitations in establishing conclusive proof of allelopathy. In Principals and Practices in Plant Ecology: Allelochemical Interactions; Inderjit, Dakshini, K.M.M., Foy, C.L., Eds.; CRC Press: Boca Raton, FL, USA, 1999; pp. 57–74. [Google Scholar]
- Klessig, D.F.; Malamy, J. The salicylic acid signal in plants. Plant Mol. Biol. 1994, 26, 1439–1458. [Google Scholar] [CrossRef]
- Muhsinah, A.B.; Alsayari, A.; Thajudeen, K.Y.; Asiri, Y.I.; Nanjaian, M. Simultaneous estimation of rutin and quercetin in Bidens pilosa, Convolvulus arvensis and neurada procumbens by RP-HPLC. Res. J. Pharm. Technol. 2020, 13, 3305–3310. [Google Scholar] [CrossRef]
- Parvez, M.M.K.; Yokotani, T.; Fujii, Y.; Konishi, T.; Iwashina, T. Effects of quercetin and its seven derivatives on the growth of Arabidopsis thaliana and Neurospora crassa. Biochem. Syst. Ecol. 2004, 32, 631–635. [Google Scholar] [CrossRef]
- Okada, S.; Iwasaki, A.; Kataoka, I.; Suenaga, K.; Kato-Noguchi, H. Isolation and identification of a phytotoxic substance in kiwifruit leaves. Acta Hortic. 2018, 1218, 207–212. [Google Scholar] [CrossRef]
- Moreland, D.E.; Novitzky, W.P. Effects of phenolic acids, coumarins, and flavonoids on isolated chloroplasts and mitochondria. In Allelochemicals: Role in Agriculture and Forestry; Waller, E.D., Ed.; Series 330; ACS Publications: Washington, DC, USA, 1987; pp. 247–274. [Google Scholar]
- Einhellig, F.A. Mechanisms of action of allelochemicals in allelopathy. In Allelopathy; Inderjit, Dakshini, K.M.M., Einhellig, F.A., Eds.; Series 582; ACS Publications: Washington, DC, USA, 1995; pp. 96–116. [Google Scholar]
- Priestap, H.A.; Bennett, B.C.; Quirke, J.M.E. Investigation of the essential oils of Bidens pilosa var. minor, Bidens alba and Flaveria linearis. J. Essent. Oil Res. 2008, 20, 396–402. [Google Scholar] [CrossRef]
- Gershenzon, J.; Dudareva, N. The function of terpene natural products in the natural world. Nat. Chem. Biol. 2007, 3, 408–414. [Google Scholar] [CrossRef] [PubMed]
- M’sou, S.; Alifriqui, M.; Romane, A. Phytochemical study and biological effects of the essential oil of Fraxinus dimorpha Coss & Durieu. Nat. Prod. Res. 2017, 31, 2797–2800. [Google Scholar] [PubMed]
- Riccobono, L.; Maggio, A.; Bruno, M.; Spadaro, V.; Raimondo, F.M. Chemical composition and antimicrobial activity of the essential oils of some species of Anthemis sect Anthemis (Asteraceae) from Sicily. Nat. Prod. Res. 2017, 31, 2759–2767. [Google Scholar] [CrossRef]
- Winters, A.E.; White, A.M.; Dewi, A.S.; Mudianta, I.W.; Wilson, N.G.; Forster, L.C.; Garson, M.J.; Cheney, K.L. Distribution of defensive metabolites in nudibranchmolluscs. J. Chem. Ecol. 2018, 44, 384–396. [Google Scholar] [CrossRef]
- Sarles, L.; Boullis, A.; Fassotte, B.; Lognay, G.; Verhaeghe, A.; Francis, F.; Verheggen, F.J. Identification of walnut husk (Juglans regia L.) volatiles and the behavioural response of the invasive walnut husk fly, Rhagoletis completa Cresson. Pest. Manag. Sci. 2017, 73, 2100–2104. [Google Scholar] [CrossRef]
- Boncan, D.A.T.; Tsang, S.S.K.; Li, C.; Lee, I.H.T.; Lam, H.M.; Chan, T.F.; Hui, J.H.L. Terpenes and Terpenoids in Plants: Interactions with Environment and Insects. Int. J. Mol. Sci. 2020, 21, 7382. [Google Scholar] [CrossRef] [PubMed]
- Macías, F.A.; Mejías, F.J.; Molinillo, J.M. Recent advances in allelopathy for weed control: From knowledge to applications. Pest Manag. Sci. 2019, 75, 2413–2436. [Google Scholar] [CrossRef] [PubMed]
- Mack, R.M. Predicting the identity and fate of plant invaders: Emergent and emerging approaches. Biol. Conserv. 1996, 78, 107–121. [Google Scholar] [CrossRef]
- Keane, R.M.; Crawley, M.L. Exotic plant invasions and the enemy release hypothesis. Trend. Ecol. Evol. 2002, 17, 164–170. [Google Scholar] [CrossRef]
- Warren, R.J.; Matt Candeias, M.; Labatore, A.; Olejniczak, M.; Yang, L. Multiple mechanisms in woodland plant species invasion. J. Plant Ecol. 2019, 12, 201–209. [Google Scholar] [CrossRef]
- Liu, J.; Xu, G.; Yin, L.; Xu, X.; Armitage, D.W.; Dong, T. Invasive plants exert disproportionately negative allelopathic effects on the growth and physiology of the earthworm Eisenia fetida. Sci. Total Environ. 2020, 747, 141534. [Google Scholar] [CrossRef]
- Taba, S.; Sawada, J.; Moromizato, Z.I. Nematicidal activity of Okinawa Island plants on the root-knot nematode Meloidogyne incognita (Kofoid and White) Chitwood. Plant Soil 2008, 303, 207–216. [Google Scholar] [CrossRef]
- Taba, S.; Ashikaga, K.; Oohama, T.; Ajitomi, A.; Kiyuna, C.; Kinjo, M.; Sekine, K.T. Bidens pilosa extract effects on pine wilt: Causal agents and their natural enemies. Forest Sci. 2020, 66, 284–290. [Google Scholar] [CrossRef]
- Ahmed, S.S.; Abdel-Aziz, S.Y.; Taha, A.; Elhefny, A. Insecticidal effects of two plant extracts of (Bidens pilosa and Rumex dentatus) and neem oil against certain stored grains insects. Egyp. Acad. J. Biol. Sci. F. Toxicol Pest Control 2021, 13, 149–158. [Google Scholar] [CrossRef]
- Mkindi, A.; Mpumi, N.; Tembo, Y.; Stevenson, P.C.; Ndakidemi, P.A.; Mtei, K.; Machunda, R.; Belmain, S.R. Invasive weeds with pesticidal properties as potential new crops. Ind. Crops Prod. 2017, 110, 113–122. [Google Scholar] [CrossRef]
- Li, B.L.; Xie, J.D.; Xie, F.D.; Li, H.Y.; Hu, J.J.; Li, Q.R.; Liang, H.J.; Ao, Z.H.; Fan, Y.W.; Zhang, W.J.; et al. Polyacetylenes from Bidens pilosa Linn., promising insecticides with anti-inflammatory effects on HaCaT cells. Ind. Crops Prod. 2021, 171, 113929. [Google Scholar] [CrossRef]
- Kimura, Y.; Hiraoka, K.; Kawano, T.; Fujioka, S.; Shimada, A. Nematicidal activities of acetylene compounds from Coreopsis lanceolata L. Z. Naturforsch. C 2008, 63, 843–847. [Google Scholar] [CrossRef] [PubMed]
- Campbell, G.; Lambert, J.D.; Arnason, T.; Towers, G.N. Allelopathic properties of α-terthienyl and phenylheptatriyne, naturally occurring compounds from species of Asteraceae. J. Chem. Ecol. 1982, 8, 961–972. [Google Scholar] [CrossRef] [PubMed]
- Chiang, Y.M.; Chuang, D.Y.; Wang, S.Y.; Kuo, Y.H.; Tsai, P.W.; Shyur, L.F. Metabolite profiling and chemopreventive bioactivity of plant extracts from Bidens pilosa. J. Ethnopharmacol. 2004, 95, 409–419. [Google Scholar] [CrossRef]
- Bairwa, K.; Kumar, R.; Sharma, R.J.; Roy, R.K. An updated review on Bidens pilosa L. Der Pharma Chem. 2010, 2, 325–337. [Google Scholar]
- Cortés-Rojas, D.F.; Chagas-Paula, D.A.; Da Costa, F.B.; Souza, C.R.; Oliveira, W.P. Bioactive compounds in Bidens pilosa L. populations: A key step in the standardization of phytopharmaceutical preparations. Rev. Bras. Farmacogn. 2013, 23, 28–35. [Google Scholar] [CrossRef]
- Yang, W.C. Botanical, pharmacological, phytochemical, and toxicological aspects of the antidiabetic plant Bidens pilosa L. Evid.-Based Complement. Altern. Med. 2014, 2014, 698617. [Google Scholar] [CrossRef]
- Silva, L.F.; Fischer, D.C.H.; Tavares, F.J.; Silva, S.S.M.; de Athayde-Filho, F.P.; Barbosa-Filho, J.M. Compilation of secondary metabolites from Bidens pilosa L. Molecules 2011, 16, 1070–1102. [Google Scholar] [CrossRef]
- Lockwood, J.L.; Simberloff, D.; McKinney, M.L.; Von Holle, B. How many, and which, plants will invade natural areas. Biol. Invasions 2001, 3, 1–8. [Google Scholar] [CrossRef]
- Kato-Noguchi, H.; Kurniadie, D. Allelopathy of Lantana camara as an invasive plant. Plants 2021, 10, 1028. [Google Scholar] [CrossRef]
- Kato-Noguchi, H. Involvement of allelopathy in the invasive potential of Tithonia diversifolia. Plants 2020, 9, 766. [Google Scholar] [CrossRef] [PubMed]
- Walther, G.R.; Roques, A.; Hulme, P.E.; Sykes, M.T.; Pyšek, P.; Kühn, I.; Zobel, M.; Bacher, S.; Botta-Dukát, Z.; Bugmann, H. Alien species in a warmer world: Risks and opportunities. Trends Ecol. Evol. 2009, 24, 686–693. [Google Scholar] [CrossRef] [PubMed]
- Mainka, S.A.; Howard, G.W. Climate change and invasive species: Double jeopardy. Integr. Zool. 2010, 5, 102–111. [Google Scholar] [CrossRef] [PubMed]
- Verlinden, M.; Nijs, I. Alien plant species favoured over congeneric natives under experimental climate warming in temperate Belgian climate. Biol. Invasions 2010, 12, 2777–2787. [Google Scholar] [CrossRef]
- Rogers, C.E.; McCarty, J.P. Climate change and ecosystems of the Mid-Atlantic region. Clim. Res. 2000, 14, 235–244. [Google Scholar] [CrossRef]
- Bradley, B.A.; Wilcove, D.S.; Oppenheimer, M. Climate change increases risk of plant invasion in the Eastern United States. Biol. Invasions 2010, 12, 1855–1872. [Google Scholar] [CrossRef]
- Duell, E.B.; Londe, D.W.; Hickman, K.R.; Greer, M.J.; Wilson, G.W.T. Superior performance of invasive grasses over native counterparts will remain problematic under warmer and drier conditions. Plant Ecol. 2021, 222, 993–1006. [Google Scholar] [CrossRef]
- Osaki, S.; Wasaki, J.; Nakatsubo, T. Phenological shifts of the invasive annual weed Bidens pilosa var. pilosa in response to warmer temperature. Plant Ecol. 2022, 223, 1155–1165. [Google Scholar] [CrossRef]
- Kishun, R.; Chand, R. New collateral hosts for Pseudomonas solanacearum. Indian J. Mycol. Plant Pathol. 1987, 17, 237–249. [Google Scholar]
- Phillips, A.J.L. Some common weed species as alternative hosts for Sclerotinia sclerotiorum. Phytophylactica 1992, 24, 207–210. [Google Scholar]
- Guatimosim, E.; Pinto, H.J.; Pereira, O.L.; Fuga, C.A.; Vieira, B.S.; Barreto, R.W. Pathogenic mycobiota of the weeds Bidens pilosa and Bidens subalternans. Trop. Plant Pathol. 2015, 40, 298–317. [Google Scholar] [CrossRef]
- Ferreira, F.A.; da Silva, A.A.; Ferreira, L.R. Effectiveness of nicosulfuron in two formulations, in controlling weeds in maize (Zea mays L.). Ciênc. Agrotec. 1996, 20, 19–24. [Google Scholar]
- Paulo, E.M.; Fujiwara, M.; Terra, M.M.; Martins, F.P.; Pires, E.J.P. Chemical weed control in grapevine Niagara Rosada. Bragantia 1997, 56, 135–143. [Google Scholar] [CrossRef]
- Takano, H.K.; Oliveira, O.S.D., Jr.; Constantin, J.; Braz, G.B.P.; Franchini, L.H.M.; Burgos, N.R. Multiple resistance to atrazine and imazethapyr in hairy beggarticks (Bidens pilosa). Ciênc. Agrotec. 2016, 40, 547–554. [Google Scholar] [CrossRef]
- la Cruz, R.A.; Fernández-Moreno, P.T.; Ozuna, C.V.; Rojano-Delgado, A.M.; Cruz-Hipolito, H.E.; Domínguez-Valenzuela, J.A.; Baro, F.; de Prado, R. Target and non-target site mechanisms developed by glyphosate-resistant hairy beggarticks (Bidens pilosa L.) populations from Mexico. Front. Plant Sci. 2016, 7, 1492. [Google Scholar]
- Feng, P.C.C.; CaJacob, C.A.; Martino-Catt, S.J.; Cerny, R.E.; Elmore, G.A.; Heck, G.R.; Haung, J.; Kruger, W.M.; Malven, M.; Miklos, J.A.; et al. Glyphosate-resistant crops: Developing the next generation products. In Glyphosate Resistance in Crops and Weeds, History, Development, and Management; Nandula, V.K., Ed.; Wiley: New York, NY, USA, 2010; pp. 45–65. [Google Scholar]
- Green, J.M.; Castle, L.A. Transitioning from single to multiple herbicide resistant crops. In Glyphosate Resistance in Crops and Weeds, History, Development, and Management; Nandula, V.K., Ed.; Wiley: New York, NY, USA, 2010; pp. 67–91. [Google Scholar]
- Heap, I. Global perspective of herbicide-resistant weeds. Pest Manag. Sci. 2014, 70, 1306–1315. [Google Scholar] [CrossRef] [PubMed]
- Henckes, J.R.; Cechin, J.; Schmitz, M.F.; Piasecki, C.; Vargas, L.; Agostinetto, D. Fitness cost and competitive ability of ryegrass susceptible and with multiple resistance to glyphosate, iodosulfuron-methyl, and pyroxsulam. Planta Daninha 2019, 37, e019197532. [Google Scholar] [CrossRef]
Figure 1.
Leaves and capitulum of B. pilosa. The pictures were taken by the authors.

Figure 2.
Monospecific stands of B. pilosa. The pictures were taken by the authors.

Figure 3.
Capitulum and cypselae of B. pilosa. The pictures were taken by the authors.

Figure 4.
Possible allelochemicals in B. pilosa. 1: palmitic acid; 2: p-coumaric acid; 3: caffeic acid; 4: ferulic acid; 5: p-hydroxybenzoic acid; 6: vanillic acid; 7: salycilic acid; 8: quercetin; 9: α-pinene; 10: limonene.
Figure 4.
Possible allelochemicals in B. pilosa. 1: palmitic acid; 2: p-coumaric acid; 3: caffeic acid; 4: ferulic acid; 5: p-hydroxybenzoic acid; 6: vanillic acid; 7: salycilic acid; 8: quercetin; 9: α-pinene; 10: limonene.

Figure 5.
Possible compounds involved in the defense function of B. pilosa. 11: 1-phenylhepta-1,3,5-trine, 12: 5-phenyl-2-(1-propynyl)-thiophene, 13: 5-actoxy-2-phenylethinyl-thiophene, 14: icthyothereol acetate.
Figure 5.
Possible compounds involved in the defense function of B. pilosa. 11: 1-phenylhepta-1,3,5-trine, 12: 5-phenyl-2-(1-propynyl)-thiophene, 13: 5-actoxy-2-phenylethinyl-thiophene, 14: icthyothereol acetate.

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Kato-Noguchi, H.; Kurniadie, D. The Invasive Mechanisms of the Noxious Alien Plant Species Bidens pilosa. Plants 2024, 13, 356. https://doi.org/10.3390/plants13030356
AMA Style
Kato-Noguchi H, Kurniadie D. The Invasive Mechanisms of the Noxious Alien Plant Species Bidens pilosa. Plants. 2024; 13(3):356. https://doi.org/10.3390/plants13030356
Chicago/Turabian StyleKato-Noguchi, Hisashi, and Denny Kurniadie. 2024. "The Invasive Mechanisms of the Noxious Alien Plant Species Bidens pilosa" Plants 13, no. 3: 356. https://doi.org/10.3390/plants13030356
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.