

Xanthones: Biosynthesis and Trafficking in Plants, Fungi and Lichens

Department of Environmental Biology, Sapienza University of Rome, P.le Aldo Moro 5, 00185 Rome, Italy
*
Author to whom correspondence should be addressed.
Plants 2023, 12(4), 694; https://doi.org/10.3390/plants12040694
Submission received: 23 December 2022
/
Revised: 24 January 2023
/
Accepted: 26 January 2023
/
Published: 4 February 2023
(This article belongs to the Special Issue Production, Sorting, and Accumulation of Proteins and Metabolites within Plant Cells)
Abstract
:Xanthones are a class of secondary metabolites produced by plant organisms. They are characterized by a wide structural variety and numerous biological activities that make them valuable metabolites for use in the pharmaceutical field. This review shows the current knowledge of the xanthone biosynthetic pathway with a focus on the precursors and the enzymes involved, as well as on the cellular and organ localization of xanthones in plants. Xanthone biosynthesis in plants involves the shikimate and the acetate pathways which originate in plastids and endoplasmic reticulum, respectively. The pathway continues following three alternative routes, two phenylalanine-dependent and one phenylalanine-independent. All three routes lead to the biosynthesis of 2,3′,4,6-tetrahydroxybenzophenone, which is the central intermediate. Unlike plants, the xanthone core in fungi and lichens is wholly derived from polyketide. Although organs and tissues synthesizing and accumulating xanthones are known in plants, no information is yet available on their subcellular and cellular localization in fungi and lichens. This review highlights the studies published to date on xanthone biosynthesis and trafficking in plant organisms, from which it emerges that the mechanisms underlying their synthesis need to be further investigated in order to exploit them for application purposes.
1. Introduction
The designation “xanthone” derives from the Greek word “xanthós”, meaning yellow, and was coined by Schmid in 1855 to indicate the yellow color of the compound isolated from the pericarp of mangosteen (Garcinia mangostana L.), a tropical fruit belonging to the Clusiaceae (or Guttiferae) family.
Xanthone is an aromatic oxygenated heterocyclic molecule, with a dibenzo-γ-pirone scaffold, known as 9H-xanthen-9-one, with the molecular formula of C13H8O2 (Figure 1). The number and class of rings A and B derive from the biosynthetic pathways in higher plants leading to the acetate-derived A-ring (carbons 1–4) and the shikimic acid pathway-derived B-ring (carbons 5–8). Xanthone derivatives consist of slight differences that can be found depending on the nature of the substituents and their localization on the scaffold. Different possible configurations of the two benzene rings and various substituents can be found, leading to higher complexity.
Xanthones are widely distributed in nature and synthesized by several living organisms, even when phylogenetically distant from each other. By January 2016, the Dictionary of Natural Products revealed that natural xanthones are ca. 2000, including their reduced derivatives di-, tetra- and hexahydroxanthones. Plants remain the prevalent source of xanthones, accounting for nearly 80% of natural xanthones. In a pie diagram, shown by Pierre Le Pogam and Joël Boustie, fungi represent 15% while lichens represent the remaining 5% [1]. Algae and bacteria are also able to synthesize xanthones [2,3,4]. Xanthones have been found even in fossil fuels, demonstrating their chemical stability [3,5].
Historically, the first natural xanthone described was gentisin (1,7-dihydroxy-3-methoxyxanthone) isolated from the plant Gentiana lutea L. in 1821 [6] and the first prenylxanthone derivative, tajixanthone, was isolated from the mycelium of Aspergillus stellatus in 1970 [7]. The most studied plant species producing xanthones belong to the Clusiaceae, Hypericaceae, Gentianaceae and Cariophyllaceae families [8,9,10]. Xanthone is well-known to have “privileged structures” because this simple tricyclic compound exhibits wide biological activities such as anticancer, antimicrobial, antifungal, antimalarial, anti-HIV, anticonvulsant, anticholinesterase, antioxidant, anti-inflammatory, and antimalarial activities, among others [11,12,13,14,15,16,17,18,19]. Their interesting structural scaffold and pharmacological importance have encouraged scientists to isolate these compounds from natural products and synthesize them as novel drug candidates. For this reason, biotechnological strategies for xanthone production, such as cell and root cultures grown in bioreactors, and hairy roots, have been studied in recent years in order to obtain efficient protocols for future large-scale production [15,20,21,22,23,24,25,26,27,28].
In view of the importance of xanthone derivatives in medicinal chemistry, we have made efforts to summarize the different classes of xanthones, their biosynthesis and trafficking mechanisms in plant organisms reported in the literature over the last decades.
This review shows the current knowledge of the xanthone biosynthetic pathway with a focus on the precursors and the enzymes involved, as well as on the cellular, tissue and organ localization of xanthones in plants. As far as fungi and lichens are concerned, only biosynthetic pathways have been studied; no information is yet available on the subcellular and cellular localization in these organisms. The selected articles about the topic xanthone biosynthesis, and trafficking in plant organisms, have been screened from Web of Science, Scopus, Pubmed, and Google Scholar to highlight the current advancement and future direction toward completion of the biosynthetic pathways of xanthones.
2. Classification
In recent decades, there has been widespread interest in studying the classification of xanthones, motivated primarily by the great potential of these compounds for their medically useful biological properties. Because of the great diversity of substituents and the discovery and synthesis of new xanthones, their classification by groups has evolved. With the exception of simple xanthones that have only methyl groups attached to the core structure, all other xanthones can be divided into six main groups based on their substituents: oxygenated xanthones, glycosylated xanthones, prenylated xanthones, xantholignoids, bisxanthones, and various xanthones [29,30]. These molecules are biosynthesized and accumulated in various plant organs (leaves, stems, roots, flowers, and fruits) and tissues, and in other organisms [3,31,32,33]. The xanthones identified in plants, fungi, and lichens are reported in Table 1, Table 2 and Table 3.
2.1. Oxygenated Xanthones
Oxygenated simple xanthones are xanthones with simple substituents such as hydroxyl or methoxy, subdivided according to the degree of oxygenation into unoxygenated, mono-, di-, tri-, tetra-, penta-, and hexaoxygenated molecules [232,233]. This class of xanthones is produced both by plants, but also by fungi and lichens (Table 1, Table 2 and Table 3). This group is abundant in many natural products and is the starting point for many more complex xanthones [232,234]. Many monooxygenated xanthones have been isolated from the Gentianaceae family, for example, 2-hydroxyxanthone (Figure 2), 4-hydroxyxanthone and 2-methoxanthone; however, these xanthones have also been found in other plant families such as Clusiaceae, Hypericaceae, Moraceae (Table 1).
More than fifteen deoxygenated xanthones have been reported from plants in the family Clusiaceae. 1,5-dihydroxyanthone, 1,7-dihydroxyanthone, and 2,6-dihydroxyanthone have been found quite widely. Other deoxygenated xanthones such as 1-hydroxy-5-methoxanthone, 1-hydroxy-7-methoxanthone, 2-hydroxy-1-methoxanthone, 3-hydroxy-2-methoxanthone, 3-hydroxy-4-methoxanthone, 5-hydroxy-1-methoxanthone, and 1,2-methylenedioxanthone have been reported from plants belonging to Polygalaceae and Calophyllaceae families (Table 1) [97,193]. The natural trioxygenated xanthones 1,3-dihydroxy-5-methoxy-xanthone-4-sulfonate and 5-O-β-D-glucopyranosyl-1,3-dihydroxy-xanthone-4-sulfonate were isolated from Hypericum sampsoni Hance., and Centaurium erythraea [137,176]. Tetraoxygenated xanthones have been reported mainly from plants in the families Gentianaceae, Clusiaceae, and Polygalaceae. The xanthones isolated from Polygala vulgaris L. were 7-chloro-1,2,3-trihydroxy-6-methoxanthone, 1,3,5,6-, 1,3,5,7-, and 1,3,6,7-tetrahydroxanthones [196].
Xanthones isolated from Gentiana rhodantha Franch. were methylated pentaoxygenated xanthones, namely 1,8-dihydroxy-2,3,7-trimethoxanthone, 5,6-dihydroxy-1,3,7-trimethoxanthone, 1,7-dihydroxy-2,3,8-trimethoxanthone, and 3,8-dihydroxy-1,2,6-trimethoxanthone [147]. In addition, hexoxygenated xanthones such as 8-hydroxy-1,2,3,4,6-pentamethoxanthone, 1,8-dihydroxy-2,3,4,6-tetramethoxanthone and 1-hydroxy-3,5,6,7,8-pentamethoxyxanthone have been isolated from two species of Centaurium [4,137,138] (Table 1).
2.2. Xanthone Glycosides
Xanthone glycosides are synthesized in higher plants and mainly found in the families Gentianaceae and Polygalaceae (Table 1). These xanthones are known to have a glycosidic residue associated with a C- or O-. Many xanthones of this group have the glycosidic residue linked to C-6 carbon and may consist of xylose, glucose, or epiose. The presence of two glycosidic residues can also be observed, the second generally being linked to the C-2 of the main structure [235]. Mangiferin and isomangiferin are the most common C-glycosides (Figure 3a). Mangiferin (2,-C-β-Dglucopyranosyl-1,3,6,7-tetrahydroxyxanthone) is of widespread occurrence in angiosperms and ferns and was first isolated from Mangifera indica [236]. Natural xanthone O-glycosides are restricted to the family Gentianaceae and in this review gentiacauloside from Gentiana acaulis L., gentioside from G. lutea, and swertianolin (Figure 3b) from Swertia japonica Makino have been reported [143,144,156].
2.3. Prenylated Xanthones
There is considerable variability in the classification of prenylated xanthones; in fact, it is possible to observe the presence of the prenyl group in different positions of the basic structure. These groups can have carbon chains consisting of a single prenyl group, composed of five carbons (e.g., allanxanthone A, Figure 4), or multiples such as 10 or 15 carbons. In addition, prenylated xanthones can have varying degrees of oxygenation. These molecules are predominantly extracted from species belonging to Hypericaceae family, such as H. sampsonii (Table 1), from which an antiviral xanthone against hepatitis B, hyperxanthone, has been isolated [176]. Numerous prenylated xanthones have also been isolated from fungi belonging to the genera Actinomadura, Emericella and Paecilomyces [200,201,204,212]. Other prenylated xanthones isolated from fungi are reported in Table 2.
2.4. Bisxanthones
Bisxanthones were identified in higher plants, fungi, and lichens [3,165], and reported in Table 1, Table 2 and Table 3. Their chemical structure consists of a 9H-xanthen-9-one dimer with several substituent groups. These include jacarelhyperols A and B isolated from the aerial parts of Hypericum japonicum Thunb and the dimeric xanthone, and globulixanthone E (Figure 5), from the roots of Symphonia globulifera L.f. [132,133,165]. Many xanthones belonging to this group have also been isolated in lichens, particularly in the genera Teloschistale and Diploicia (Table 3). In addition, tetrahydroxanthone dimeric C2-C2 dicerandrols A, B, and C have been isolated from the fungus Phomopsis longicolla [237] (Table 2). A dimeric C4-C4 xanthone isolated from the root bark of Centaurium erythrea Raf. has remarkable antimicrobial properties against gram positives such as Staphylococcus spp. [139].
2.5. Xantholignoids
The group of xantholignoids, characterized by a connection between xanthones and lignin (conifer alcohol) structures, is limited. Cadensins A and B were isolated from Caraipa densifora Kubitzki. Kielcorin (Figure 6) was initially obtained from Hypericum species but has also been isolated from Vismia guaramirangae Huber [181], Kielmeyera variabilis Mart. & Zucc. [99], and Hypericum canariensis L. (Table 1), [163]. In fungi and lichens, this class of xanthones is present in many genera (Table 2 and Table 3).
2.6. Miscellaneous Xanthones
Miscellaneous xanthones are defined for all xanthone derivatives that cannot be classified into other groups. They are found in the kingdom of plants and fungi (Table 1, Table 2 and Table 3). Among them, we include xanthofulvin (Figure 7) and vinaxanthone, isolated from Penicillium spp. (Table 1) SPF-3059 [238], and thioxanthones and azaxanthones [239].
3. Xanthone Biosynthesis
3.1. Xanthone Biosynthesis in Plants
3.1.1. Shikimate Pathway
Xanthones are synthesized in plants via the shikimate pathway with the contribution of the acetate (or polyketide) pathway. Shikimate links carbohydrate metabolism, glycolysis and pentose phosphate pathway, to aromatic compound biosynthesis (Figure 8). The shikimate pathway occurs in green and non-green plastids, thus dependently or independently from light [240]. However, it is known that non-photosynthetic tissues are partially supplied with amino acids transported by the phloem, so production does not occur exclusively within the cell; it can also occur in other tissues or organs, and then transport to other locations occurs [241]. Moon and Mitra [149] showed that shikimate dehydrogenase (SKD) and shikimate kinase (SK), key enzymes of the shikimate pathway, are activated after elicitation by a Ca2+-mediated H2O2 generation, leading to a consequent increase in the xanthone biosynthesis, giving further confirmation to the role of xanthones as defense metabolites as described by numerous articles on the subject [242]. This study revealed for the first time the link between ROS and the pathways involved in xanthone biosynthesis.
After the shikimate pathway, xanthone biosynthesis can proceed with an L-phenylalanine-dependent pathway, as in Hypericum androsaemum L. [243], G. mangostana, and G. lutea [145,244] or an L-phenylalanine-independent pathway, as in Swertia chirata Buch.-Ham. ex Wall. [151], C. erythraea [243,245,246] and Hoppea fastigiata Griseb. [151]. Both the phenylalanine-dependent and phenylalanine-independent pathways pass through the production of 2,3′,4,6-tetrahydroxybenzophenone (2,3′,4,6-THB), which is therefore a central intermediate in the biosynthesis of xanthones (Figure 8).
3.1.2. Phenylalanine-Dependent Pathway
In the phenylalanine-dependent pathway, shikimate forms the amino acid phenylalanine through numerous reactions occurring in two different cell compartments, plastid and cytosol [247,248] (Figure 8). Phenylalanine is biosynthesized from chorismate, the final product of the shikimate pathway. In plastids, chorismate is converted to prephenate which in turn is transaminated producing arogenate. This compound is then dehydrated/decarboxylated to phenylalanine which is then transported to cytosol by the plastidial cationic amino acid transporter (pCAT) [249]. In plants, the arogenate pathway is the predominant route for phenylalanine biosynthesis although another pathway, more common in microorganisms [250], has been described. This route, which has yet to be clarified, involves phenylpyruvate, another product downstream of prephenate. Phenylpyruvate may originate from prephenate in plastids by the action of arogenate dehydratases (ADTs) [251,252] or in the cytosol, requiring a cytosolic pool of prephenate supposedly formed by the action of a cytosolic chorismate mutase (CM) from chorismate previously synthesized in the plastid and then transported to the cytosol [249]. Indeed, as described by Yoo and co-workers [253] in Petunia hybrida E.Vilm, prephenate seems to be produced in the plastid but converted to phenylalanine in the cytosol by a phenylpyruvate aminotransferase (PPY-AT), which preferentially uses prephenate as a substrate, suggesting that this alternative route to phenylalanine biosynthesis is also active in the plants. Once in the cytosol, the amino acid is converted to trans-cinnamic acid by the action of the enzyme phenylalanine ammonia-lyase (PAL), which catalyzes the deamination. Trans-cinnamic acid is the substrate of cinnamate-CoA ligase (CNL), which leads to cinnamoyl-CoA. Cinnamoyl-CoA is an intermediate from which benzoyl-CoA is formed as a result of three reactions that are catalyzed by the enzymes cinnamoyl-CoA hydratase/lyase (CHL), benzaldehyde dehydrogenase (BD), and benzoate-CoA ligase (BZL). BZL expression has been demonstrated to increase before xanthone biosynthesis, when the plant is exposed to elicitation, suggesting its role in the biosynthetic pathway upstream of xanthones. Singh and co-workers [254] have shown in Hypericum calycinum L. that BZL is localized in both peroxisomes and cytosol, indicating the activation of the CoA-dependent non-β-oxidative pathway for benzoyl-CoA production. The activation of this pathway was previously demonstrated at the biochemical level in Hypericum androsaemum L. cell cultures [255]. Furthermore, it is hypothesized that the enzyme is purely involved in the phenylalanine-dependent pathway having benzoic acid as a preferential substrate.
The subsequent reaction is catalyzed by benzophenone synthase (BPS), a type III polyketide synthase, which condenses the benzoyl-CoA molecule with three malonyl-CoAs originating 2,4,6-trihydroxybenzophenone (2,4,6-THB). BPS in H. androsaemum and G. mangostana has benzoyl-CoA as a specific substrate, suggesting that the phenylalanine-dependent pathway is the one followed for xanthone production in these species [243]. CYP81AA, a cytochrome P450 (CYP) monooxygenase that possesses benzophenone 3′-hydroxylase (B3′H) activity, converts 2,4,6-THB to 2,3′,4,6-THB. Thus, these compounds are the precursors of various benzophenones and xanthones. The two main precursors of xanthones are formed from 2,3′,4,6-THB ring closure. 1,3,5-trihydroxyxanthone (1,3,5-THX) and 1,3,7-trihydroxyxanthone (1,3,7-THX) originate from oxidative phenol coupling reaction that occurs either at the ortho or para position of the 3′-OH group, respectively. Cyclization to 1,3,5-THX and 1,3,7-THX depends on the species [146,256,257,258]. These reactions are catalyzed by two xanthone synthases belonging to the CYP oxidases [256]. They are now known as 1,3,5-THX synthase (CYP81AA2) and 1,3,7-THX synthase (CYP81AA1), respectively [258,259]. One of the two pathways could be used preferentially by a species, but it has been shown from transcriptome databases of Hypericum spp. that genes for both CYPs are present, so both isomers of the enzyme could be synthesized in a species in response to certain signals [145,258]. Kitanov and Nedialkov [260] proposed that 1,3,7-THX is generated from 2,4,5′,6-tetrahydroxybenzophenone-2′-O-glucoside (hypericophenonoside) in H. annulatum firstly removing the glucoside group by hydrolysis before cyclization. Many different xanthones will then be produced from these precursors, although to date the biosynthetic pathways of many of them are only assumed.
3.1.3. Phenylalanine-Independent Pathway
In the phenylalanine-independent pathway, the biosynthetic pathway originates from shikimate to produce 3-hydroxybenzoic acid in the cytoplasm without the involvement of phenylalanine (Figure 8). To date, how shikimate leaves plastids and which enzymes are responsible for the conversion to 3-hydroxybenzoic acid is unknown. The 3-hydroxybenzoic acid is then thioesterified by 3-hydroxybenzoateCoA ligase (3-BZL) to form 3-hydroxybenzoyl-CoA, and subsequent condensation by BPS leads to the formation of 2,3′,4,6-THB. In Centaurium species, 3-BZL enzyme has been shown to have 3-hydrobenzoic acid rather than benzoic acid as a preferred substrate, suggesting that the phenylalanine-independent pathway is the one followed in these species [261]. The biosynthetic pathway continues as described for the phenylalanine-dependent route.
Although the phenylalanine-dependent pathway is more studied and it is assumed that most xanthones are produced downstream of phenylalanine or indistinctly by both the phenylalanine-dependent and independent pathways, some xanthones such as 1,3,5,8-tetrahydroxy xanthone and 1,5,7-trihydroxy-3-methoxy xanthone appear to be produced only through the phenylalanine-independent pathway [151] (Figure 8).
3.1.4. Xanthone Derivatives of 1,3,5-Trihydroxyxanthone
Xanthone-6-hydroxylases (X6H), a CYP-dependent monooxygenase, has been shown to hydroxylate 1,3,5-THX to 1,3,5,6-tetrahydroxyxanthone (1,3,5,6-THX) in H. androsaemum and C. erythraea [262]. In S. chirata, the hydroxylation of 1,3,5-THX occurs at the C-8 position of the ring, originating 1,3,5,8-tetrahydroxyxanthone (1,3,5,8-THX) [263]. On the other hand, Beerhues and Berger [245], studied the elicited cell cultures of C. erythraea and C. littorale proposing a direct formation of 1,5-dihydroxy-3-methoxyxanthone from 1,3,5-THX. Moreover, the authors proposed a biosynthetic pathway downstream 1,3,5-THX in the cell cultures of these species which produce xanthones such as 1,5-dihydroxy-3-methoxyxanthone, 1-hydroxy-3,5,6,7tetramethoxyxanthone, and 1,8-dihydroxy-3,5-dimethoxyxanthone. However, the enzymes involved in these reactions have not been identified [10,245].
3.1.5. Xanthone Derivatives of 1,3,7-Trihydroxyxanthone
Many more xanthones derive from 1,3,7-THX. X6H is also involved in the formation of 1,3,7-THX derivatives [264,265]. Indeed, the hydroxylation of 1,3,7-THX forms 1,3,6,7-tetrahydroxyxanthone (1,3,6,7-THX) in H. androsaemum and G. mangostana [262] and potentially resides in the endoplasmic reticulum [258].
1,3,7-THX is proposed to be a precursor compound for prenylated xanthones, such as rubraxanthone from Garcinia [266,267] and Calophyllum species [9,268,269] and scortechinone B from Garcinia scortechinii King [270,271], as well as simple xanthones, such as 1,7-dihydroxy-3-methoxyxanthone (gentisin) and 1,3-dihydroxy-7-methoxyxanthone (isogentisin) from G. lutea [146,272]. In G. mangostana, γ-mangostin is proposed to be generated by prenylation of the 1,3,6,7-tetrahydroxyxanthones, and α-mangostin by the subsequent O-methylation [273,274]. Another pathway which produces patulone, hyperxanthone E, and hyperixanthone A starting from 1,3,6,7tetrahydroxyxanthones has been reported in Hypericum spp. [275,276]. Two enzymes involved in these reactions are known: 8-prenylxanthone-forming prenyltransferase (PT8PX) and patulone-forming prenyltransferase (PTpat). The former has prenylation activity and is mainly localized at the envelope of the chloroplast [276] (Figure 8). The latter is also a prenyltransferase which prenylates the reaction product of the previous reaction, 8-prenyl-1,3,6,7tetrahydroxyxanthone, and produces patulone [275,276]. Other xanthones are supposed to be formed from this route, such as hyperxanthone A and E, but the enzymes involved are unknown [275,276].
Among the glycosylated xanthones, norathyriol 6-O-glucoside (tripteroside or Xt1) and norathyriol-6-O-(6′-Omalonyl)-glucoside (Xt2) have recently been characterized at the molecular level [33]. The enzymes responsible for the reaction that produces these xanthones from 1,3,6,7-THX are norathyriol 6-O-glucosyltransferase (StrGT9) and malonyl-CoA acyltransferase (StrAT2). StrGT9 glucosylates 1,3,6,7-THX to Xt1, which in turn is malonylated in the presence of malonyl-CoA to Xt2 by StrAT2 (Figure 8).
Mangiferin is a well-studied C-glucoside xanthone. A route for its biosynthesis was proposed by Fujita and Inoue [40] and Chen and co-workers [277] in Anemarrhena asphodeloides Bunge and M. indica, respectively, and reviewed by Ehianeta and co-workers [278]. The results suggest that mangiferin and related xanthone C-glycosides are produced through an intermediate, maclurin 3-C-glucoside, which is converted to mangiferin and isomangiferin by C-glycosyltransferase (CGT).
3.2. Xanthone Biosynthesis in Fungi and Lichens
Xanthones are highly unique in fungi and lichens, legitimating, compared to plants, the vastness of chemical diversity of these “privileged structures” with a pronounced biological activity. The biosynthesis of xanthones in fungi and lichens is a topic of considerable interest, but up to now only a few studies report detailed information on the metabolic pathways and trafficking in these organisms.
Xanthones from fungi result from biosynthetically distinct pathways compared to those in plants. The synthesis of the xanthones in fungi has been suggested in 1953 by Birch and Donovan [279], and studied in detail in 1961 by Roberts through the radiolabeled acetate feeding experiments [280] that showed polyketides are the biosynthetic precursors of the xanthone core in Penicillium and Aspergillus species. Acetyl-CoA is the starter compound cyclized to form anthraquinone emodin (6-methyl-1,3,8-trihydroxyanthraquinone), which in turn produces chrysophanol as a result of 6-deoxygenation. The quinone ring of chrysophanol is cleaved by enzymes MdpL (Baeyer–Villiger oxidase) and MdpJ (glutathione S transferase) leading to the formation of thioester intermediate, which is in turn reduced by the action of an oxidoreductase MdpK to benzophenone alcohol. The latter compound is dehydrated producing 1-hydroxy-6-methyl-8-hydroxymethylxanthone which is further hydroxylated to 1,7-dihydroxy-6-methyl-8-hydroxymethylxanthone by MdpD, a monooxygenase [281]. As observed in Aspergillus nidulans, this compound then undergoes two consecutive prenylations (O-prenylation and C-prenylation) by prenyltransferase enzymes XptB and XptA. The O-prenylation catalyzed by XptB forms variecoxanthone Awhich undergoes a C-prenylation step by XptA leading to the final product emericellin, which in turn cyclizes due to the oxidoreductase XptC to shamixanthone and epishamixanthone, as reported in Aspergillus variecolor and Aspergillus rugulosus, respectively [282,283]. Other prenylated xanthones were identified in Ascomycetes fungi as Aspergillus and Penicillium genera. In other fungi, such as Paecilomyces variotii, Acetyl-CoA/Malonyl-CoA are cyclized by AgnPKS to form a PKS-bound octaketide that is hydrolyzed in the atochrysone carboxylic acid by AgnL7. The atochrysone carboxylic acid is decarboxylated to emodin anthrone by AgnL1 and then oxidized to emodin by AgnL2. Emodin can be reduced by AgnL4 to dihydroquinone and then to hydroxyketone by AgnL6. AgnL8 is responsible for dehydration reactions leading to chrysophanol. The following Baeyer–Villiger oxidation carried out by monooxygenase AgnL3 forms monodictylactone, whose hydrolyzation to monodictyphenone and reduction to dihydro-monodictyphenone leads to Agnestin C and a rearrangement to either A and B, which are interconvertible [284]. The xanthone biosynthetic pathway in fungi is reported in Figure 9. In a recent review, Khattab and Farag 2022 [281] widely describe the unique structural characteristics of dimeric, dihydro-, tetrahydro, or hexahydroxanthones as well as prenylated and chlorinated xanthones in terrestrial and marine fungi. Fungi-derived lichens are reported in Table 2.
Lichens are symbiotic organisms that are composed of fungi and algae and/or cyanobacteria. They produce a variety of characteristic xanthones metabolites with various biological properties including antimicrobial, antiviral, and antitumor activities.
The biosynthesis of xanthones proceeds through the polyacetate/polymalonate pathway, where the single polyketide chain undergoes ring-closure, and possibly through a benzophenone intermediate gives two distinct series of xanthones, depending on this folding pattern. In the first pathway, the single polyketide undergoes the aldol condensation and Claisen-type cyclization to form a benzophenone intermediate that might spontaneously dehydrate to obtain the central pyrone core. This biosynthetic pathway gives rise to the common oxygen substitution pattern of lichexanthone and norlichexanthone characterized by a methyl group in the 8-position (1,3,6-trihydroxy-8-methylxanthone) [2]. A limited number of structures derived from a biosynthetically distinct pathway gives the ravenelin skeleton characterized by a methyl group in the 3-position. This biosynthetic pathway starts with the widespread anthraquinone emodin as a precursor. The cleavage of the hydroxyl group on C-6 of the emodin leads to chrysophanol, as observed in the fungus Pyrenochaeta terrestris [285,286]. After the oxidative ring opens, the hydroxyl group on C-4 is then incorporated and an aryl epoxidation across an A-ring edge of chrysophanol yields an intermediate which has lost its A-ring aromaticity, as proposed by Henry and Townsend [287]. This intermediate, which is stabilized by a hydrogen bond between its newly formed phenol group and the neighboring quinone group, recovers its A-ring aromaticity to grant islandicin as a shunt product. Alternatively, a second oxidation, most likely by the same P450 oxygenase, occurs to afford a Baeyer–Villiger cleavage of the central quinone ring to yield an ortho carboxybenzophenone that might follow several metabolic fates [2].
A first possibility is the 1,4-addition of a B-ring phenol to the A-ring dienone followed by dehydration and decarboxylation to access ravenelin-like xanthones after a final oxidation [287]. These reactions lead to xanthones displaying an archetypical 1,4,8-trihydroxy-3-methylxanthone skeleton. A second metabolic pathway, granting access to eumitrins and secalonic acids, assumes the methylation of the carboxy group to prevent its subsequent elimination after a similar 1,4-addition. Finally, a subsequent 1,2-addition to the benzophenone intermediate leads to further cores similar to that of tajixanthone produced by A. variecolor, a skeleton thus far unknown from lichens. The xanthone biosynthetic pathway in lichens is reported in Figure 10.
From this point, the other compounds differ in the position and extent of substitution, including hydroxylation, methylation of these hydroxyl groups, and chlorination [2,3]. Lichen-derived xanthones are reported in Table 3.
It is interesting to underline that even though xanthones from free-living fungi are well known, an algae-fungus collaboration has been suggested for the synthesis of several lichen xanthones. As observed in lichen Lecanora dispersa [222] or in Lecanora rupicola [288], when the fungus is cultivated in the absence of the alga, the xanthone production is diverted to other secondary metabolites being produced (e.g., depsidones such as pannarin and related compounds). However, lichens offer the widest diversity of compounds in the fungal realm, even though their bioactivities remain under-investigated despite being widely considered a promising class of compounds exerting pleiotropic pharmacological activities.
4. Organ and Tissue Localization of Xanthones and Their Possible Functions in Plants
From the chemical studies, data shown in the literature made it possible to obtain information about the different types of plant xanthones identified in plants and which are the organs of accumulation. On the contrary, little is known about the tissue sites of xanthones biosynthesis in plants. Tissue localization of xanthone biosynthesis has been investigated in a few studies which are described below. Immunofluorescence localization of polyketide synthase key enzymes of flavonoids, and xanthone biosynthesis, namely chalcone, and benzophenone synthases were carried out in the leaves [289] of H. perforatum. Benzophenones are metabolized to xanthones through benzophenone synthase (BPS) activity [258,290]. Upon mutation in a single active site position, H. androsaemum BPS formed phenylpyrones [291]. Berkleir and collaborators [289] studied cross-sectioned leaves of H. perforatum incubated with anti-His6–BPS IgG and anti–His6-CHS IgG at various developmental stages. Immunofluorescence localization of both CHS and BPS was in the mesophyll and the intensity of immunofluorescence varied with leaf age. Maximum immunolabeling of CHS was observed in approximately 0.5 cm long leaves, while BPS was undetectable. The CHS-specific fluorescence rapidly decreased in more developed leaves (1 cm long), which instead presented high levels of BPS immunofluorescence. Unlike leaves, the roots appear to be the richest organs in xanthones [15,178,292], which is consistent with the high level of BPS transcription found in both H. sampsonii and H. perforatum roots [172,178]. In situ detection of BPS transcripts and proteins has also been carried out in situ mRNA hybridization and indirect immunofluorescence detection. Moreover, label-free localization of xanthones was studied by AP-SMALDI-FT/MS imaging [172]. It should also be noted that the concentration of xanthone precursors, particularly polyprenylated benzophenones, is very high in the root system [172]. BPS protein was immunodetected in the root exodermis and the endodermis but not in the epidermis. The exodermis and the endodermis are the outermost and innermost layers of the root cortex. The authors emphasize that these tissues are structurally and functionally related [293,294], sharing the barrier role and controlling the radial transport of water and solutes in the root. As is well known, both tissues also play a role in defense against pathogens. Tocci and co-workers [22] demonstrated that root cultures of H. perforatum treated with the elicitor chitosan, which mimics the fungal pathogen attack, showed high levels of xanthone content. In a subsequent study performed through the 1H-NMR-based metabolomics approach, it has been observed that H. perforatum root cultures elicited by chitosan, and under “overcrowding stress”, produced a yield of total xanthones ten times higher compared to the previous study. Moreover, in this study the brasilixanthone B has been isolated and identified in H. perforatum for the first time [295]. Strengthening the defensive role of xanthones, Huang and collaborators in H. sampsonii [178,296] showed that cDNAs encoding HsBPS and HsCHS were differentially regulated in the vegetative and in reproductive stages. In the vegetative stage, HsBPS was highly expressed in the roots; its transcript level was approximately 100 times higher than that of HsCHS, whereas the young leaves contained higher transcript levels of HsCHS. In the reproductive stage, maximum HsCHS expression was detected in flowers, the transcript level being approximately five times higher than that of HsBPS. The inverted situation with a 10-fold difference in the expression levels was observed in the fruits.
To prove the defensive role of xanthones against fungal pathogens, Crockett and co-workers [297] demonstrated that 1,6-dihydroxy-5-methoxy-4′,5′-dihydro-4′,4′,5′-trimethylfurano-(2′,3′:3,4)-xanthone, isolated by H. perforatum roots, inhibited the plant pathogenic fungi Phomopsis obscurans and P. viticola.
As shown in Table 1, several papers are published on xanthone production in other genera and species belonging to other families besides the Hypericaeae; however, organ and tissue localization studies are few.
Xanthones in Calophyllum inophyllum L. (Calophyllaceae) roots have been studied [298]. Different xanthones were accumulated in root bark and in root hearthwood. A new xanthone named caloxanthone D has been found in the bark and caloxanthone E in the hearthwood. Chemical investigation of dichloromethane and ethyl acetate extracts from the stem and root bark of Trema orientalis L. (Ulmaceae) led to the isolation of 16 compounds, including four xanthones [101].
Xanthones have also recently been found in the flowers of Japanese Gentiana cultivars, which show red petals rather than blue. The authors characterized the pigments responsible for the red color in these cultivars revealing the presence of cyanidin-based anthocyanins and xanthones. In particular, two compounds have been identified for the xanthones: norathyriol 6-O-glucoside and norathyriol-6-O-(60 -O-malonyl)-glucosid. These compounds contributed to the red color of flowers [33].
Xanthone content has also been studied in the leaves of Coffea pseudozanguebariae Bridson (family Rubiaceae) wild-grown plants, collected at different developmental stages [197]. The authors showed that C-glycosylated xanthones, i.e., mangiferin 1 and isomangiferin 2, represented 6% of the dry mass in the young leaves, while they were much lower in the older leaves; these results support the hypothesis that the xanthones play a defense role in the most delicate phases of the leaf development, as demonstrated in Hypericum. In a subsequent study, Talamond and co-workers [32] using a multiphoton fluorescence imaging, demonstrated that mangiferin, identified as the spectrum emission, was localized in the upper epidermis and in some mesophyll cells.
Still in Coffea, leaf phenolic composition has been studied in 23 species and focus on mangiferin content [299]. Leaves of Coffea arabica L., Coffea canephora Pierre ex A. Froehner, Coffea eugenioides S. Moore and C. pseudozanguebariae were sampled at different developmental stages. In particular, leaves were collected at three stages: (1) young leaves from the apex; (2) leaves from the first node below the apex; (3) leaves from the second node below the apex. Microscopic observations carried out with UV light (filter UV-1A: 365 nm excitation filter) revealed yellow autofluorescence of mangiferin and its preferential localization in palisade and spongy parenchyma tissues of C. pseudozanguebariae leaves. In contrast, mangiferin was absent in C. canephora and present at low concentration in C. arabica. In the same study, xanthone localization in the fruits has been studied. Samples were obtained from C. pseudozanguebariae and C. canephora and collected at three developmental stages: (1) fruit when green with a partially formed seed (immature); (2) fruit when yellowish green, pericarp (exocarp, mesocarp, endocarp) and (3) fruit when reddish yellow. An intense yellow autofluorescence was shown in the cells of the exocarp and the external layers of the mesocarp of young green fruits, indicating high content of mangiferin. Mangiferin was absent in the seeds and endocarp of the three species examined. The authors speculate that the mangiferin accumulation within the fruit could be associated with photosynthetic tissues; in fact, the receptacle and young fruit are green and photosynthetic, while the ovary is not.
Moreover, in the same study, Campa and collaborators [299] demonstrated that mangiferin is accumulated in the leaves and fruits of seven of 23 Coffea species (24 taxa) studied, including two hybrids (C. arabica ‘Laurina’ and C. heterocalyx Stoff. cf.), originating from different localities in Africa. On the contrary, none of the Madagascan species contained mangiferin, perhaps due to the different environmental conditions. A relationship between mangiferin accumulation, altitude, and UV levels was speculated. The peripheral localization of mangiferin in Coffea plant organs and its association with photosynthetic tissue strengthens the hypothesis of a protective action against UV-radiation. Thus, in addition to a defense role against pathogens, xanthones also appear to play a role toward environmental factors. Soil conditions and altitude influenced xanthone content of H. perforatum roots. Young wild plants of H. perforatum subsp.angustifolium collected in two areas (Lazio Region, Italy) at two different altitudes (68 and 453 m above sea level) and in different soils (calcareous and volcanic) showed different amounts and quality of xanthones [13].
5. Recent Insight on Biological Activities of Xanthones
The ability of xanthones to bind to multiple and unrelated classes of protein receptors as high affinity ligands, allow these molecules to be considered “promiscuous binders”, because they are able to interfere with a variety of biological targets exerting pronounced pharmacological activity against several diseases [17]. This ability is related with some special molecular features, such as the presence of the heteroaromatic tricyclic ring system being predominantly planar and rigid, the carbonyl group at the central ring capable of several interactions, the biaryl ether group contributing to the electronic system, and the xanthone core that accommodates a vast variety of substituents at different positions. Their interesting structural scaffold and pharmacological importance have encouraged scientists to isolate these compounds from natural products and synthesize them as novel drug candidates in the field of medicinal chemistry. Numerous naturally occurring and synthetic xanthone derivatives have been reported in the literature with several beneficial heterogeneous pharmacological activities. According to several authors, anticancer, antimicrobial, antimalarial, anti-HIV, anticonvulsant, anticholinesterase, antioxidant, anti-inflammatory, as well as an inhibitory activity on different enzymes, including a-glucosidase, topoisomerase, protein kinase C, miRNA, intestinal P-glycoprotein, acyl-CoA:cholesterol acyltransferase, xanthine oxidase, and aromatase have been attributed to xanthones [16,30]. All xanthone classes have already demonstrated cytotoxic effects. An antiproliferative activity of xanthone carbaldehyde derivatives, prenylated xanthones, and chiral xanthone derivatives has been demonstrated in MCF-7 (breast adenocarcinoma), KB 3.1 (squamous cell oral carcinoma), A375-C5 (melanoma), and NCI-H460 (non-small cell lung cancer) cell lines [303,304,305]. The growth inhibitory effect on human tumor cell lines was dependent on the nature and position of substituents on the xanthone scaffold and the stereochemistry of the xanthones. Among them, the major group of naturally occurring xanthones are prenylated xanthones, in which the presence of prenyl groups in key positions on the xanthone nucleus can influence the physicochemical properties, namely lipophilicity, and affect the interaction with the biological targets exerting several biological activities, such as antitumor, anti-inflammatory, and human lymphocyte proliferation inhibitory effects [306]. In this context, C-prenylated xanthones are able to decrease cellular proliferation and induce S-phase cell cycle arrest and apoptosis, increasing cleaved PARP and Bid levels and decreasing Bcl-xL in K-562 cells [307] in MCF-7 (breast adenocarcinoma), NCI-H460 (non-small cell lung cancer), A375- C5 (melanoma), and HL-60 (acute myeloid leukemia) cell lines [30]. It is known that the p53 tumor suppressor is a major transcription factor with a crucial role in cell proliferation and death. The activity of p53 is commonly lost in cancers either by mutation in the TP53 gene, or by inactivation due to the overexpression of the main endogenous negative regulator, murine double minute 2 (MDM2). Therefore, restoration of p53 activity by inhibiting the MDM2-p53 interaction represents an appealing therapeutic strategy for many wild-type p53 tumors with overexpressed MDM2 [308]. It has been demonstrated that prenylated xanthone α-mangostin and gambogic acid are inhibitors of MDM2-p53 interaction [309], while the pyranoxanthone has shown a promising growth inhibitory activity as a putative inhibitor of MDM2-p53 interaction in human tumor cells expressing wild-type p53 and overexpressed MDM2 [308]. Moreover, the oxygenated xanthones characterized by simple substituents such as hydroxyl, methoxy, or methyl groups showed antioxidant properties implicating cancer chemopreventive [310], hepatoprotective [311], antifungal [312], antibacterial [313], and anti-obesity [314] actions involving targets such as monoaminoxidase (MAO) [315], P-glycoprotein (P-gp) [316], protein kinase C (PKC) [317], and tyrosinase [318]. Concerning MAO studies, xanthones acted preferentially as MAO-A competitive, reversible inhibitors with IC50 values in the micro- to nanomolar range, and 1,5- dihydroxy-3-methoxyxanthone with an IC50 of 40 nM for MAO-A emerged as the most active inhibitor. Along with xanthonolignoids, 3,4-dihydroxy xanthone derivatives with synthetic intermediates of 3,4-dihydroxy-2-methoxyxanthone and 2,3-dihydroxy-4-methoxyxanthone were found to be the most potent lignoids, with promising antiproliferative and apoptotic effects in leukemia cell lines [30,307]. 1,2-dihydroxyxanthone, initially considered promising for its effect against melanoma [319], due to its catechol structure peri to carbonyl, is also the most promising antioxidant agent for its chelating properties, stability, phototoxicity, cytotoxic effect on a human keratinocyte cell line [320], and its modulatory effects on the activity of the THP-1 macrophage cell line, namely cytokine production [321]. Rosa et al. [318] found that the partial negative surface area, the relative number of oxygen atoms, and the substitution pattern of the 1-methyl-3,4,6-trihydroxyxanthone contributed to the tyrosinase inhibitory activity. Methoxylated xanthone derivatives were found to be promising PKC activators showing high selectivity for individual PKC isoforms, proving their utility for a detailed study of the physiological and pathophysiological roles of PKC isoforms [317,322,323]. Among the most promising xanthones for activities in which redox mechanisms are involved, it is interesting to mention dihydroxyxanthones, particularly those with a catechol moiety considered PAINS, or pan-assay interference compounds [324]. Although their activity does not depend on a specific, drug-like interaction between the molecule and a protein, dihydroxyxanthones are able to coat a protein or sequester metal ions that are essential to a protein’s function. These mechanisms are recognized for some FDA approved-drugs. Several authors have highlighted the antimicrobial activity of xanthones against diverse human pathogenic microorganisms. The antimicrobial activity of synthetic xanthones, xanthenediones, and spirobenzofurans against the yeasts Cryptococcus neoformans and Candida albicans has been reported [22,325,326]. Hydroxyxanthones have been proposed as novel antimalarial agents, with activity against multidrug-resistant Plasmodium parasites being able to exert complexation to the heme and inhibition of hemozoin formation [327]. Interestingly, 1,3-dihydroxyxanthone derivatives showed the ability to inhibit acetylcholinesterase (AChE) and block the acetylcholinesterase-induced by-amyloid aggregation [70]. The research for new cholinesterase inhibitors is an important strategy to identify new drug candidates to treat Alzheimer’s disease and related dementias. Most currently known natural inhibitors of acetylcholinesterase (AChE) are alkaloids, which have the disadvantages of short half-lives and/or undesirable side effects [328]. A pool of xanthones, such as bellidifolin, bellidin, swertianolin, and norswertianolin from Gentiana campestris Geners. exhibited potent inhibitory activities against AChE with MIC values of 0.01, 0.04, 0.08, and 0.5 μM, respectively [329]. Reutrakul et al. [330,331] demonstrated the anti-HIV-1 activities of the 1,3,8-trihydroxy-2,4-dimethoxyxanthone and euxanthone from Cratoxylum arborescens Blume in the syncytium assay, with EC50 values of 17.9 and 18.8 μM [330]. In addition, morellic acid, gambogic acid, and dihydroisomorellin have shown moderate HIV-1 inhibitory activities in the reverse transcriptase assay, with IC50 values of 11, 15, and 42.3 μM, respectively [331]. Xanthones have been shown to have beneficial effects on several cardiovascular diseases, including atherosclerosis, hypertension, thrombosis and ischemic heart disease [332]. Wang et al. [333] have shown that 1-hydroxy 2,3,5-trimethoxyxanthone, a tetraoxygenated xanthone from Halenia elliptica D.don (Gentianaceae), induces potent concentration-dependent relaxation in rat coronary artery rings pre-contracted with 1 μM of 5- hydroxytryptamine (EC50, 1.67 μM), while one of its major metabolites, 1,5-dihydroxy-2,3-dimethoxyxanthone, induces a relaxation effect with an EC50 of 4.4 μM. It is important to underline that single xanthones may have multiple pharmacological effects, since pharmacophores with diverse effects share the same tricyclic scaffold but differ in the nature and/or positions of substituents. It should be noted that the inventory of natural xanthones remains far from complete, and the functional-group diversity and architectural platforms of natural products generated in their biosynthesis continue to provide new information for synthetic and medicinal chemists in strategies for making biologically active mimics.
6. Conclusions
This review describes the biosynthetic process of xanthone in plants, fungi, and lichens which has yet to be updated comprehensively in the last decade. In higher plants, xanthone biosynthesis involves the shikimate and the acetate pathways which originate in plastids and endoplasmic reticulum, respectively. The pathway continues following three alternative routes, two phenylalanine-dependent and one phenylalanine-independent. However, all three routes lead to the biosynthesis of 2,3′,4,6-tetrahydroxybenzophenone, which is the central intermediate of xanthone biosynthesis. Unlike plants, the xanthone core in fungi and lichens is wholly derived from polyketide. Several xanthone derivatives can be originated from these precursors and differ between plants, fungi, and lichens. Despite there being several studies on chemical and biochemical synthesis of xanthones in plants, there has been little investigation on their subcellular, cellular, and tissue trafficking. As it was reported in this review, these issues have been deeply explored only in few species, including Hypericum spp. and a few others. No study has been reported about these aspects in fungi and lichens. Xanthones are molecules involved in defense response to both biotic and abiotic agents in plants, although their role in fungi and lichens has not yet been exhaustively explored. Interestingly, xanthones derived from plants, fungi, and lichens show biological activities in many human diseases. In this context, further knowledge of mechanisms underlying xanthone biosynthesis in different plant organisms will be useful to optimize the production of these high-value products for application purposes.
Author Contributions
C.B. and G.P. conceived the review; C.B., V.P., E.B. and G.P. wrote the review; G.P. edited and supervised the manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by Sapienza University of Rome Funds for Large Projects 2019–2022, protocol number: RG11916B427D180B.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Schmid, W. Ueber das Mangostin. Justus Liebigs Ann. Chem. 1855, 93, 83–88. [Google Scholar] [CrossRef]
- Le Pogam, P.; Boustie, J. Xanthones of Lichen Source: A 2016 Update. Molecules 2016, 21, 294. [Google Scholar] [CrossRef] [PubMed]
- Masters, K.S.; Bräse, S. Xanthones from fungi, lichens, and bacteria: The natural products and their synthesis. Chem. Rev. 2012, 112, 3717–3776. [Google Scholar] [CrossRef] [PubMed]
- Soares, J.X.; Loureiro, D.R.P.; Dias, A.L.; Reis, S.; Pinto, M.M.M.; Afonso, C.M.M. Bioactive Marine Xanthones: A Review. Mar. Drugs 2022, 20, 58. [Google Scholar] [CrossRef] [PubMed]
- Oldenburg, T.B.; Wilkes, H.; Horsfield, B.; Van Duin, A.C.; Stoddart, D.; Wilhelms, A. Xanthones—Novel aromatic oxygen-containing compounds in crude oils. Org. Geochem. 2002, 33, 595–609. [Google Scholar] [CrossRef]
- Henry, L.; Caventou, J.B. From root of Gentiana lutea L., Gentianaceae: Henry, Caventou. J. Pharm. Chim. 1821, 7, 178. [Google Scholar]
- Kamal, A.; Husain, S.A.; Noorani, R.; Murtaza, N.; Qureshi, I.H.; Qureshi, A.A. Studies in the biochemistry of microorganisms. XI. Isolation of tajixanthone, shamixanthone, ajamxanthone, shahenxanthone, najamxanthone, radixanthone and mannitol from mycelium of Aspergillus stellatus, Curzi. Pak. J. Sci. Indus. Res. 1970, 13, 251–255. [Google Scholar]
- Rawat, M.M.; Pant, G.; Renu, P. Xanthones from Swertia alternifolia. Indian J. Pharm. Sci. 2001, 63, 516. [Google Scholar]
- Zailan, A.A.D.; Karunakaran, T.; Abu Bakar, M.H.; Mian, V.J.Y. The Malaysian genus Calophyllum (Calophyllaceae): A review on its phytochemistry and pharmacological activities. Nat. Prod. Res. 2021, 36, 4569–4579. [Google Scholar] [CrossRef]
- Remali, J.; Sahidin, I.; Aizat, W.M. Xanthone Biosynthetic Pathway in Plants: A Review. Front. Plant Sci. 2022, 916, 809497. [Google Scholar] [CrossRef]
- Chen, L.G.; Yang, L.L.; Wang, C.C. Anti-inflammatory activity of mangostins from Garcinia mangostana. Food Chem. Toxicol. 2008, 46, 688–693. [Google Scholar] [CrossRef]
- Panda, S.S.; Chand, M.; Sakhuja, R.; Jain, S.C. Xanthones as potential antioxidants. Curr. Med. Chem. 2013, 20, 4481–4507. [Google Scholar] [CrossRef]
- Tocci, N.; Simonetti, G.; D’Auria, F.D.; Panella, S.; Palamara, A.T.; Ferrari, F.; Pasqua, G. Chemical composition and antifungal activity of Hypericum perforatum subsp. angustifolium roots from wild plants and plants grown under controlled conditions. Plant Biosyst. 2013, 147, 557–562. [Google Scholar] [CrossRef]
- Wölfle, U.; Seelinger, G.; Schempp, C.M. Topical application of St. John’s wort (Hypericum perforatum). Planta Med. 2014, 80, 109–120. [Google Scholar] [CrossRef]
- Zubrická, D.; Mišianiková, A.; Henzelyová, J.; Valletta, A.; De Angelis, G.; D’Auria, F.D.; Simonetti, G.; Pasqua, G.; Čellárová, E. Xanthones from roots, hairy roots and cell suspension cultures of selected Hypericum species and their antifungal activity against Candida albicans. Plant Cell Rep. 2015, 34, 1953–1962. [Google Scholar] [CrossRef]
- Ahmad, I. Recent insight into the biological activities of synthetic xanthone derivatives. Eur. J. Med. Chem. 2016, 116, 267–280. [Google Scholar]
- Klein-Júnior, L.C.; Campos, A.; Niero, R.; Corrêa, R.; Vander Heyden, Y.; Filho, V.C. Xanthones and cancer: From natural sources to mechanisms of action. Chem. Biodivers. 2020, 17, e1900499. [Google Scholar] [CrossRef]
- Badiali, C.; De Angelis, G.; Simonetti, G.; Brasili, E.; Tobaruela, E.D.C.; Purgatto, E.; Yin, H.; Valletta, A.; Pasqua, G. Chitosan oligosaccharides affect xanthone and VOC biosynthesis in Hypericum perforatum root cultures and enhance the antifungal activity of root extracts. Plant Cell Rep. 2018, 37, 1471–1484. [Google Scholar] [CrossRef]
- Wang, K.; Yu, L.; Shi, J.; Liu, W.; Sang, Z. Multifunctional indanone–chalcone hybrid compounds with anti-β-amyloid (Aβ) aggregation, monoamine oxidase B (MAO-B) inhibition and neuroprotective properties against Alzheimer’s disease. Med. Chem. Res. 2019, 28, 1912–1922. [Google Scholar] [CrossRef]
- Dias, A.C.; Seabra, R.M.; Andrade, P.B.; Ferreres, F.; Ferreira, M.F. Xanthone production in calli and suspended cells of Hypericum perforatum. J. Plant Physiol. 2001, 158, 821–827. [Google Scholar] [CrossRef]
- Tocci, N.; Ferrari, F.; Santamaria, A.R.; Valletta, A.; Rovardi, I.; Pasqua, G. Chitosan enhances xanthone production in Hypericum perforatum subsp. angustifolium cell cultures. Nat. Prod. Res. 2010, 24, 286–293. [Google Scholar] [CrossRef] [PubMed]
- Tocci, N.; Simonetti, G.; D’Auria, F.D.; Panella, S.; Palamara, A.T.; Valletta, A.; Pasqua, G. Root cultures of Hypericum perforatum subsp. angustifolium elicited with chitosan and production of xanthone-rich extracts with antifungal activity. Appl. Microbiol. Biotechnol. 2011, 91, 977–987. [Google Scholar] [CrossRef] [PubMed]
- Tusevski, O.; Stanoeva, J.P.; Stefova, M.; Kungulovski, D.; Pancevska, N.A.; Sekulovski, N.; Panov, S.; Simic, S.G. Hairy roots of Hypericum perforatum L.: A promising system for xanthone production. Cent. Eur. J. Biol. 2013, 8, 1010–1022. [Google Scholar] [CrossRef]
- Brasili, E.; Pratico, G.; Marini, F.; Valletta, A.; Capuani, G.; Sciubba, F.; Miccheli, A.; Pasqua, G. A non-targeted metabolomics approach to evaluate the effects of biomass growth and chitosan elicitation on primary and secondary metabolism of Hypericum perforatum in vitro roots. Metabolomics 2014, 10, 1186–1196. [Google Scholar] [CrossRef]
- Simonetti, G.; Tocci, N.; Valletta, A.; Brasili, E.; D’Auria, F.D.; Idoux, A.; Pasqua, G. In vitro antifungal activity of extracts obtained from Hypericum perforatum adventitious roots cultured in a mist bioreactor against planktonic cells and biofilm of Malassezia furfur. Nat. Prod. Res. 2016, 30, 544–550. [Google Scholar] [CrossRef]
- Valletta, A.; De Angelis, G.; Badiali, C.; Brasili, E.; Miccheli, A.; Di Cocco, M.E.; Pasqua, G. Acetic acid acts as an elicitor exerting a chitosan-like effect on xanthone biosynthesis in Hypericum perforatum L. root cultures. Plant Cell Rep. 2016, 35, 1009–1020. [Google Scholar] [CrossRef]
- Gaid, M.; Singh, P.; El-Awaad, I.; Nagia, M.; Beerhues, L. Biotechnological production of prenylated xanthones for pharmaceutical use. In Pharmaceutical Biocatalysis; Jenny Stanford Publishing: Dubai, United Arab Emirates, 2019; pp. 103–142. [Google Scholar]
- Kaur, P.; Gupta, R.C.; Dey, A.; Malik, T.; Pandey, D.K. Optimization of salicylic acid and chitosan treatment for bitter secoiridoid and xanthone glycosides production in shoot cultures of Swertia paniculata using response surface methodology and artificial neural network. BMC Plant Biol. 2020, 20, 225. [Google Scholar] [CrossRef]
- El-Seedi, H.R.; El-Ghorab, D.M.; El-Barbary, M.A.; Zayed, M.F.; Goransson, U.; Larsson, S.; Verpoorte, R. Naturally occurring xanthones; latest investigations: Isolation, structure elucidation and chemosystematic significance. Curr. Med. Chem. 2009, 16, 2581–2626. [Google Scholar] [CrossRef]
- Pinto, M.M.; Palmeira, A.; Fernandes, C.; Resende, D.I.; Sousa, E.; Cidade, H.; Tiritan, M.E.; Correia-da-Silva, M.; Cravo, S. From natural products to new synthetic small molecules: A journey through the world of xanthones. Molecules 2021, 26, 431. [Google Scholar] [CrossRef]
- Mandal, S.; Das, P.C.; Joshi, P.C. Naturally occuring xanthones from terrestrial flora. J. Indian Chem. Soc. 1992, 69, 611–636. [Google Scholar]
- Talamond, P.; Conejero, G.; Verdeil, J.L.; Poëssel, J.L. Isolation of C-glycosyl xanthones from Coffea pseudozanguebariae and their location. Nat. Prod. Commun. 2011, 6, 1934578X1100601223. [Google Scholar] [CrossRef]
- Sasaki, N.; Nemoto, K.; Nishizaki, Y.; Sugimoto, N.; Tasaki, K.; Watanabe, A.; Goto, F.; Higuchi, A.; Morgan, E.; Hikage, T.; et al. Identification and characterization of xanthone biosynthetic genes contributing to the vivid red coloration of red-flowered gentian. Plant J. 2021, 107, 1711–1723. [Google Scholar] [CrossRef]
- Dua, V.K.; Ojha, V.P.; Roy, R.; Joshi, B.C.; Valecha, N.; Devi, C.U.; Bhatnagar, M.C.; Sharma, V.P.; Subbarao, S.K. Anti-malarial activity of some xanthones isolated from the roots of Andrographis paniculata. J. Ethnopharmacol. 2004, 95, 247–251. [Google Scholar] [CrossRef]
- Panneerselvam, C.; Ponarulselvam, S.; Murugan, K. Potential anti-plasmodial activity of synthesized silver nanoparticle using Andrographis paniculata Nees (Acanthaceae). Arch. Appl. Sci. Res. 2011, 3, 208–217. [Google Scholar]
- Schieber, A.; Berardini, N.; Carle, R. Identification of flavonol and xanthone glycosides from mango (Mangifera indica L. Cv.“Tommy Atkins”) peels by high-performance liquid chromatography-electrospray ionization mass spectrometry. J. Agric. Food Chem. 2003, 51, 5006–5011. [Google Scholar] [CrossRef]
- Dineshkumar, B.; Mitra, A.; Manjunatha, M. Studies on the anti-diabetic and hypolipidemic potentials of mangiferin (xanthone glucoside) in streptozotocin-induced type 1 and type 2 diabetic model rats. Int. J. Adv. Pharm. 2010, 1, 75–85. [Google Scholar]
- Chalisa, S.; Thiendanai, S.; Pattara, S. Cholinesterase inhibitory activities of xanthones from Anaxagorea luzonensis A. Gray. J. Med. Plants Res. 2012, 6, 3781–3785. [Google Scholar]
- Cane, H.P.C.A.; Saidi, N.; Mustanir, M.; Darusman, D.; Idroes, R.; Musman, M. Evaluation of antibacterial and antioxidant activities of xanthone isolated from Orophea corymbosa leaf. Rasayan J. Chem. 2020, 13, 2215–2222. [Google Scholar] [CrossRef]
- Fujita, M.; Inoue, T. Biosynthesis of mangiferin in Anemarrhena asphodeloides Bunge. I. The origin of the xanthone nucleus. Chem. Pharm. Bull. 1980, 28, 2476–2481. [Google Scholar] [CrossRef]
- Ji, D.; Huang, Z.Y.; Fei, C.H.; Xue, W.W.; Lu, T.L. Comprehensive profiling and characterization of chemical constituents of rhizome of Anemarrhena asphodeloides Bge. J. Chromatogr. B 2017, 1060, 355–366. [Google Scholar] [CrossRef]
- Hong, Y.F.; Han, G.Y.; Guo, X.M. Isolation and structure determination of xanthone glycosides of Anemarrhena asphodeloides. Acta Pharm. Sin. 1997, 32, 473–475. [Google Scholar]
- Mulholland, D.A.; Koorbanally, C.; Crouch, N.R.; Sandor, P. Xanthones from Drimiopsis maculata. J. Nat. Prod. 2004, 67, 1726–1728. [Google Scholar] [CrossRef] [PubMed]
- Mutanyatta, J.; Matapa, B.G.; Shushu, D.D.; Abegaz, B.M. Homoisoflavonoids and xanthones from the tubers of wild and in vitro regenerated Ledebouria graminifolia and cytotoxic activities of some of the homoisoflavonoids. Phytochemistry 2003, 62, 797–804. [Google Scholar] [CrossRef] [PubMed]
- Catalano, S.; Luschi, S.; Flamini, G.; Luigi, P.; Neri, E.M.; Morelli, I. A xanthone from Senecio mikanioides leaves. Phytochemistry 1996, 42, 1605. [Google Scholar] [CrossRef]
- Pauletti, P.M.; Castro-Gamboa, I.; Siqueira Silva, D.H.; Young, M.C.M.; Tomazela, D.M.; Eberlin, M.N.; da Silva Bolzani, V. New antioxidant C-glucosylxanthones from the stems of Arrabidaea samydoides. J. Nat. Prod. 2003, 66, 1384–1387. [Google Scholar] [CrossRef]
- Faizi, S.; Zikr-ur-Rehman, S.; Naz, A.; Versiani, M.A.; Dar, A.; Naqvi, S. Bioassay-guided studies on Bombax ceiba leaf extract: Isolation of shamimoside, a new antioxidant xanthone C-glucoside. Chem. Nat. Comp. 2012, 48, 774–779. [Google Scholar] [CrossRef]
- Joshi, K.R.; Devkota, H.P.; Yahara, S. Chemical analysis of flowers of Bombax ceiba from Nepal. Nat. Prod. Commun. 2013, 8, 1934578X1300800508. [Google Scholar] [CrossRef]
- Shahat, A.A.; Hassan, R.A.; Nazif, N.M.; Van Miert, S.; Pieters, L.; Hammuda, F.M.; Vlietinck, A.J. Isolation of mangiferin from Bombax malabaricum and structure revision of shamimin. Planta Med. 2003, 69, 1068–1070. [Google Scholar]
- de Oliveira, W.G.; Mesquita, A.A.; Kubitzki, K.; Gottlieb, O.R. Xanthones from Bonnetia dinizii. Phytochemistry 1990, 29, 1893–1894. [Google Scholar] [CrossRef]
- Iinuma, M.; Ito, T.; Tosa, H.; Tanaka, T.; Miyake, R.; Chelladurai, V. Prenylated xanthonoids from Calophyllum apetalum. Phytochemistry 1997, 46, 1423–1429. [Google Scholar] [CrossRef]
- Iinuma, M.; Tosa, H.; Toriyama, N.; Tanaka, T.; Ito, T.; Chelladurai, V. Six xanthones from Calophyllum austroindicum. Phytochemistry 1996, 43, 681–685. [Google Scholar] [CrossRef]
- Somanathan, R.; Sultanbawa, M.U.S. Chemical investigation of ceylonese plants. Part I. Extractives of Calophyllum calaba L. and Calophyllum bracteatum Thw. (Guttiferae). J. Chem. Soc. 1972, 1935–1943. [Google Scholar] [CrossRef]
- Ito, C.; Itoigawa, M.; Mishina, Y.; Filho, V.C.; Mukainaka, T.; Tokuda, H.; Nishino, H.; Furukawa, H. Chemical Constituents of Calophyllum brasiliensis: Structure Elucidation of Seven New Xanthones and Their Cancer Chemopreventive Activity. J. Nat. Prod. 2002, 65, 267–272. [Google Scholar] [CrossRef]
- Ito, C.; Itoigawa, M.; Mishina, Y.; Filho, V.C.; Enjo, F.; Tokuda, H.; Nishino, H.; Furukawa, H. Chemical constituents of Calophyllum brasiliense. 2. Structure of three new coumarins and cancer chemopreventive activity of 4-substituted coumarins. J. Nat. Prod. 2003, 66, 368–371. [Google Scholar] [CrossRef]
- Pretto, J.B.; Cechinel-Filho, V.; Noldin, V.F.; Sartori, M.R.; Isaias, D.E.; Cruz, A.B. Antimicrobial activity of fractions and compounds from Calophyllum brasiliense (Clusiaceae/Guttiferae). Z. Nat. C. 2004, 59, 657–662. [Google Scholar] [CrossRef]
- Isaias, D.E.B.; Niero, R.; Noldin, V.F.; de Campos-Buzzi, F.; Yunes, R.A.; Delle-Monache, F.; Cechinel-Filho, V. Pharmacological and phytochemical investigations of different parts of Calophyllum brasiliense (Clusiaceae). Die Pharmazie-An Int. J. Pharm. 2004, 59, 879–881. [Google Scholar]
- Reyes-Chilpa, R.; Jimenez-Estrada, M.; Estrada-Muñiz, E. Antifungal xanthones from Calophyllum brasiliensis heartwood. J. Chem. Ecol. 1997, 23, 1901–1911. [Google Scholar] [CrossRef]
- King, F.E.; King, T.J.; Manning, L.C. 804. The chemistry of extractives from hardwoods. Part XIV. The constitution of jacareubin, a pyranoxanthone from Calophyllum brasiliense. J. Chem. Soc. 1953, 804, 3932–3937. [Google Scholar] [CrossRef]
- Na Pattalung, P.; Thongtheeraparp, W.; Wiriyachitra, P.; Taylor, W.C. Xanthones of Garcinia cowa. Planta Med. 1994, 60, 365–368. [Google Scholar] [CrossRef]
- Kumar, V.; Sotheeswaran, S.; Surendrakumar, S.; Balasubramaniam, S. Calocalabaxanthone, the putative isoprenyl precursor of calabaxanthone from Calophyllum calaba. Phytochemistry 1982, 21, 807–809. [Google Scholar] [CrossRef]
- Carpenter, I.; Locksley, H.D.; Scheinmann, F. Extractives from Guttiferae. Part X. The isolation and structure of four xanthones from Calophyllum canum Hook f. J. Chem. Soc. Org. 1969, 486–488. [Google Scholar] [CrossRef]
- Morel, C.; Séraphin, D.; Oger, J.M.; Litaudon, M.; Sévenet, T.; Richomme, P.; Bruneton, J. New xanthones from Calophyllum caledonicum. J. Nat. Prod. 2000, 63, 1471–1474. [Google Scholar] [CrossRef] [PubMed]
- Hay, A.E.; Hélesbeux, J.J.; Duval, O.; Labaïed, M.; Grellier, P.; Richomme, P. Antimalarial xanthones from Calophyllum caledonicum and Garcinia vieillardii. Life Sci. 2004, 75, 3077–3085. [Google Scholar] [CrossRef] [PubMed]
- Morel, C.; Séraphin, D.; Teyrouz, A.; Larcher, G.; Bouchara, J.P.; Litaudon, M.; Richomme, P.; Bruneton, J. New and antifungal xanthones from Calophyllum caledonicum. Planta Med. 2002, 68, 41–44. [Google Scholar] [CrossRef] [PubMed]
- Morel, C.; Hay, A.E.; Litaudon, M.; Sévenet, T.; Séraphin, D.; Bruneton, J.; Richomme, P. Thirteen new xanthone derivatives from Calophyllum caledonicum (Clusiaceae). Molecules 2002, 7, 38–50. [Google Scholar] [CrossRef]
- Oger, J.M.; Morel, C.; Helesbeux, J.J.; Litaudon, M.; Séraphin, D.; Dartiguelongue, C.; Larcher, G.; Richomme, P.; Duval, O. First 2-hydroxy-3-methylbut-3-enyl substituted xanthones isolated from plants: Structure elucidation, synthesis and antifungal activity. Nat. Prod. Res. 2003, 17, 195–199. [Google Scholar] [CrossRef]
- Locksley, H.D.; Murray, I.G. Extractives from Guttiferae. Part XII. The isolation and structure of seven xanthones from Calophyllum fragrans Ridley. J. Chem. Soc. Org. 1969, 1567–1571. [Google Scholar] [CrossRef]
- Yimdjo, M.C.; Azebaze, A.G.; Nkengfack, A.E.; Meyer, A.M.; Bodo, B.; Fomum, Z.T. Antimicrobial and cytotoxic agents from Calophyllum inophyllum. Phytochemistry 2004, 65, 2789–2795. [Google Scholar] [CrossRef]
- Qin, J.; Lan, W.; Liu, Z.; Huang, J.; Tang, H.; Wang, H. Synthesis and biological evaluation of 1, 3-dihydroxyxanthone mannich base derivatives as anticholinesterase agents. Chem. Cent. J. 2013, 7, 78. [Google Scholar] [CrossRef]
- Iinuma, M.; Tosa, H.; Tanaka, T.; Ito, T.; Yonemori, S.; Chelladurai, V.; Aquil, M.; Takahashi, Y.; Naganawa, H. Occurrence of xanthonolignoids in Guttifereous plants. Heterocycles 1996, 7, 1521–1527. [Google Scholar] [CrossRef]
- Al-Jeboury, F.S.; Locksley, H.D. Xanthones in the heartwood of Calophyllum inophyllum: A geographical survey. Phytochemistry 1971, 10, 603–606. [Google Scholar] [CrossRef]
- Goh, S.H.; Jantan, I. A xanthone from Calophyllum inophyllum. Phytochemistry 1991, 30, 366–367. [Google Scholar] [CrossRef]
- Mah, S.H.; Ee, G.C.L.; Teh, S.S.; Sukari, M.A. Calophyllum inophyllum and Calophyllum soulattri source of anti-proliferative xanthones and their structure–activity relationships. Nat. Prod. Res. 2015, 29, 98–101. [Google Scholar] [CrossRef]
- Iinuma, M.; Tosa, H.; Tanaka, T.; Yonemori, S. Two xanthones from root bark of Calophyllum inophyllum. Phytochemistry 1994, 35, 527–532. [Google Scholar] [CrossRef]
- Su, X.H.; Zhang, M.L.; Li, L.G.; Huo, C.H.; Gu, Y.C.; Shi, Q.W. Chemical constituents of the plants of the genus Calophyllum. Chem. Biodivers. 2008, 5, 2579. [Google Scholar] [CrossRef]
- Jackson, B.; Locksley, H.D.; Scheinmann, F. The isolation of 6-desoxyjacareubin, 2-(3, 3-dimethylallyl)-1, 3, 5, 6-tetrahydroxyxanthone and jacareubin from Calophyllum inophyllum. Phytochemistry 1969, 8, 927–929. [Google Scholar] [CrossRef]
- Peres, V.; Nagem, T.J. Naturally occurring pentaoxygenated, hexaoxygenated and dimeric xanthones: A literature survey. Quím. Nova 1997, 20, 388–397. [Google Scholar] [CrossRef]
- Dharmaratne, H.R.W.; Wijesinghe, W.N.M. A trioxygenated diprenylated chromenxanthone from Calophyllum moonii. Phytochemistry 1997, 46, 1293–1295. [Google Scholar] [CrossRef]
- Scheinmann, F.; Sripong, N.A. Xanthones from the heartwood of Calophyllum neo-ebudicum: Comments, on the taxonomic value of jacareubin in Calophyllum species. Phytochemistry 1971, 10, 1331–1333. [Google Scholar] [CrossRef]
- Bhanu, S.; Scheinmann, F.; Jefferson, A. Xanthones from the heartwood of Calophyllum ramiflorum. Phytochemistry 1975, 14, 298–299. [Google Scholar] [CrossRef]
- Jackson, B.; Locksley, H.D.; Scheinmann, F. Extractives from Guttiferae. Part I. Extractives of Calophyllum sclerophyllum Vesq. J. Chem. Soc. C 1966, 178–181. [Google Scholar] [CrossRef]
- Jackson, B.; Locksley, H.D.; Scheinmann, F. Extractives from Guttiferae. Part VII. The isolation and structure of seven xanthones from Calophyllum scriblitifolium Henderson and Wyatt-Smith. J. Chem. Soc. C 1967, 2500–2507. [Google Scholar] [CrossRef]
- Tjahjandarie, T.S.; Saputri, R.D.; Tanjung, M. 5, 9, 11-Trihydroxy-2, 2-dimethyl-3-(2-methylbut-3-en-2-yl) pyrano [2, 3-a] xanthen-12 (2H)-one from the Stem Bark of Calophyllum tetrapterum Miq. Molbank 2017, 2017, M936. [Google Scholar] [CrossRef]
- Banerji, A.; Deshpande, A.D.; Prabhu, B.R.; Pradhan, P. Tomentonone, a new xanthonoid from the stem bark of Calophyllum tomentosum. J. Nat. Prod. 1994, 57, 396–399. [Google Scholar] [CrossRef]
- Karunanayake, S.; Sotheeswaran, S.; Uvais, M.; Sultanbawa, S.; Balasubramaniam, S. Xanthones and triterpenes of Calophyllum tomentosum. Phytochemistry 1981, 20, 1303–1304. [Google Scholar] [CrossRef]
- Gunasekera, S.P.; Sotheeswaran, S.; Sultanbawa, M.U.S. Two new xanthones, calozeyloxanthone and zeyloxanthonone, from Calophyllum zeylanicum (Guttiferae). J. Chem. Soc. 1981, 1831–1835. [Google Scholar] [CrossRef]
- Castelão Jr, J.F.; Gottlieb, O.R.; De Lima, R.A.; Mesquita, A.A.; Gottliebb, H.E.; Wenkert, E. Xanthonolignoids from Kielmeyera and Caraipa species—13C NMR spectroscopy of xanthones. Phytochemistry 1977, 16, 735–740. [Google Scholar] [CrossRef]
- De Lima, R.A.; Gottlieb, O.R.; Mesquita, A.L. Xanthones from Caraipa densiflora. Phytochemistry 1972, 11, 2307–2309. [Google Scholar] [CrossRef]
- Nagem, T.J.; Da Silveira, J.C. Leiaxanthone, a 1,3,5,6-tetraoxygenated xanthone from Haploclathra leiantha. Phytochemistry 1986, 25, 503–504. [Google Scholar] [CrossRef]
- Nagem, T.J. Anthaxanthone, A 1,3,7,8-tetraoxygenated xanthone from Haploclathra leiantha. Phytochemistry 1988, 27, 646–647. [Google Scholar] [CrossRef]
- Nagem, T.J.; Da Silveira, J.C. A 1, 3, 7, 8-tetraoxygenated xanthone from Haploclathra paniculata. Phytochemistry 1986, 25, 2681–2682. [Google Scholar] [CrossRef]
- Nagem, T.J.; Da Silveira, J.C. Haploxanthone from Haploclathra species. Phytochemistry 1989, 28, 2211–2212. [Google Scholar] [CrossRef]
- Gottlieb, O.R.; Mesquita, A.L.; da Silva, E.M.; de Melo, M.T. Xanthones of Kielmeyera ferruginea. Phytochemistry 1969, 8, 665–666. [Google Scholar] [CrossRef]
- Duarte, A.P.; Corrêa, D.D.B.; Silva, L.G.F.; Janot, S.; Gottlieb, O.R. A química das Gutíferas brasileiras. XVI–Kielmeyera rupestris n. sp. An. Acad. Bras. Ciênc. 1968, 40, 307–311. [Google Scholar]
- Correa, D.; Silva, L.G. The chemistry of Brazilian Guttiferae XVIII. Quinone and xanthone constituents of Kielmeyera rupestris wood. Phytochemistry 1970, 9, 447–451. [Google Scholar] [CrossRef]
- de Barros Corrêa, D.; Fonseca, L.G.; Gottlieb, O.R.; Gonçalves, S.J. Quinone and xanthone constituents of Kielmeyera rupestris. Phytochemistry 1970, 9, 447–451. [Google Scholar] [CrossRef]
- Gottlieb, O.R.; Mesquita, A.L.; De Oliveira, G.G.; De Melo, M.T. Xanthones from Kielmeyera speciosa. Phytochemistry 1970, 9, 2537–2544. [Google Scholar] [CrossRef]
- Pinheiro, L.; Nakamura, C.V.; Dias Filho, B.P.; Ferreira, A.G.; Young, M.C.M.; Cortez, A.G. Antibacterial xanthones from Kielmeyera variabilis mart. (Clusiaceae). Mem. Inst. Oswaldo Cruz 2003, 98, 549–552. [Google Scholar] [CrossRef]
- Walia, S.; Mukerjee, S.K. Ferrxanthone, a 1,3,5,6-tetraoxygenated xanthone from Mesua ferrea. Phytochemistry 1984, 23, 1816–1817. [Google Scholar] [CrossRef]
- Noungoué Tchamo, D.; Cartier, G.; Dijoux-Franca, M.G.; Tsamo, E.; Mariotte, A.M. Xanthones and other constituents of Trema orientalis. Pharm. Biol. 2001, 39, 202–205. [Google Scholar] [CrossRef]
- Kazmi, S.N.U.H.; Ahmed, Z.; Malik, A. Vaccaxanthone, a novel xanthone acid from Saponaria vaccaria. Heterocycles 1989, 29, 1923–1927. [Google Scholar]
- Kazmi, S.N.U.H.; Ahmed, Z.; Malik, A. Sapxanthone, a pentasubstituted xanthone from Saponaria vaccaria. Phytochemistry 1989, 28, 3572–3574. [Google Scholar] [CrossRef]
- Yoshikawa, M.; Nishida, N.; Shimoda, H.; Takada, M.; Kawahara, Y.; Matsuda, H. Polyphenol constituents from Salacia species: Quantitative analysis of mangiferin with alpha-glucosidase and aldose reductase inhibitory activities. Yakugaku zasshi: J. Pharm. Soc. Jpn. 2001, 121, 371–378. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nkengfack, A.E.; Azebaze, G.A.; Vardamides, J.C.; Fomum, Z.T.; van Heerden, F.R. A prenylated xanthone from Allanblackia floribunda. Phytochemistry 2002, 60, 381–384. [Google Scholar] [CrossRef] [PubMed]
- Locksley, H.D.; Murray, I.G. Extractives from Guttiferae. Part XIX. The isolation and structure of two benzophenones, six xanthones and two biflavonoids from the heartwood of Allanblackia floribunda Oliver. J. Chem. Soc. Org. 1971, 1332–1340. [Google Scholar] [CrossRef]
- Azebaze, A.G.; Meyer, M.; Bodo, B.; Nkengfack, A.E. Allanxanthone B, a polyisoprenylated xanthone from the stem bark of Allanblackia monticola Staner LC. Phytochemistry 2004, 65, 2561–2564. [Google Scholar] [CrossRef]
- Azebaze, A.G.B.; Menasria, F.; Noumi, L.G.; Nguemfo, E.L.; Tchamfo, M.F.; Nkengfack, A.E.; Kolb, J.P.; Meyer, M. Xanthones from the Seeds of Allanblackia monticola and Their Apoptotic and Antiproliferative Activities. Planta Med. 2009, 75, 243–248. [Google Scholar] [CrossRef]
- Likhitwitayawuid, K.; Phadungcharoen, T.; Mahidol, C.; Ruchirawat, S. 7-O-Methylgarcinone E from Garcinia cowa. Phytochemistry 1997, 45, 1299–1301. [Google Scholar] [CrossRef]
- Lee, H.H.; Chan, H.K. 1, 3, 6-Trihydroxy-7-methoxy-8-(3, 7-dimethyl-2, 6-octadienyl) xanthone from Garcinia cowa. Phytochemistry 1977, 16, 2038–2040. [Google Scholar] [CrossRef]
- Bandaranayake, W.M.; Selliah, S.S.; Sultanbawa, M.U.S.; Ollis, W.D. Biflavonoids and xanthones of Garcinia terpnophylla and G. echinocarpa. Phytochemistry 1975, 14, 1878–1880. [Google Scholar] [CrossRef]
- Harrison, L.J.; Leong, L.S.; Sai, G.L.; Sim, K.Y.; Tan, H.T.W. Xanthones from Garcinia forbesii. Phytochemistry 1993, 33, 727–728. [Google Scholar] [CrossRef]
- Huang, Y.L.; Chen, C.C.; Chen, Y.J.; Huang, R.L.; Shieh, B.J. Three xanthones and a benzophenone from Garcinia mangostana. J. Nat Prod. 2001, 64, 903–906. [Google Scholar] [CrossRef]
- Nilar, H.L.; Harrison, L.J. Xanthones from the heartwood of Garcinia mangostana. Phytochemistry 2002, 60, 541–548. [Google Scholar] [CrossRef]
- Fu, M.; Qiu, S.X.; Xu, Y.; Wu, J.; Chen, Y.; Yu, Y.; Xiao, G. A new xanthone from the pericarp of Garcinia mangostana. Nat. Prod. Commun. 2013, 8, 1934578X1300801219. [Google Scholar] [CrossRef]
- Suksamrarn, S.; Suwannapoch, N.; Ratananukul, P.; Aroonlerk, N.; Suksamrarn, A. Xanthones from the Green Fruit Hulls of Garcinia mangostana. J. Nat. Prod. 2002, 65, 761–763. [Google Scholar] [CrossRef]
- Kurniawan, Y.S.; Priyangga, K.T.A.; Pranowo, H.D.; Sholikhah, E.N.; Zulkarnain, A.K.; Fatimi, H.A.; Julianus, J. An update on the anticancer activity of xanthone derivatives: A review. Pharmaceuticals 2021, 14, 1144. [Google Scholar] [CrossRef]
- Asai, F.; Tosa, H.; Tanaka, T.; Linuma, M.A. Xanthone from pericarps of Garcinia mangostana. Phytochemistry 1995, 39, 943–944. [Google Scholar] [CrossRef]
- Balasubramanian, K.; Rajagopalan, K. Novel xanthones from Garcinia mangostana, structures of BR-xanthone-A and BR-xanthone-B. Phytochemistry 1988, 27, 1552–1554. [Google Scholar] [CrossRef]
- Govindachari, T.R.; Kalyanaraman, P.S.; Muthukumaraswamy, N.; Pai, B.R. Xanthones of Garcinia mangostana Linn. Tetrahedron 1971, 27, 3919–3926. [Google Scholar] [CrossRef]
- Holloway, D.M.; Scheinmann, F. Phenolic compounds from the heartwood of Garcinia mangostana. Phytochemistry 1975, 14, 2517–2518. [Google Scholar] [CrossRef]
- Parveen, M.; Khan, N.U.D. Two xanthones from Garcinia mangostana. Phytochemistry 1988, 27, 3694–3696. [Google Scholar] [CrossRef]
- Sen, A.K.; Sarkar, K.K.; Mazumder, P.C.; Banerji, N.; Uusvuori, R.; Haset, T.A. A xanthone from Garcinia mangostana. Phytochemistry 1980, 19, 2223–2225. [Google Scholar] [CrossRef]
- Kuete, V.; Sandjo, L.P.; Ouete, J.L.N.; Fouotsa, H.; Wiench, B.; Efferth, T. Cytotoxicity and modes of action of three naturally occurring xanthones (8-hydroxycudraxanthone G, morusignin I and cudraxanthone I) against sensitive and multidrug-resistant cancer cell lines. Phytomedicine 2014, 21, 315–322. [Google Scholar] [CrossRef] [PubMed]
- Goh, S.H.; Jantan, I.; Gray, A.I.; Waterman, P.G. Prenylated xanthones from Garcinia opaca. Phytochemistry 1992, 31, 1383–1386. [Google Scholar] [CrossRef]
- Waterman, P.G.; Crichton, E.G. Xanthones, benzophenones and triterpenes from the stem bark of Garcinia ovalifolia. Planta Med. 1980, 40, 351–355. [Google Scholar] [CrossRef]
- Rao, A.R.; Sarma, M.R.; Venkataraman, K.; Yemul, S.S. A benzophenone and xanthone with unusual hydroxylation patterns from the heartwood of Garcinia pedunculata. Phytochemistry 1974, 13, 1241–1244. [Google Scholar] [CrossRef]
- Waterman, P.G.; Hussain, R.A. Major xanthones from Garcinia quadrifaria and Garcinia staudtii stem barks. Phytochemistry 1982, 21, 2099–2101. [Google Scholar] [CrossRef]
- Hay, A.E.; Aumond, M.C.; Mallet, S.; Dumontet, V.; Litaudon, M.; Rondeau, D.; Richomme, P. Antioxidant xanthones from G. vieillardii. J. Nat Prod. 2004, 67, 707–709. [Google Scholar] [CrossRef]
- Nguyen, C.N.; Trinh, B.T.; Tran, T.B.; Nguyen, L.T.T.; Jäger, A.K.; Nguyen, L.H.D. Anti-diabetic xanthones from the bark of Garcinia xanthochymus. Bioorg. Med. Chem. Lett. 2017, 27, 3301–3304. [Google Scholar] [CrossRef]
- Wabo, H.K.; Kikuchi, H.; Katou, Y.; Tane, P.; Oshima, Y. Xanthones and a benzophenone from the roots of Pentadesma butyracea and their antiproliferative activity. Phytochem. Lett. 2010, 3, 104–107. [Google Scholar] [CrossRef]
- Lenta, B.N.; Ngouela, S.; Noungoue, D.T.; Tsamo, E.; Connolly, J.D. Symphonin: A new prenylated pyranoxanthone with antimicrobial activity from the seeds of Symphonia globulifera (Guttiferae). Bull. Chem. Soc. Ethiop. 2004, 18, 175–180. [Google Scholar] [CrossRef]
- Nkengfack, A.E.; Mkounga, P.; Meyer, M.; Fomum, Z.T.; Bodo, B. Globulixanthones C, D and E: Three prenylated xanthones with antimicrobial properties from the root bark of Symphonia globulifera. Phytochemistry 2002, 61, 181–187. [Google Scholar] [CrossRef]
- Santos, L.C.; Piacente, S.; De Riccardis, F.; Eletto, A.M.; Pizza, C.; Vilegas, W. Xanthones and flavonoids from Leiothrix curvifolia and Leiothrix flavescens. Phytochemistry 2001, 56, 853–856. [Google Scholar] [CrossRef]
- Kapingu, M.C.; Magadula, J.J. Prenylated xanthones and a benzophenone from Baphia kirkii. Nat. Prod. Commun. 2008, 3, 1934578X0800300921. [Google Scholar] [CrossRef]
- Joubert, E.; Otto, F.; Grüner, S.; Weinreich, B. Reversed-phase HPLC determination of mangiferin, isomangiferin and hesperidin in Cyclopia and the effect of harvesting date on the phenolic composition of C. genistoides. Eur. Food Res. Technol. 2003, 216, 270–273. [Google Scholar] [CrossRef]
- Janković, T.; Krstić, D.; Šavikin-Fodulović, K.; Menković, N.; Grubišić, D. Xanthones and secoiridoids from hairy root cultures of Centaurium erythraea and C. pulchellum. Planta Med. 2002, 68, 944–946. [Google Scholar] [CrossRef]
- Valentão, P.; Andrade, P.B.; Silva, E.; Vicente, A.; Santos, H.; Bastos, M.L.; Seabra, R.M. Methoxylated xanthones in the quality control of small centaury (Centaurium erythraea) flowering tops. J. Agric. Food Chem. 2002, 50, 460–463. [Google Scholar] [CrossRef]
- Valentão, P.; Areias, F.; Amaral, J.; Andrade, P.; Seabra, R. Tetraoxygenated xanthones from Centaurium erythraea. Nat. Prod. Lett. 2000, 14, 319–323. [Google Scholar] [CrossRef]
- Versluys, C.; Cortés, M.; López, J.T.; Sierra, J.R.; Razmilić, I. A novel xanthone as secondary metabolite from Centaurium cachanlahuen. Experientia 1982, 38, 771–772. [Google Scholar] [CrossRef]
- Parra, M.; Seoane, E.; Tortajada, A. Additional new xanthones isolated from Centaurium linarifolium. J. Nat. Prod. 1984, 47, 868–871. [Google Scholar] [CrossRef]
- Stout, G.H.; Balkenhol, W.J. Xanthones of the Gentianaceae-I Frasera caroliniensis Walt. Tetrahedron 1969, 25, 1947–1960. [Google Scholar] [CrossRef]
- Plouvier, V.; Massicot, J.; Rivaille, P. On gentiacauleine, a new tetra-substituted xanthone, aglycone of gentiacauloside of Gentiana acaulis L. Comptes Rendus Hebd. Seances L’academie Sci. Ser. D Sci. Nat. 1967, 264, 1219–1222. [Google Scholar]
- Menković, N.; Šavikin-Fodulović, K.; Savin, K. Chemical composition and seasonal variations in the amount of secondary compounds in Gentiana lutea leaves and flowers. Planta Med. 2000, 66, 178–180. [Google Scholar] [CrossRef] [PubMed]
- Gupta, P.; Lewis, J.R. Biogenesis of xanthones in Gentiana lutea. J. Chem. Soc. C 1971, 629–631. [Google Scholar] [CrossRef]
- Mudrić, J.; Janković, T.; Šavikin, K.; Bigović, D.; Đukić-Ćosić, D.; Ibrić, S.; Đuriš, J. Optimization and modelling of gentiopicroside, isogentisin and total phenolics extraction from Gentiana lutea L. roots. Ind. Crops Prod. 2020, 155, 112767. [Google Scholar] [CrossRef]
- Chen, Y.; Wang, G.K.; Wu, C.; Qin, M.J. Chemical constituents of Gentiana rhodantha. China J. Chin. Mater. Med. 2013, 38, 362–365. [Google Scholar]
- Zhang, L.; Zou, D.Z.; Bai, S.; Li, Z.H.; Zhang, C.H.; Li, M.H. Chemical constituents from Gentianella turkestanorum (Gentianaceae). Biochem. Syst. Ecol. 2016, 65, 89–92. [Google Scholar] [CrossRef]
- Moon, U.R.; Mitra, A. A mechanistic insight into hydrogen peroxide-mediated elicitation of bioactive xanthones in Hoppea fastigiata shoot cultures. Planta 2016, 244, 259–274. [Google Scholar] [CrossRef]
- Banerjee, S.; Sur, T.K.; Mandal, S.; Das, P.C.; Sikdar, S. Assessment of the anti-inflammatory effects of Swertia chirata in acute and chronic experimental models in male albino rats. Indian J. Pharmacol. 2000, 32, 21–24. [Google Scholar]
- Wang, C.Z.; Maier, U.H.; Keil, M.; Zenk, M.H.; Bacher, A.; Rohdich, F.; Eisenreich, W. Phenylalanine-independent biosynthesis of 1,3,5,8-tetrahydroxyxanthone: A retrobiosynthetic NMR study with root cultures of Swertia chirata. Eur. J. Biochem. 2003, 270, 2950–2958. [Google Scholar] [CrossRef]
- Zhou, Y.; Di, Y.T.; Gesang, S.; Peng, S.L.; Ding, L.S. Secoiridoid glycosides from Swertia mileensis. Helv. Chim. Acta 2006, 89, 94–102. [Google Scholar] [CrossRef]
- Tan, P.; Hou, C.Y.; Liu, Y.L.; Lin, L.J.; Cordell, G.A. Swertipunicoside. The First Bisxanthone C-Glycoside. J. Org. Chem. 1991, 56, 7130–7133. [Google Scholar] [CrossRef]
- Huang, Q.; Wang, Y.; Wu, H.; Yuan, M.; Zheng, C.; Xu, H. Xanthone glucosides: Isolation, bioactivity and synthesis. Molecules 2021, 26, 5575. [Google Scholar] [CrossRef]
- Ghosal, S.; Sharma, P.V.; Chaudhuri, R.K. Chemical constituents of Gentianaceae X: Xanthone-O-glucosides of Swertia purpurascens Wall. J. Pharm. Sci. 1974, 63, 1286–1290. [Google Scholar] [CrossRef]
- Tomimori, T.; Komatsu, M. Studies on the Constituents of Swertia japonica. VI. on the Flavonoid and Xanthone Constituents of Swertia randaiensis. HAYATA and S. swertopsis MAKINO. Yakugaku Zasshi 1969, 89, 1276–1282. [Google Scholar] [CrossRef]
- Zeng, G.Y.; Tan, G.S.; Xu, K.P.; Xu, X.P.; Li, F.S.; Tan, J.B.; Hu, G.Y. Water-soluble chemical constituents of Swertia davidii Franch. Acta Pharm. Sin. 2004, 39, 351–353. [Google Scholar]
- Mahabusarakam, W.; Nuangnaowarat, W.; Taylor, W.C. Xanthone derivatives from Cratoxylum cochinchinense roots. Phytochemistry 2006, 67, 470–474. [Google Scholar] [CrossRef]
- Iinuma, M.; Tosa, H.; Ito, T.; Tanaka, T.; Madulid, D.A. Two xanthones from roots of Cratoxylum formosanum. Phytochemistry 1996, 42, 1195–1198. [Google Scholar] [CrossRef]
- Boonsri, S.; Karalai, C.; Ponglimanont, C.; Kanjana-Opas, A.; Chantrapromma, K. Antibacterial and cytotoxic xanthones from the roots of Cratoxylum formosum. Phytochemistry 2006, 67, 723–727. [Google Scholar] [CrossRef]
- Kitanov, G.M.; Assenov, I. Flavonols and xanthones from Cratoxylum pruniflorum Kurz. (Guttiferae). Pharmazie 1998, 43, 879–880. [Google Scholar]
- Nielsen, H.; Arends, P. Xanthone constituents of Hypericum androsaemum. J. Nat. Prod. 1979, 42, 301–304. [Google Scholar] [CrossRef]
- Cardona, M.L.; Fernández, M.I.; Pedro, J.R.; Seoane, E.; Vidal, R. Additional new xanthones and xanthonolignoids from Hypericum canariensis. J. Nat. Prod. 1986, 49, 95–100. [Google Scholar] [CrossRef]
- Chung, M.I.; Weng, J.R.; Wang, J.P.; Teng, C.M.; Lin, C.N. Antiplatelet and anti-inflammatory constituents and new oxygenated xanthones from Hypericum geminiflorum. Planta Med. 2002, 68, 25–29. [Google Scholar] [CrossRef] [PubMed]
- Ishiguro, K.; Nagata, S.; Oku, H.; Yamaki, M. Bisxanthones from Hypericum japonicum: Inhibitors of PAF-induced hypotension. Planta Med. 2002, 68, 258–261. [Google Scholar] [CrossRef] [PubMed]
- Arends, P. Maculatoxanthone, a new pyranoxanthone prom Hypericum maculatum. Tetrahedron Lett. 1969, 10, 4893–4896. [Google Scholar] [CrossRef]
- Feng, Z.; Lu, X.; Gan, L.; Zhang, Q.; Lin, L. Xanthones, a promising anti-inflammatory scaffold: Structure, activity, and drug likeness analysis. Molecules 2020, 25, 598. [Google Scholar] [CrossRef]
- Ishiguro, K.; Nakajima, M.; Fukumoto, H.; Isoi, K. A xanthone substituted with an irregular monoterpene in cell suspension cultures of Hypericum patulum. Phytochemistry 1995, 39, 903–905. [Google Scholar] [CrossRef]
- Ishiguro, K.; Nakajima, M.; Fukumoto, H.; Isoi, K. Co-occurrence of prenylated xanthones and their cyclization products in cell suspension cultures of Hypericum patulum. Phytochemistry 1995, 38, 867–869. [Google Scholar] [CrossRef]
- Ishiguro, K.; Fukumoto, H.; Suitani, A.; Nakajima, M.; Isoi, K. Prenylated xanthones from cell suspension cultures of Hypericum patulum. Phytochemistry 1996, 42, 435–437. [Google Scholar] [CrossRef]
- Ishiguro, K.; Nagareya, N.; Suitani, A.; Fukumoto, H. A prenylated xanthone from cell suspension cultures of Hypericum patulum. Phytochemistry 1997, 44, 1065–1066. [Google Scholar] [CrossRef]
- Tocci, N.; Gaid, M.; Kaftan, F.; Belkheir, A.K.; Belhadj, I.; Liu, B.; Svatoš, A.; Hänsch, R.; Pasqua, G.; Beerhues, L. Exodermis and endodermis are the sites of xanthone biosynthesis in Hypericum perforatum roots. New Phytol. 2018, 217, 1099–1112. [Google Scholar] [CrossRef]
- Cardona, M.L.; Fernández, I.; Pedro, J.R.; Serrano, A. Xanthones from Hypericum reflexum. Phytochemistry 1990, 29, 3003–3006. [Google Scholar] [CrossRef]
- Tala, M.F.; Tchakam, P.D.; Wabo, H.K.; Talontsi, F.M.; Tane, P.; Kuiate, J.R.; Laatsch, H. Chemical constituents, antimicrobial and cytotoxic activities of Hypericum riparium (Guttiferae). Rec. Nat. Prod. 2013, 7, 65. [Google Scholar]
- Rath, G.; Potterat, O.; Mavi, S.; Hostettmann, K. Xanthones from Hypericum roeperanum. Phytochemistry 1996, 43, 513–520. [Google Scholar] [CrossRef]
- Hong, D.; Yin, F.; Hu, L.H.; Lu, P. Sulfonated xanthones from Hypericum sampsonii. Phytochemistry 2004, 65, 2595–2598. [Google Scholar] [CrossRef]
- Don, M.J.; Huang, Y.J.; Huang, R.L.; Lin, Y.L. New phenolic principles from Hypericum sampsonii. Chem. Pharm. Bull. 2004, 52, 866–869. [Google Scholar] [CrossRef]
- Huang, L.; Wang, H.; Ye, H.; Du, Z.; Zhang, Y.; Beerhues, L.; Liu, B. Differential expression of benzophenone synthase and chalcone synthase in Hypericum sampsonii. Nat. Prod. Commun. 2012, 7, 1615–1618. [Google Scholar] [CrossRef]
- Chen, M.T.; Kuoh, Y.P.; Wang, C.H.; Chen, C.M.; Kuoh, C.S. Additional constituents of Hypericum subalatum. J. Chin. Chem. Soc. 1989, 36, 165–168. [Google Scholar] [CrossRef]
- Maurice, T.; Wache Blandine Marlyse, O.; Jean Robert, N.; Pierre, M.; Victor, K.; Sterner, O.; Augustin Ephrem, N. Antimicrobial prenylated xanthones and anthraquinones from barks and fruits of Psorospermum adamauense (Engl). Nat. Prod. J. 2013, 3, 60–65. [Google Scholar] [CrossRef]
- Delle Monache, F.; Mac-Quhae, M.M.; Delle Monache, G.; Bettolo, G.M.; De Lima, R.A. Xanthones, xanthonolignoids and other constituents of the roots of Vismia guaramirangae. Phytochemistry 1983, 22, 227–232. [Google Scholar] [CrossRef]
- Bukvicki, D.; Novakovic, M.; Ab Ghani, N.; Marin, P.D.; Asakawa, Y. Secondary metabolites from endemic species Iris adriatica Trinajstic ex Mitic (Iridaceae). Nat. Prod. Res. 2018, 32, 1849–1852. [Google Scholar] [CrossRef] [PubMed]
- Abdel-Mageed, W.M.; Al-Wahaibi, L.H.; Al-Saleem, M.S.M.; Gouda, Y.G.; Abdel-Kader, M.S.; Ibraheim, Z.Z. Phytochemical and chemotaxonomic study on Iris albicans Lange leaves. Biochem. Syst. Ecol. 2018, 76, 32–34. [Google Scholar] [CrossRef]
- Fujita, M.; Inoue, T. Studies on the Constituents of Iris florentina L. II. C-Glucosides of Xanthones and Flavones from the Leaves. Chem. Pharm. Bull. 1982, 30, 2342–2348. [Google Scholar] [CrossRef]
- Xie, G.Y.; Chen, Y.J.; Wen, R.; Xu, J.Y.; Wu, S.S.; Qin, M.J. Chemical constituents from rhizomes of Iris germanica. China J. Chin. Mater. Med. 2014, 39, 846–850. [Google Scholar]
- Alkhalil, S.; Tosa, H.; Iinuma, M. A xanthone C-glycoside from Iris nigricans. Phytochemistry 1995, 38, 729–731. [Google Scholar] [CrossRef]
- Wang, D.Y.; Xu, S.Y. Two new xanthones from Premna microphylla. Nat. Prod. Res. 2003, 17, 75–77. [Google Scholar] [CrossRef]
- Pettit, G.R.; Zhang, Q.; Pinilla, V.; Herald, D.L.; Doubek, D.L.; Duke, J.A. Isolation and Structure of Gustastatin from the Brazilian Nut Tree Gustavia hexapetala, 1. J. Nat Prod. 2004, 67, 983–985. [Google Scholar] [CrossRef]
- Fukai, T.; Yonekawa, M.; Hou, A.J.; Nomura, T.; Sun, H.D.; Uno, J. Antifungal agents from the roots of Cudrania cochinchinensis against candida, cryptococcus, and aspergillus species. J. Nat. Prod. 2003, 66, 1118–1120. [Google Scholar] [CrossRef]
- Marques, V.L.L.; De Oliveira, F.M.; Conserva, L.M.; Brito, R.G.L.; Guilhon, G.M.S. Dichromenoxanthones from Tovomita brasiliensis. Phytochemistry 2000, 55, 815–818. [Google Scholar] [CrossRef]
- Pu, X.; Li, J.; Guo, Z.; Wang, M.; Lei, M.; Yang, S.; Huang, Q. Structure-based identification and pathway elucidation of flavonoids in Camptotheca acuminate. Synth. Syst. Biotechnol. 2022, 7, 824–836. [Google Scholar] [CrossRef]
- Fenner, M.; Lee, W.G.; Duncan, S.J. Chemical features of Chionochloa species in relation to grazing by ruminants in South Island, New Zealand. N. Z. J. Ecol. 1993, 17, 35–40. [Google Scholar]
- Lin, L.L.; Huang, F.; Chen, S.B.; Yang, D.J.; Chen, S.L.; Yang, J.S.; Xiao, P.G. Xanthones from the roots of Polygala caudata and their antioxidation and vasodilatation activities in vitro. Planta Med. 2005, 71, 372–375. [Google Scholar] [CrossRef]
- Miyase, T.; Noguchi, H.; Chen, X.M. Sucrose Esters and Xanthone C-Glycosides from the Roots of Polygala sibirica. J. Nat. Prod. 1999, 62, 993–996. [Google Scholar] [CrossRef]
- Jiang, Y.; Zhang, W.; Tu, P.F.; Xu, X.J. Xanthone Glycosides from Polygala tenuifolia and Their Conformational Analyses. J. Nat. Prod. 2005, 68, 875–879. [Google Scholar] [CrossRef]
- Dall’Acqua, S.; Innocenti, G.; Viola, G.; Piovan, A.; Caniato, R.; Cappelletti, E.M. Cytotoxic compounds from Polygala vulgaris. Chem. Pharm. Bull. 2002, 50, 1499–1501. [Google Scholar] [CrossRef]
- Talamond, P.; Mondolot, L.; Gargadennec, A.; de Kochko, A.; Hamon, S.; Fruchier, A.; Campa, C. First report on mangiferin (C-glucosyl-xanthone) isolated from leaves of a wild coffee plant, Coffea pseudozanguebariae (Rubiaceae). Acta Bot. Gall. 2008, 155, 513–519. [Google Scholar] [CrossRef]
- Ferrari, J.; Terreaux, C.; Sahpaz, S.; Msonthi, J.D.; Wolfender, J.L.; Hostettmann, K. Benzophenone glycosides from Gnidia involucrata. Phytochemistry 2000, 54, 883–889. [Google Scholar] [CrossRef]
- Carvalho, M.J.; Carvalho, L.M.; Ferreira, A.M.; Silva, A.M. A new xanthone from Hedychium gardnerianum. Nat. Prod. Res. 2003, 17, 445–449. [Google Scholar] [CrossRef]
- Malet-Cascon, L.; Romero, F.; Espliego-Vazquez, F.; Gravalos, D.; Fernandez-Puentes, J.L. IB-00208, a new cytotoxic polycyclic xanthone produced by a marine-derived Actinomadura I. Isolation of the strain, taxonomy and biological activites. J. Antibiot. 2003, 56, 219–225. [Google Scholar] [CrossRef]
- Ullmann, F.; Panchaud, L. Synthese des Euxanthons. Justus Liebigs Ann. Chem. 1906, 350, 108–117. [Google Scholar] [CrossRef]
- Ignatushchenko, M.V.; Winter, R.W.; Bächinger, H.P.; Hinrichs, D.J.; Riscoe, M.K. Xanthones as antimalarial agents; studies of a possible mode of action. FEBS Lett. 1997, 409, 67–73. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Minto, R.E.; Townsend, C.A. Enzymology and molecular biology of aflatoxin biosynthesis. Chem. Rev. 1997, 97, 2537–2556. [Google Scholar] [CrossRef] [PubMed]
- Pinto, M.M.; Castanheiro, R.A.; Kijjoa, A. Xanthones from Marine-Derived Microorganisms: Isolation, Structure Elucidation and Biological Activities. Encycl. Anal. Chem. Appl. Theory Instrum. 2006, 1–21. [Google Scholar]
- Essery, J.M.; O’Herron, F.A.; McGregor, D.N.; Bradner, W.T. Preparation and antitumor activities of some derivatives of 5-methoxysterigmatocystin. J. Med. Chem. 1976, 19, 1339–1342. [Google Scholar] [CrossRef] [PubMed]
- Kamal, A.; Haider, Y.; Khan, Y.A.; Qureshi, I.H.; Qureshi, A.A. Studies in the biochemistry of microorganisms. X. Isolation, structure and stereochemistry of yasimin and other metabolic products of Aspergillus unguis Emile-Weil and Gaudin. Pakistan J. Sci. Indus. Res. 1970, 13, 251. [Google Scholar]
- Chexal, K.K.; Holker, J.S.; Simpson, T.J. The biosynthesis of fungal metabolites. Part VI. Structures and biosynthesis of some minor metabolites from variant strains of Aspergillus variecolor. J. Chem. Soc. Perkin Trans. 1 1975, 6, 549–554. [Google Scholar] [CrossRef]
- Ishida, M.; Hamasaki, T.; Hatsuda, Y. Biosynthesis of shamixanthone. Agric. Biol. Chem. 1978, 42, 465–466. [Google Scholar]
- Valdomir, G.; Tietze, L.F. Chromanone Lactones: A Neglected Group of Natural Products–Isolation, Structure Elucidation, Bioactivity, and Synthesis. Eur. J. Org. Chem. 2022, 20, e202200201. [Google Scholar] [CrossRef]
- Wu, Z.J.; Ouyang, M.A.; Tan, Q.W. New asperxanthone and asperbiphenyl from the marine fungus Aspergillus sp. Pest Manag. Sci. 2009, 65, 60–65. [Google Scholar] [CrossRef]
- Marks, K.M.; Park, E.S.; Arefolov, A.; Russo, K.; Ishihara, K.; Ring, J.E.; Clardy, J.; Clarke, A.S.; Pelish, H.E. The selectivity of austocystin D arises from cell-line-specific drug activation by cytochrome P450 enzymes. J. Nat. Prod. 2011, 74, 567–573. [Google Scholar] [CrossRef]
- Fredimoses, M.; Zhou, X.; Ai, W.; Tian, X.; Yang, B.; Lin, X.; Liu, J.; Liu, Y. Emerixanthone E, a new xanthone derivative from deep sea fungus Emericella sp SCSIO 05240. Nat. Prod. Res. 2019, 33, 2088–2094. [Google Scholar] [CrossRef]
- Kjaer, D.; Kjaer, A.; Pedersen, C.; Bu’Lock, J.D.; Smith, J.R. Bikaverin and norbikaverin, benzoxanthentrione pigments of Gibberella fujikuroi. J. Chem. Soc. C 1971, 16, 2792–2797. [Google Scholar] [CrossRef]
- Peres, V.; Nagem, T.; Faustino de Oliviera, F.; Peres, V.; Nagem, T.J.; de Oliveira, F.F. Tetraoxygenated naturally occurring xanthones. Phytochemistry 2000, 55, 683–710. [Google Scholar] [CrossRef]
- Gnerre, C.; Thull, U.; Gaillard, P.; Carrupt, P.A.; Testa, B.; Fernandes, E.; Silva, F.; Pinto, M.; Pinto, M.M.M.; Wolfender, J.L.; et al. Natural and synthetic xanthones as monoamine oxidase inhibitors: Biological assay and 3D-QSAR. Helv. Chim. Acta 2001, 84, 552–570. [Google Scholar] [CrossRef]
- Michael, A. On the Action of Aldehydes on Phenols I. Am. Chem. J. 1883, 5, 338–349. [Google Scholar]
- Casillas, L.K.; Townsend, C.A. Total synthesis of O-methylsterigmatocystin using N-alkylnitrilium salts and carbonyl− alkene interconversion in a new xanthone synthesis. J. Org. Chem. 1999, 64, 4050–4059. [Google Scholar] [CrossRef]
- El-Seedi, H.R.; El-Barbary, M.A.; El-Ghorab, D.M.H.; Bohlin, L.; Borg-Karlson, A.K.; Goransson, U.; Verpoorte, R. Recent insights into the biosynthesis and biological activities of natural xanthones. Curr. Med. Chem. 2010, 17, 854–901. [Google Scholar] [CrossRef]
- Xie, F.; Pan, X.; Lin, S.; Hu, Y. A base-promoted desalicyloylative dimerization of 3-(1-alkynyl) chromones: An unusual approach to 2-alkynyl xanthones. Org. Biomol. Chem. 2010, 8, 1378–1381. [Google Scholar] [CrossRef]
- Elix, J.; Ovstedal, D.O. Lichen phytochemistry II: Some species of Calopadia. Australas. Lichenol. 2009, 65, 7–9. [Google Scholar]
- Millot, M.; Tomasi, S.; Studzinska, E.; Rouaud, I.; Boustie, J. Cytotoxic constituents of the lichen Diploicia canescens. J. Nat. Prod. 2009, 72, 2177–2180. [Google Scholar] [CrossRef]
- Leuckert, C.; Ahmadjian, V.; Culberson, C.F.; Johnson, A. Xanthones and depsidones of the lichen Lecanora dispersa in nature and of its mycobiont in culture. Mycologia 1990, 82, 370–378. [Google Scholar] [CrossRef]
- Bertrand, M.; Monnat, J.Y.; Lohézic-Le Dévéhat, F. Myriolecis massei, a new species of Lecanoraceae from the coasts of the Armorican Massif in Western Europe. Bryologist 2018, 121, 253–263. [Google Scholar] [CrossRef]
- Veríssimo, A.C.; Pinto, D.C.; Silva, A.M. Marine-Derived Xanthone from 2010 to 2021: Isolation, Bioactivities and Total Synthesis. Mar. Drugs 2022, 20, 347. [Google Scholar] [CrossRef] [PubMed]
- Gartman, J.A.; Tambar, U.K. Recent total syntheses of anthraquinone-based natural products. Tetrahedron 2021, 105, 132501. [Google Scholar] [CrossRef] [PubMed]
- Perez-Vargas, I.; Hernandez-Padron, C.; Etayo, J. New species of Pertusaria (lichenized Ascomycota: Pertusariaceae) from the Canary Islands. Lichenologist 2010, 42, 35–41. [Google Scholar] [CrossRef]
- Elix, J. Five new species of Phyllopsora (lichenized Ascomycota) from Australia; ANU Research Publications: Australia, 1328; ISSN 1328-4401. [Google Scholar]
- Holker, J.S.; Lapper, R.D.; Simpson, T.J. The biosynthesis of fungal metabolites. Part IV. Tajixanthone: 13 C nuclear magnetic resonance spectrum and feedings with [1-13 C]-and [2-13 C]-acetate. J. Chem. Soc. Perkin Trans. 1 1974, 2135–2140. [Google Scholar] [CrossRef]
- Tanahashi, T.; Takenaka, Y.; Ikuta, Y.; Tani, K.; Nagakura, N.; Hamada, N. Xanthones from the cultured lichen mycobionts of Pyrenula japonica and Pyrenula pseudobufonia. Phytochemistry 1999, 52, 401–405. [Google Scholar] [CrossRef]
- Sérusiaux, E.; Lücking, R.; Lumbsch, T. Sporopodium isidiatum (Pilocarpaceae), new from Papua New Guinea and Sri Lanka, with a key to the world’s Sporopodium species. Mycotaxon 2008, 103, 255–262. [Google Scholar]
- Santesson, J. Chemical studies on lichens. 21. Two novel chlorinated lichen xanthones. J. Ark. Kemi 1969, 31, 121–124. [Google Scholar]
- Vieira, L.M.M.; Kijjoa, A. Naturally-occurring xanthones: Recent developments. Curr. Med. Chem. 2005, 12, 2413–2446. [Google Scholar] [CrossRef]
- Sultanbawa, M.U.S. Xanthonoids of tropical plants. Tetrahedron 1980, 36, 1465–1506. [Google Scholar] [CrossRef]
- Sousa, M.E.; Pinto, M.M.M. Synthesis of Xanthones: An Overview. Curr. Med. Chem. 2005, 12, 2447–2479. [Google Scholar] [CrossRef]
- Ahmad, V.U.; Khana, M.A.; Arfan, M.; Siddiqui, M.T. A xanthone dixylopyranoside from Swertia thomsonii. Z. Naturforsch. B 2002, 57, 122–126. [Google Scholar] [CrossRef]
- Iseda, S. On mangiferin, the coloring matter of mango (Mangifera indica Linn.). IV. Isolation of 1, 3, 6, 7-tetrahydroxyxanthone and the skeletal structure of mangiferin. Bull. Chem. Soc. Jpn. 1957, 30, 625–629. [Google Scholar] [CrossRef]
- Wagenaar, M.M.; Clardy, J. Dicerandrols, new antibiotic and cytotoxic dimers produced by the fungus Phomopsis longicolla Isolated from an endangered mint. J. Nat. Prod. 2001, 64, 1006–1009. [Google Scholar] [CrossRef]
- Kumagai, K.; Hosotani, N.; Kikuchi, K.; Kimura, T.; Saji, I. Xanthofulvin, a novel semaphorin inhibitor produced by a strain of Penicillium. J. Antibiot. 2003, 56, 610–616. [Google Scholar] [CrossRef]
- Resende, D.I.; Durães, F.; Maia, M.; Sousa, E.; Pinto, M.M. Recent advances in the synthesis of xanthones and azaxanthones. Org. Chem. Front. 2020, 7, 3027–3066. [Google Scholar] [CrossRef]
- Leuschner, C.; Schultz, G. Non-light-dependent Shikimate Pathway in Plastids from Pea Roots. Bot. Acta 1991, 104, 240–244. [Google Scholar] [CrossRef]
- De Schepper, V.; De Swaef, T.; Bauweraerts, I.; Steppe, K. Phloem transport: A review of mechanisms and controls. J. Exp. Bot. 2013, 64, 4839–4850. [Google Scholar] [CrossRef]
- Franklin, G.; Conceição, L.F.; Kombrink, E.; Dias, A.C. Xanthone biosynthesis in Hypericum perforatum cells provides antioxidant and antimicrobial protection upon biotic stress. Phytochemistry 2009, 70, 60–68. [Google Scholar] [CrossRef]
- El-Mawla, A.; Abd, M.A.; Schmidt, W.; Beerhues, L. Cinnamic acid is a precursor of benzoic acids in cell cultures of Hypericum androsaemum L. but not in cell cultures of Centaurium erythraea RAFN. Planta 2001, 212, 288–293. [Google Scholar] [CrossRef] [PubMed]
- Bennett, G.J.; Lee, H.H.; Das, N.P. Biosynthesis of mangostin. Part 1. The origin of the xanthone skeleton. J. Chem. Soc. Perkin. Trans. 1990, 1, 2671–2676. [Google Scholar] [CrossRef]
- Beerhues, L.; Berger, U. Differential accumulation of xanthones in methyl-jasmonate-and yeast-extract-treated cell cultures of Centaurium erythraea and Centaurium littorale. Planta 1995, 197, 608–612. [Google Scholar] [CrossRef]
- Beerhues, L. Benzophenone synthase from cultured cells of Centaurium erythraea. FEBS Lett. 1996, 383, 264–266. [Google Scholar] [CrossRef] [PubMed]
- Adams, Z.P.; Ehlting, J.; Edwards, R. The regulatory role of shikimate in plant phenylalanine metabolism. J. Theor. Biol. 2019, 462, 158–170. [Google Scholar] [CrossRef]
- Perez de Souza, L.; Garbowicz, K.; Brotman, Y.; Tohge, T.; Fernie, A.R. The acetate pathway supports flavonoid and lipid biosynthesis in Arabidopsis. Plant Physiol. 2020, 182, 857–869. [Google Scholar] [CrossRef]
- Qian, Y.; Lynch, J.H.; Guo, L.; Rhodes, D.; Morgan, J.A.; Dudareva, N. Completion of the cytosolic post-chorismate phenylalanine biosynthetic pathway in plants. Nat. Commun. 2019, 10, 15. [Google Scholar] [CrossRef]
- Valera, M.J.; Boido, E.; Ramos, J.C.; Manta, E.; Radi, R.; Dellacassa, E.; Carrau, F. The Mandelate pathway, an alternative to the phenylalanine ammonia lyase pathway for the synthesis of benzenoids in ascomycete yeasts. Appl. Environ. Microbiol. 2020, 86, e00701-20. [Google Scholar] [CrossRef]
- Cho, M.H.; Corea, O.R.; Yang, H.; Bedgar, D.L.; Laskar, D.D.; Anterola, A.M.; Moog-Anterola, F.A.; Hood, R.L.; Kohalmi, S.E.; Bernards, M.A.; et al. Phenylalanine biosynthesis in Arabidopsis thaliana: Identification and characterization of arogenate dehydratases. J. Biol. Chem. 2007, 282, 30827–30835. [Google Scholar] [CrossRef]
- Rippert, P.; Puyaubert, J.; Grisollet, D.; Derrier, L.; Matringe, M. Tyrosine and phenylalanine are synthesized within the plastids in Arabidopsis. Plant Physiol. 2009, 149, 1251–1260. [Google Scholar] [CrossRef]
- Yoo, H.; Widhalm, J.R.; Qian, Y.; Maeda, H.; Cooper, B.R.; Jannasch, A.S.; Gonda, I.; Lewinsohn, E.; Rhodes, D.; Dudareva, N. An alternative pathway contributes to phenylalanine biosynthesis in plants via a cytosolic tyrosine: Phenylpyruvate aminotransferase. Nat. Commun. 2013, 4, 2833. [Google Scholar] [CrossRef]
- Singh, P.; Preu, L.; Beuerle, T.; Kaufholdt, D.; Hänsch, R.; Beerhues, L.; Gaid, M. A promiscuous coenzyme A ligase provides benzoyl-coenzyme A for xanthone biosynthesis in Hypericum. Plant J. 2020, 104, 1472–1490. [Google Scholar] [CrossRef]
- El-Mawla, A.; Ahmed, M.; Beerhues, L. Benzoic acid biosynthesis in cell cultures of Hypericum androsaemum. Planta 2002, 214, 727–733. [Google Scholar] [CrossRef]
- Peters, S.; Schmidt, W.; Beerhues, L. Regioselective oxidative phenol couplings of 2,3′,4,6-tetrahydroxybenzophenone in cell cultures of Centaurium erythraea RAFN and Hypericum androsaemum L. Planta 1997, 204, 64–69. [Google Scholar] [CrossRef]
- Beerhues, L.; Liu, B. Biosynthesis of biphenyls and benzophenones–evolution of benzoic acid-specific type III polyketide synthases in plants. Phytochemistry 2009, 70, 1719–1727. [Google Scholar] [CrossRef]
- El-Awaad, I.; Bocola, M.; Beuerle, T.; Liu, B.; Beerhues, L. Bifunctional CYP81AA proteins catalyse identical hydroxylations but alternative regioselective phenol couplings in plant xanthone biosynthesis. Nat. Commun. 2016, 7, 11472. [Google Scholar] [CrossRef]
- Khattab, A.R.; Farag, M.A. Current status and perspectives of xanthones production using cultured plant biocatalyst models aided by in-silico tools for its optimization. Crit. Rev. Biotechnol. 2020, 40, 415–431. [Google Scholar] [CrossRef]
- Kitanov, G.M.; Nedialkov, P.T. Benzophenone O-glucoside, a biogenic precursor of 1,3,7-trioxygenated xanthones in Hypericum annulatum. Phytochemistry 2001, 57, 1237–1243. [Google Scholar] [CrossRef]
- Barillas, W.; Beerhues, L. 3-Hydroxybenzoate: Coenzyme A ligase and 4-coumarate: Coenzyme A ligase from cultured cells of Centaurium erythraea. Planta 1997, 202, 112–116. [Google Scholar] [CrossRef]
- Schmidt, W.; Abd El-Mawla, A.M.; Wolfender, J.L.; Hostettmann, K.; Beerhues, L. Xanthones in cell cultures of Hypericum androsaemum. Planta Med. 2000, 66, 380–381. [Google Scholar] [CrossRef]
- Velíšek, J.; Davidek, J.; Cejpek, K. Biosynthesis of food constituents: Natural pigments. Part 2—A review. Czech J. Food Sci. 2008, 26, 73. [Google Scholar] [CrossRef]
- Barillas, W.; Beerhues, L. 3-Hydroxybenzoate: Coenzyme A ligase from cell cultures of Centaurium erythraea: Isolation and characterization. Biol. Chem. 2000, 381, 155–160. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, W.; Peters, S.; Beerhues, L. Xanthone 6-hydroxylase from cell cultures of Centaurium erythraea RAFN and Hypericum androsaemum L. Phytochemistry 2000, 53, 427–431. [Google Scholar] [CrossRef] [PubMed]
- Jantan, I.; Pisar, M.M.; Idris, M.S.; Taher, M.; Ali, R.M. In vitro inhibitory effect of rubraxanthone isolated from Garcinia parvifolia on plateletactivating factor receptor binding. Planta Med. 2002, 68, 1133–1134. [Google Scholar] [CrossRef] [PubMed]
- Susanti, M.; Lena, D.I.; Dachriyanus, D. Development and validation of a HPLC method for determination and quantification of rubraxanthone in stem bark extract of mangosteen. Indones. J. Pharm. 2014, 25, 237–244. [Google Scholar] [CrossRef]
- Daud, S.; Karunakaran, T.; Santhanam, R.; Nagaratnam, S.R.; Jong, V.Y.M.; Ee, G.C.L. Cytotoxicity and nitric oxide inhibitory activities of xanthones isolated from Calophyllum hosei Ridl. Nat. Prod. Res. 2021, 35, 6067–6072. [Google Scholar] [CrossRef]
- Gómez-Verjan, J.C.; Rodríguez-Hernández, K.D.; Reyes-Chilpa, R. Bioactive Coumarins and Xanthones from Calophyllum genus and analysis of their Druglikeness and toxicological properties. Stud. Nat. Prod. 2017, 53, 277–307. [Google Scholar]
- Rukachaisirikul, V.; Phainuphong, P.; Sukpondma, Y.; Phongpaichit, S.; Taylor, W.C. Antibacterial caged-tetraprenylated xanthones from the stem bark of Garcinia scortechinii. Planta Med. 2005, 71, 165–170. [Google Scholar] [CrossRef]
- Araújo, J.; Fernandes, C.; Pinto, M.; Tiritan, M.E. Chiral derivatives of xanthones with antimicrobial activity. Molecules 2019, 24, 314. [Google Scholar] [CrossRef] [Green Version]
- Atkinson, J.; Gupta, P.; Lewis, J. Benzophenone participation in xanthone biosynthesis (Gentianaceae). Chem. Commun. 1968, 1386–1387. [Google Scholar] [CrossRef]
- Nualkaew, N.; Morita, H.; Shimokawa, Y.; Kinjo, K.; Kushiro, T.; De-Eknamkul, W.; Ebizuka, Y.; Abe, I. Benzophenone synthase from Garcinia mangostana L. pericarps. Phytochemistry 2012, 77, 60–69. [Google Scholar] [CrossRef]
- Mazimba, O.; Nana, F.; Kuete, V.; Singh, G.S. Xanthones and anthranoids from the medicinal plants of Africa. Med. Plant Res. Africa 2013, 393–434. [Google Scholar]
- Fiesel, T.; Gaid, M.; Müller, A.; Bartels, J.; El-Awaad, I.; Beuerle, T.; Ernst, L.; Behrends, S.; Beerhues, L. Molecular cloning and characterization of a xanthone prenyltransferase from Hypericum calycinum cell cultures. Molecules 2015, 20, 15616–15630. [Google Scholar] [CrossRef]
- Nagia, M.; Gaid, M.; Biedermann, E.; Fiesel, T.; El-Awaad, I.; Hänsch, R.; Wittstock, U.; Beerhues, L. Sequential regiospecific gem-diprenylation of tetrahydroxyxanthone by prenyltransferases from Hypericum sp. New Phytol. 2019, 222, 318–334. [Google Scholar] [CrossRef]
- Chen, D.; Chen, R.; Wang, R.; Li, J.; Xie, K.; Bian, C.; Sun, L.; Zhang, X.; Liu, J.; Yang, L.; et al. Probing the catalytic promiscuity of a regio-and stereospecific C-glycosyltransferase from Mangifera indica. Angew. Chem. 2015, 127, 12869–12873. [Google Scholar] [CrossRef]
- Ehianeta, T.S.; Laval, S.; Yu, B. Bio-and chemical syntheses of mangiferin and congeners. BioFactors 2016, 42, 445–458. [Google Scholar] [CrossRef]
- Birch, A.J.; Donovan, F.W. Studies in relation to biosynthesis. I. Some possible routes to derivatives of orcinol and phloroglucinol. Aust. J. Chem. 1953, 6, 360–368. [Google Scholar] [CrossRef]
- Roberts, J.C. Naturally Occurring Xanthones. Chem. Rev. 1961, 61, 591–605. [Google Scholar] [CrossRef]
- Khattab, A.R.; Farag, M.A. Marine and terrestrial endophytic fungi: A mine of bioactive xanthone compounds, recent progress, limitations, and novel applications. Crit. Rev. Biotechnol. 2022, 42, 403–430. [Google Scholar] [CrossRef]
- Chexal, K.K.; Fouweather, C.; Holker, J.S.; Simpson, T.J.; Young, K. The biosynthesis of fungal metabolites. Part III. Structure of shamixanthone and tajixanthone, metabolites of Aspergillus variecolor. J. Chem. Soc. Perkin. Trans. 1974, 1, 1584–1593. [Google Scholar] [CrossRef]
- Ishida, M.; Hamasaki, T.; Hatsuda, Y.; Fukuyama, K.; Tsukihara, T.; Katsube, Y. Epishamixanthone, a new metabolite from Aspergillus rugulosus. Agric. Biol. Chem. 1976, 40, 1051–1052. [Google Scholar] [CrossRef]
- Szwalbe, A.J.; Williams, K.; Song, Z.; de Mattos-Shipley, K.; Vincent, J.L.; Bailey, A.M.; Simpson, T.J. Characterisation of the biosynthetic pathway to agnestins A and B reveals the reductive route to chrysophanol in fungi. Chem Sci. 2019, 10, 233–238. [Google Scholar] [CrossRef] [PubMed]
- Anderson, J.A.; Lin, B.K.; Williams, H.J.; Scott, A.I. Deoxygenation of phenolic natural products. Enzymic conversion of emodin to chrysophanol. J. Am. Chem. Soc. 1988, 110, 1623–1624. [Google Scholar] [CrossRef]
- Anderson, J.A.; Lin, B.-K.; Shan, S.W. Purification and properties of emodin deoxygenase from Pyrenochaeta terrestris. Phytochemistry 1990, 29, 2415–2418. [Google Scholar] [CrossRef]
- Henry, K.M.; Townsend, C.A. Ordering the reductive and cytochrome P450 oxidative steps in demethylsterigmatocystin formation yields general insights into the biosynthesis of aflatoxin and related fungal metabolites. J. Am. Chem. Soc. 2005, 127, 3724–3733. [Google Scholar] [CrossRef]
- Brunauer, G.; Hager, A.; Krautgartner, W.D.; Tuerk, R.; Stocker-Wörgötter, E. Experimental studies on (L.) Zahlbr.: Chemical and microscopical investigations of the mycobiont and re-synthesis stages. Lichenologist 2006, 38, 577–585. [Google Scholar] [CrossRef]
- Belkheir, A.K.; Gaid, M.; Liu, B.; Hänsch, R.; Beerhues, L. Benzophenone synthase and chalcone synthase accumulate in the mesophyll of Hypericum perforatum leaves at different developmental stages. Front. Plant Sci. 2016, 7, 921. [Google Scholar] [CrossRef]
- Winkel-Shirley, B. Flavonoid biosynthesis. A colorful model for genetics, biochemistry, cell biology, and biotechnology. Plant Physiol. 2001, 126, 485–493. [Google Scholar] [CrossRef]
- Klundt, T.; Bocola, M.; Lütge, M.; Beuerle, T.; Liu, B.; Beerhues, L. A single amino acid substitution converts benzophenone synthase into phenylpyrone synthase. J. Biol. Chem. 2009, 284, 30957–30964. [Google Scholar] [CrossRef]
- Pasqua, G.; Avato, P.; Monacelli, B.; Santamaria, A.R.; Argentieri, M.P. Metabolites in cell suspension cultures, calli, and in vitro regenerated organs of Hypericum perforatum cv. Topas. Plant Sci. 2003, 165, 977–982. [Google Scholar] [CrossRef]
- Hose, E.; Clarkson, D.T.; Steudle, E.; Schreiber, L.; Hartung, W. The exodermis: A variable apoplastic barrier. J. Exp. Bot. 2001, 52, 2245–2264. [Google Scholar] [CrossRef]
- Barberon, M. The endodermis as a checkpoint for nutrients. New Phytol. 2017, 213, 1604–1610. [Google Scholar] [CrossRef]
- Brasili, E.; Miccheli, A.; Marini, F.; Pratico, G.; Sciubba, F.; Di Cocco, M.E.; Pasqua, G. Metabolic profile and root development of Hypericum perforatum L. in vitro roots under stress conditions due to chitosan treatment and culture time. Front. Plant Sci. 2016, 7, 507. [Google Scholar] [CrossRef]
- Huang, C.Y.; Chang, T.C.; Wu, Y.J.; Chen, Y.; Chen, J.J. Benzophenone and benzoylphloroglucinol derivatives from Hypericum sampsonii with anti-inflammatory mechanism of otogirinin A. Molecules 2020, 25, 4463. [Google Scholar] [CrossRef]
- Crockett, S.L.; Poller, B.; Tabanca, N.; Pferschy-Wenzig, E.M.; Kunert, O.; Wedge, D.E.; Bucar, F. Bioactive xanthones from the roots of Hypericum perforatum (common St John’s wort). J. Sci. Food Agric. 2011, 91, 428–434. [Google Scholar] [CrossRef]
- Iinuma, M.; Tosa, H.; Tanaka, T.; Yonemori, S. Two xanthones from roots of Calophyllum inophyllum. Phytochemistry 1995, 38, 725–728. [Google Scholar] [CrossRef]
- Campa, C.; Mondolot, L.; Rakotondravao, A.; Bidel, L.P.; Gargadennec, A.; Couturon, E.; La Fisca, P.; Rakotomalala, J.; Jay-Allemand, C.; Davis, A.P. A survey of mangiferin and hydroxycinnamic acid ester accumulation in coffee (Coffea) leaves: Biological implications and uses. Ann. Bot. 2012, 110, 595–613. [Google Scholar] [CrossRef]
- Pinto, M.M.M.; Sousa, E.P. Natural and synthetic xanthonolignoids: Chemistry and biological activities. Curr. Med. Chem. 2003, 10, 1–12. [Google Scholar] [CrossRef]
- Foti, M.C.; Sharma, S.K.; Shakya, G.; Prasad, A.K.; Nicolosi, G.; Bovicelli, P.; Ghosh, B.; Raj, H.G.; Rastogi, R.C.; Parmar, V.S. Biopolyphenolics as antioxidants: Studies under an Indo-Italian CSIR-CNR project. Pure Appl. Chem. 2005, 77, 91–101. [Google Scholar] [CrossRef]
- Boudet, A.M. Evolution and current status of research in phenolic compounds. Phytochemistry 2007, 68, 2722–2735. [Google Scholar] [CrossRef]
- Varache-Lembège, M.; Moreau, S.; Larrouture, S.; Montaudon, D.; Robert, J.; Nuhrich, A. Synthesis and antiproliferative activity of aryl-and heteroaryl-hydrazones derived from xanthone carbaldehydes. Eur. J. Med. Chem. 2008, 43, 1336–1343. [Google Scholar] [CrossRef] [PubMed]
- Castanheiro, R.A.; Silva, A.M.; Campos, N.A.; Nascimento, M.S.; Pinto, M.M. Antitumor activity of some prenylated xanthones. Pharmaceuticals 2009, 2, 33–43. [Google Scholar] [CrossRef] [PubMed]
- Fernandes, C.; Masawang, K.; Tiritan, M.E.; Sousa, E.; de Lima, V.; Afonso, C.; Bousbaa, H.; Sudprasert, W.; Pedro, M.; Pinto, M.M. New chiral derivatives of xanthones: Synthesis and investigation of enantioselectivity as inhibitors of growth of human tumor cell lines. Bioorg. Med. Chem. 2014, 22, 1049–1062. [Google Scholar] [CrossRef] [PubMed]
- Wong, K.W.; Mah, S.H. A review on xanthone derivatives with antiinflammatory effects and their structure–activity relationship. Stud. Nat. Prod. Chem. 2021, 68, 393–433. [Google Scholar]
- Palmeira, A.; Paiva, A.; Sousa, E.; Seca, H.; Almeida, G.M.; Lima, R.T.; Fernandes, M.X.; Pinto, M.; Vasconcelos, M.H. Insights into the in vitro antitumor mechanism of action of a new pyranoxanthone. Chem. Biol. Drug Des. 2010, 76, 43–58. [Google Scholar] [CrossRef]
- Lemos, A.; Leão, M.; Soares, J.; Palmeira, A.; Pinto, M.; Saraiva, L.; Sousa, M.E. Medicinal Chemistry Strategies to Disrupt the p53–MDM2/MDMX Interaction. Med. Chem. Res. 2016, 36, 789–844. [Google Scholar] [CrossRef]
- Leão, M.; Gomes, S.; Pedraza-Chaverri, J.; Machado, N.; Sousa, E.; Pinto, M.; Inga, A.; Pereira, C.; Saraiva, L. α-Mangostin and Gambogic Acid as Potential Inhibitors of the p53–MDM2 Interaction Revealed by a Yeast Approach. J. Nat. Prod. 2013, 76, 774–778. [Google Scholar] [CrossRef]
- Barua, A.; Choudhury, P.; Mandal, S.; Panda, C.K.; Saha, P. Therapeutic potential of xanthones from Swertia chirata in breast cancer cells. Indian J Med. Res. 2020, 152, 285. [Google Scholar]
- Zheng, X.Y.; Yang, Y.F.; Li, W.; Zhao, X.; Sun, Y.; Sun, H.; Pu, X.P. Two xanthones from Swertia punicea with hepatoprotective activities in vitro and in vivo. J. Ethnopharmacol. 2014, 153, 854–863. [Google Scholar] [CrossRef]
- Pinto, E.; Afonso, C.; Duarte, S.; Vale-Silva, L.; Costa, E.; Sousa, E.; Pinto, M. Antifungal Activity of Xanthones: Evaluation of their Effect on Ergosterol Biosynthesis by High-performance Liquid Chromatography. Chem. Biol. Drug Des. 2011, 77, 212–222. [Google Scholar] [CrossRef]
- Resende, D.I.S.P.; Pereira-Terra, P.; Moreira, J.; Freitas-Silva, J.; Lemos, A.; Gales, L.; Pinto, E.; De Sousa, M.E.; Da Costa, P.M.; Pinto, M.M.M. Synthesis of a Small Library of Nature-Inspired Xanthones and Study of Their Antimicrobial Activity. Molecules 2020, 25, 2405. [Google Scholar] [CrossRef]
- Urbatzka, R.; Freitas, S.; Palmeira, A.; Almeida, T.; Moreira, J.; Azevedo, C.; Afonso, C.; Correia-da-Silva, M.; Sousa, E.; Pinto, M.; et al. Lipid reducing activity and toxicity profiles of a library of polyphenol derivatives. Eur. J. Med. Chem. 2018, 151, 272–284. [Google Scholar] [CrossRef]
- Deeb, O.; Alfalah, S.; Clare, B.W. QSAR of aromatic substances: MAO inhibitory activity of xanthone derivatives. J. Enzyme Inhib. Med. Chem. 2007, 22, 277–286. [Google Scholar] [CrossRef]
- Martins, E.; Silva, V.; Lemos, A.; Palmeira, A.; Puthongking, P.; Sousa, E.; Rocha-Pereira, C.; Ghanem, C.I.; Carmo, H.; Remião, F.; et al. Newly Synthesized Oxygenated Xanthones as Potential P-Glycoprotein Activators: In Vitro, Ex Vivo, and In Silico Studies. Molecules 2019, 24, 707. [Google Scholar] [CrossRef]
- Saraiva, L.; Fresco, P.; Pinto, E.; Sousa, E.; Pinto, M.; Gonçalves, J. Inhibition of protein kinase C by synthetic xanthone derivatives. Bioorg. Med. Chem. 2003, 11, 1215–1225. [Google Scholar] [CrossRef]
- Rosa, G.P.; Palmeira, A.; Resende, D.I.S.P.; Almeida, I.F.; Kane-Pagès, A.; Barreto, M.C.; Sousa, E.; Pinto, M.M.M. Xanthones for melanogenesis inhibition: Molecular docking and QSAR studies to understand their anti-tyrosinase activity. Bioorg. Med. Chem. 2021, 29, 115873. [Google Scholar] [CrossRef]
- Pedro, M.; Cerqueira, F.; Sousa, M.E.; Nascimento, M.S.J.; Pinto, M. Xanthones as inhibitors of growth of human cancer cell lines and their effects on the proliferation of human lymphocytes in vitro. Bioorg. Med. Chem. 2002, 10, 3725–3730. [Google Scholar] [CrossRef]
- Cidade, H.; Rocha, V.; Palmeira, A.; Marques, C.; Tiritan, M.E.; Ferreira, H.; Lobo, J.S.; Almeida, I.F.; Sousa, M.E.; Pinto, M. In silico and in vitro antioxidant and cytotoxicity evaluation of oxygenated xanthone derivatives. Arab. J. Chem. 2020, 13, 17–26. [Google Scholar] [CrossRef]
- Silva, V.; Cerqueira, F.; Nazareth, N.; Medeiros, R.; Sarmento, A.; Sousa, E.; Pinto, M. 1,2-Dihydroxyxanthone: Effect on A375-C5 Melanoma Cell Growth Associated with Interference with THP-1 Human Macrophage Activity. Pharmaceuticals 2019, 12, 85. [Google Scholar] [CrossRef]
- Saraiva, L.; Fresco, P.; Pinto, E.; Sousa, E.; Pinto, M.; Gonçalves, J. Synthesis and in vivo modulatory activity of protein kinase C of xanthone derivatives. Bioorg. Med. Chem. 2002, 10, 3219–3227. [Google Scholar] [CrossRef]
- Gomes, S.; Raimundo, L.; Soares, J.; Loureiro, J.B.; Leão, M.; Ramos, H.; Monteiro, M.N.; Lemos, A.; Moreira, J.; Pinto, M.; et al. New inhibitor of the TAp73 interaction with MDM2 and mutant p53 with promising antitumor activity against neuroblastoma. Cancer Lett. 2019, 446, 90–102. [Google Scholar] [CrossRef] [PubMed]
- Baell, J.; Walters, M.A. Chemistry: Chemical con artists foil drug discovery. Nature 2014, 513, 481–483. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Omolo, J.J.; Johnson, M.M.; Van Vuuren, S.F.; De Koning, C.B. The synthesis of xanthones, xanthenediones, and spirobenzofurans: Their antibacterial and antifungal activity. Bioorg. Med. Chem. Lett. 2011, 21, 7085–7088. [Google Scholar] [CrossRef] [PubMed]
- Tocci, N.; D’Auria, F.D.; Simonetti, G.; Panella, S.; Palamara, A.T.; Debrassi, A.; Rodrigues, A.A.; Cechinel, V.F.; Sciubba, F.; Pasqua, G. Bioassay-guided fractionation of extracts from Hypericum perforatum in vitro roots treated with carboxymethylchitosans and determination of antifungal activity against human fungal pathogens. Plant Physiol. Biochem. 2013, 70, 342–347. [Google Scholar] [CrossRef]
- Winter, R.W.; Ignatushchenko, M.; Ogundahunsi, O.A.; Cornell, K.A.; Oduola, A.M.; Hinrichs, D.J.; Riscoe, M.K. Potentiation of an antimalarial oxidant drug. Antimicrob. Agents Chemother. 1997, 41, 1449–1454. [Google Scholar] [CrossRef]
- Lopez, S.; Bastida, J.; Viladomat, F.; Codina, C. Acetylcholinesterase inhibitory activity of some Amaryllidaceae alkaloids and Narcissus extracts. Life Sci. 2002, 71, 2521–2529. [Google Scholar] [CrossRef]
- Urbain, A.; Marston, A.; Queiroz, E.F.; Ndjoko, K.; Hostettmann, K. Xanthones from Gentiana campestris as new acetylcholinesterase inhibitors. Planta Med. 2004, 70, 1011–1014. [Google Scholar] [CrossRef]
- Reutrakul, V.; Chanakul, W.; Pohmakotr, M.; Jaipetch, T.; Yoosook, C.; Kasisit, J.; Napaswat, C.; Santisuk, T.; Prabpai, S.; Kongsaeree, P.; et al. Anti-HIV-1 constituents from leaves and twigs of Cratoxylum arborescens. Planta Med. 2006, 72, 1433–1435. [Google Scholar] [CrossRef]
- Reutrakul, V.; Anantachoke, N.; Pohmakotr, M.; Jaipetch, T.; Sophasan, S.; Yoosook, C.; Kasisit, J.; Napaswat, C.; Santisuk, T.; Tuchinda, P. Cytotoxic and anti-HIV-1 caged xanthones from the resin and fruits of Garcinia hanburyi. Planta Med. 2007, 73, 33–40. [Google Scholar] [CrossRef]
- Jiang, D.J.; Dai, Z.; Li, Y.J. Pharmacological effects of xanthones as cardiovascular protective agents. Cardiovas. Drug. Rev. 2004, 22, 91–102. [Google Scholar]
- Wang, Y.; Shi, J.-G.; Wang, M.-Z.; Che, C.-T.; Yeung, J.H.K. Mechanisms of the vasorelaxant effect of 1-hydroxy-2,3,5- trimethoxy-xanthone, isolated from a Tibetan herb, Halenia elliptica, on rat coronary artery. Life Sci. 2007, 81, 1016–1023. [Google Scholar] [CrossRef]
Figure 1.
Structure of 9H-xanthen-9-one. A-ring (carbons 1–4) and B-ring (carbons 5–8) are attached through an oxygen atom and a carbonyl group.
Figure 1.
Structure of 9H-xanthen-9-one. A-ring (carbons 1–4) and B-ring (carbons 5–8) are attached through an oxygen atom and a carbonyl group.

Figure 2.
Structure of 2-hydroxyxanthone, oxygenated xanthone.

Figure 3.
Structure of mangiferin, C-glycoside (a) and swertianolin, O-glycoside (b).

Figure 4.
Structure of allanxanthone A, prenylated xanthone.

Figure 5.
Structure of globulixanthone E, bisxanthone.

Figure 6.
Structure of kielcorin, xantholignoid.

Figure 7.
Structure of xanthofulvin, miscellaneous xanthone.

Figure 8.
Pathways involved in xanthone biosynthesis in plants. ?: unknown proteins; 1,3,5,6-THX: 1,3,5,6-tetrahydroxyxanthone; 1,3,5,8-THX: 1,3,5,8-tetrahydroxyxanthone; 1,3,5-THX: 1,3,5-trihydroxyxanthone; 1,3,6,7-THX: 1,3,6,7-THX-tetrahydroxyxanthone; 1,3,7-THX: 1,3,7-trihydroxyxanthone; 2,3′,4,6-THB: 2,3′,4,6-tetrahydroxybenzophenone; 2,4,6-THB: 2,4,6-trihydroxybenzophenone; 3-BZL: 3-benzoate-CoA ligase; 3-DHQS: 3-dehydroquianate synthase; 8-prenyl-1,3,6,7-THX: 8-prenyl-1,3,6,7-tetrahydroxyxanthone; ACC: acetyl-CoA carboxylase; ADT: arogenate dehydratase; B3′H: benzophenone 3′-hydroxylase; BD: benzaldehyde dehydrogenase; BPS: benzophenone synthase; BZL: benzoate-CoA ligase; CHL: cinnamoyl-CoA hydratase/lyase; CM: chorismate mutase; CNL: cinnamate-CoA ligase; CoASH: coenzyme A; CS: chorismate synthase; CYP81AA1/2: Cytochrome P450 oxydase 81AA1/2; DAHP: 3-deoxy-D-arabino-heptulosonate-7-phosphate; DAHPS: DAHP synthase; DHQD: 3-dehydroquinase dehydratase; EPSP: 5-enolpyruvylshikimate 3-phosphate; EPSPS: EPSP synthase; PAL: phenylalanine ammonia lyase; pCAT: plastidial cationic amino acid transporter; PDT: prephenate dehydratase; PPA-AT: prephenate aminotransferase; PPP: pentose phosphate pathway; PPY-AT: phenylpyruvate aminotransferase; PT8PX: 8-prenylxanthone-forming prenyltransferase; PTpat: patulone-forming prenyltransferase; SD: shikimate 5-dehydrogenase; SK: shikimate kinase; StrAT2: malonyl-CoA acyltransferase; StrGT9: norathyriol 6-O-glucosyltransferase; X6H: xanthone-6-hydroxylases; Xt1: norathyriol 6-O-glucoside; Xt2: norathyriol-6-O-(6′-Omalonyl)-glucoside.
Figure 8.
Pathways involved in xanthone biosynthesis in plants. ?: unknown proteins; 1,3,5,6-THX: 1,3,5,6-tetrahydroxyxanthone; 1,3,5,8-THX: 1,3,5,8-tetrahydroxyxanthone; 1,3,5-THX: 1,3,5-trihydroxyxanthone; 1,3,6,7-THX: 1,3,6,7-THX-tetrahydroxyxanthone; 1,3,7-THX: 1,3,7-trihydroxyxanthone; 2,3′,4,6-THB: 2,3′,4,6-tetrahydroxybenzophenone; 2,4,6-THB: 2,4,6-trihydroxybenzophenone; 3-BZL: 3-benzoate-CoA ligase; 3-DHQS: 3-dehydroquianate synthase; 8-prenyl-1,3,6,7-THX: 8-prenyl-1,3,6,7-tetrahydroxyxanthone; ACC: acetyl-CoA carboxylase; ADT: arogenate dehydratase; B3′H: benzophenone 3′-hydroxylase; BD: benzaldehyde dehydrogenase; BPS: benzophenone synthase; BZL: benzoate-CoA ligase; CHL: cinnamoyl-CoA hydratase/lyase; CM: chorismate mutase; CNL: cinnamate-CoA ligase; CoASH: coenzyme A; CS: chorismate synthase; CYP81AA1/2: Cytochrome P450 oxydase 81AA1/2; DAHP: 3-deoxy-D-arabino-heptulosonate-7-phosphate; DAHPS: DAHP synthase; DHQD: 3-dehydroquinase dehydratase; EPSP: 5-enolpyruvylshikimate 3-phosphate; EPSPS: EPSP synthase; PAL: phenylalanine ammonia lyase; pCAT: plastidial cationic amino acid transporter; PDT: prephenate dehydratase; PPA-AT: prephenate aminotransferase; PPP: pentose phosphate pathway; PPY-AT: phenylpyruvate aminotransferase; PT8PX: 8-prenylxanthone-forming prenyltransferase; PTpat: patulone-forming prenyltransferase; SD: shikimate 5-dehydrogenase; SK: shikimate kinase; StrAT2: malonyl-CoA acyltransferase; StrGT9: norathyriol 6-O-glucosyltransferase; X6H: xanthone-6-hydroxylases; Xt1: norathyriol 6-O-glucoside; Xt2: norathyriol-6-O-(6′-Omalonyl)-glucoside.

Figure 9.
Xanthone biosynthetic pathways in fungi. Shamixanthone (a) and agnestin pathway (b).

Figure 10.
Xanthone biosynthetic pathways in lichens. Lichexanthone-type lichen xanthone (a) and thiomielin-type lichen xanthone pathway (b).
Figure 10.
Xanthone biosynthetic pathways in lichens. Lichexanthone-type lichen xanthone (a) and thiomielin-type lichen xanthone pathway (b).


{kind=link}
{kind=link}
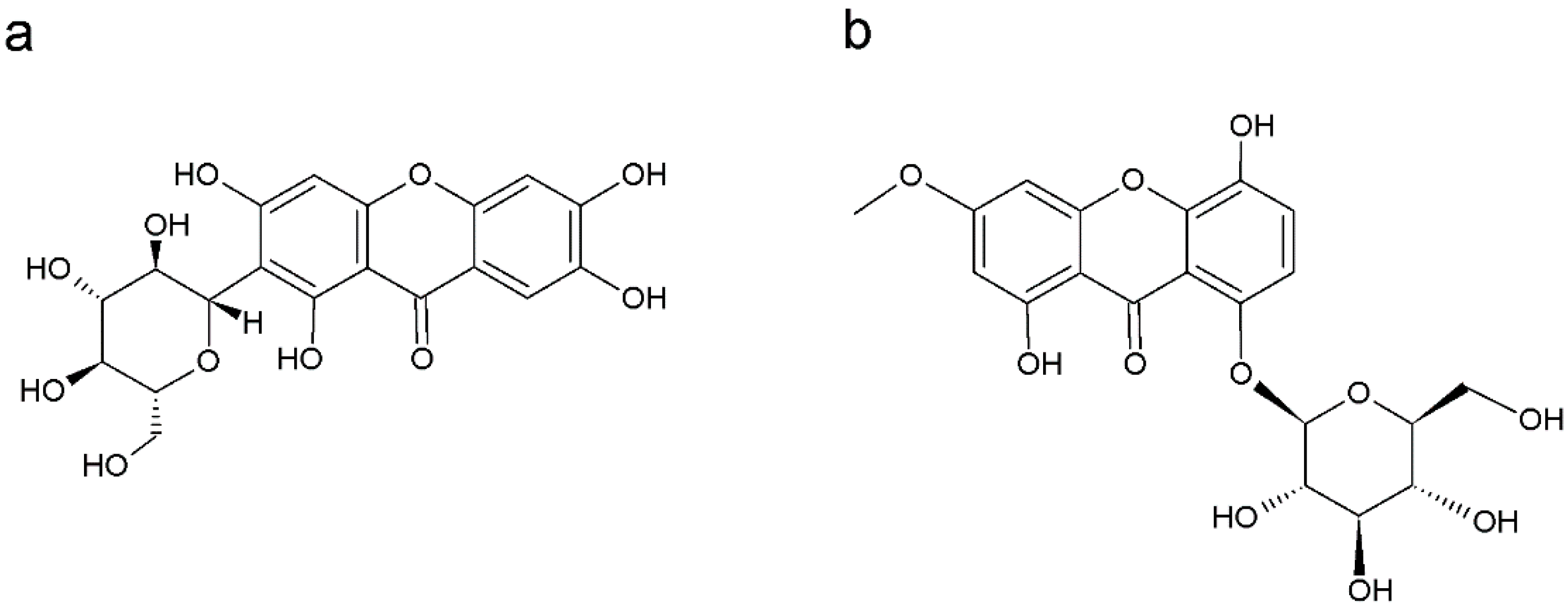{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
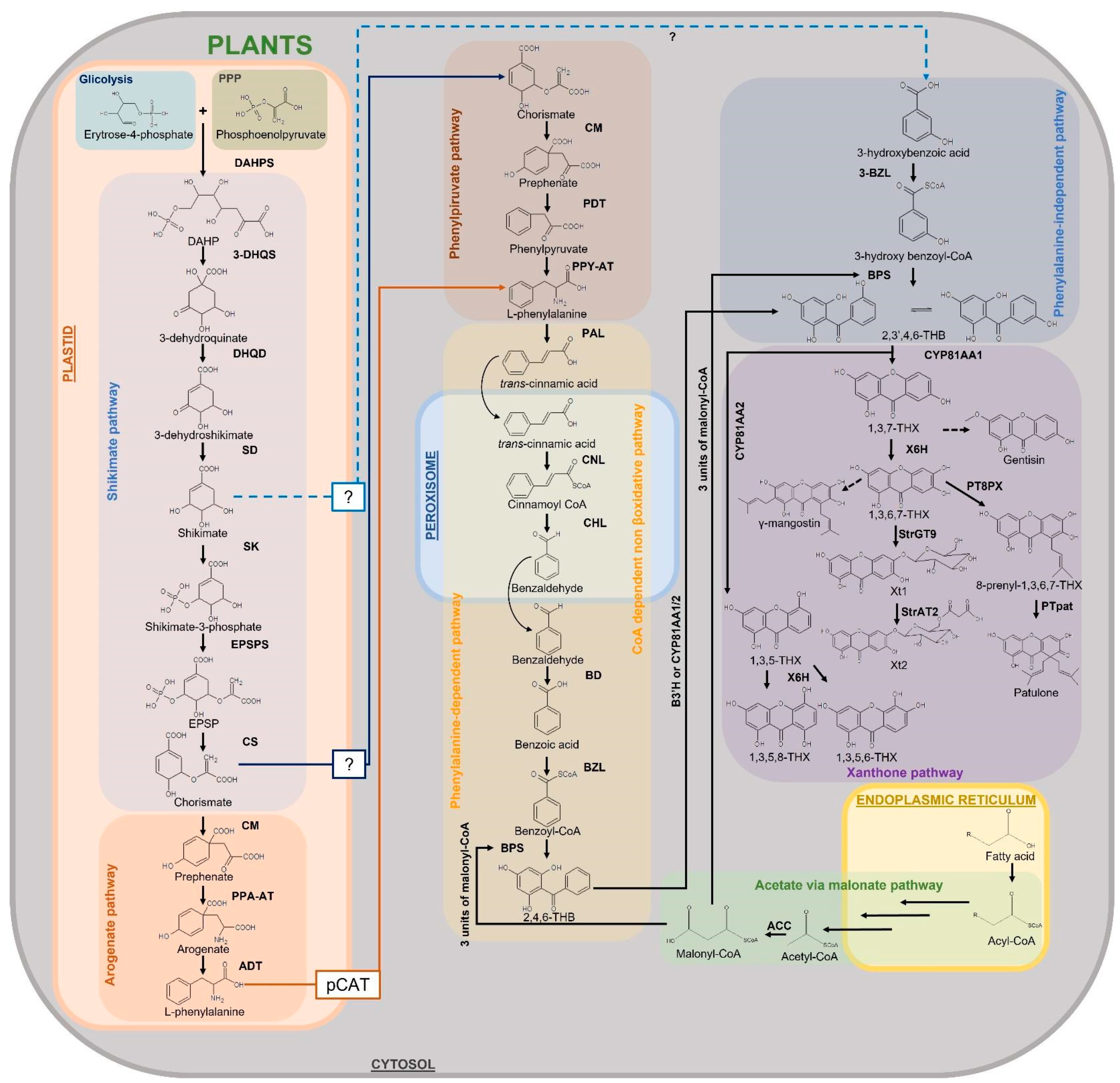{kind=link}
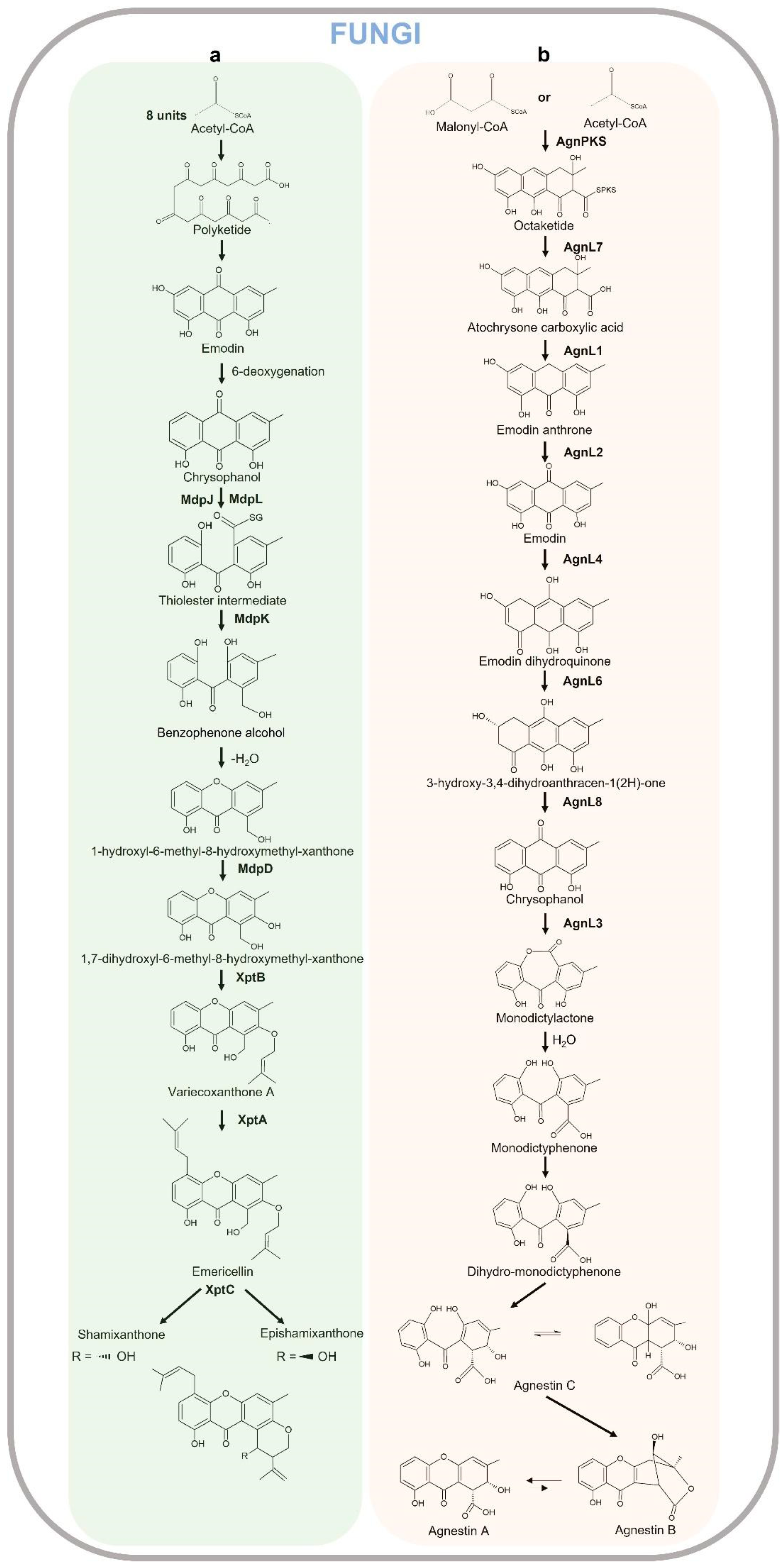{kind=link}
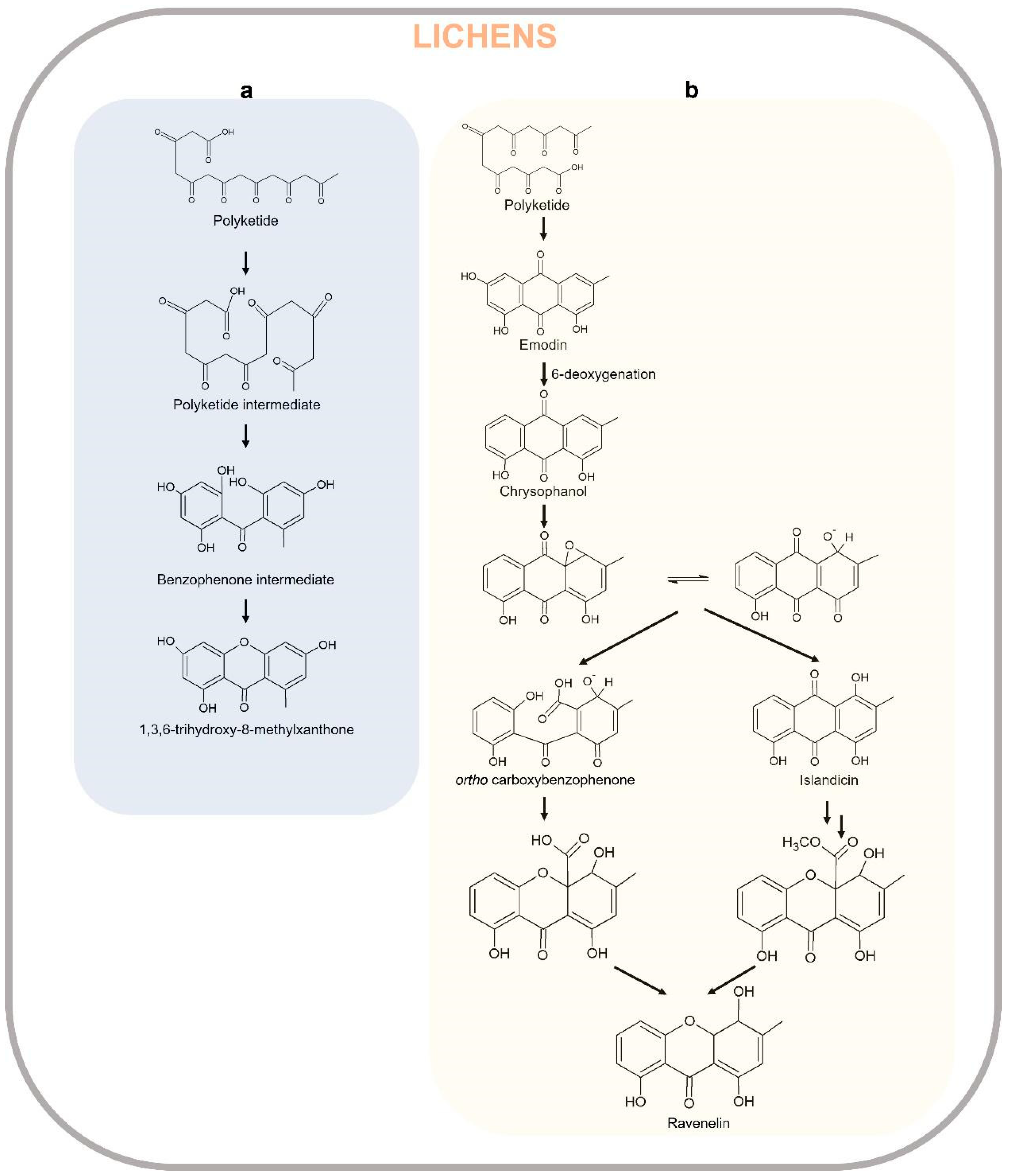{kind=link}
Table 1.
Xanthones in Plants.
| Plants | Xanthone | Organ/Tissue | References |
|---|---|---|---|
| Acanthaceae | Oxygenated xanthones | Root | |
| Andrographis paniculata (Burm.f.)Nees | [34,35] | ||
| Anacardiaceae | Xanthone glycosides | Fruit peel | |
| Mangifera indica L. | [36,37] | ||
| Annonaceae | Oxygenated xanthones Prenylated xanthones Xantholignoids | Fruit, resin, leaf, heartwood | |
| Anaxagorea luzonensis A.Gray | [38] | ||
| Orophea corymbosa Miq. | [39] | ||
| Asparagaceae | Xanthone glycosides, oxygenated xanthones Xantholignoids | Rootstock, apical part, bulb, tuber | |
| Anemarrhena asphodeloides Bunge | [40,41,42] | ||
| Drimiopsis maculata Lindl. & Paxton | [43] | ||
| Ledebouria graminifolia (Baker) Jessop | [44] | ||
| Asteraceae | Oxygenated xanthones Xantholignoids | Leaf | |
| Senecio mikanioides Otto ex Harv. | [45] | ||
| Bignoniaceae | Xanthone glycosides | Apical part | |
| Arrabidaea samydoides (Cham.) Sandwith | [46] | ||
| Bombacaceae | Xanthone glycosides | Leaf, flower | |
| Bombax ceiba L. | [47,48] | ||
| B. malabaricum DC. | [49] | ||
| Bonnetiaceae | Xanthone glycosides | Apical part | |
| Bonnetia dinizii Huber | [50] | ||
| Calophyllaceae Calophyllum apetalum Willd. | Oxygenated xanthones Xantholignoids | Heartwood stem bark, seed, root, wood | [51] |
| C. austroindicum Kosterm. ex P.F.Stevens | [52] | ||
| C. bracteatum Thwaites | [53] | ||
| C. brasiliense Vesque | [53,54,55,56,57,58,59,60] | ||
| C. calaba L. | [53,61] | ||
| C. canum Hook.f. ex T.Anderson | [62] | ||
| C. caledonicum Vieill. ex Planch. & Triana | [63,64,65,66,67] | ||
| C. castaneum P.F.Stevens | [9] | ||
| C. fragrans Ridl. | [68] | ||
| C. inophyllum L. | [69,70,71,72,73,74,75,76,77,78] | ||
| C. moonii Wight | [79] | ||
| C. neo-ebudicum Guillaumin | [80] | ||
| C. ramiflorum O.Schwarz | [81] | ||
| C. sclerophyllum Vesque | [82] | ||
| C. scriblitifolium M.R.Hend. & Wyatt-Sm. | [83] | ||
| C. tetrapterum Miq. | [84] | ||
| C. tomentosum Wight | [85,86] | ||
| C. zeylanicum Kosterm. | [87] | ||
| Caraipa densiflora Kubitzki | [88,89] | ||
| Haploclathra leiantha Benth. | [90,91] | ||
| H. paniculata Benth. | [92,93] | ||
| Kielmeyera coriacea Mart. & Zucc. | [88] | ||
| K. ferruginea A.P.B.Santos & Trad | [94] | ||
| K. rupestris Duarte | [95,96,97] | ||
| K. speciosa A.St.-Hil., A.Juss. & Cambess. | [98] | ||
| K. variabilis Mart. & Zucc. | [99] | ||
| Mesua ferrea L. | [78,100] | ||
| Cannabaceae | Bisxanthones | Bark | |
| Trema orientalis (L.) Blume | [101] | ||
| Caryophyllaceae | Oxygenated xanthones | Apical part | |
| Saponaria vaccaria L. | [102,103] | ||
| Celastraceae | Xanthone glycoside | Root | |
| Salacia reticulata Wight | [104] | ||
| Clusiaceae | Prenylated xanthones Xantholignoids Bisxanthones Oxygenated xanthones | Heartwood, stem bark, fruit, seeds, leaf, root | |
| Allanblackia floribunda Oliv. | [105,106] | ||
| A. monticola Mildbr. ex Engl. | [107,108] | ||
| Garcinia cowa Roxb. | [109,110] | ||
| G. echinocarpa Thwaites | [111] | ||
| G. forbesii King | [112] | ||
| G. mangostana L. | [113,114,115,116,117,118,119,120,121,122,123] | ||
| G. nobilis Engl. | [124] | ||
| G. opaca King | [125] | ||
| G. ovalifolia Oliv. | [126] | ||
| G. paucinervis Chun & F.C.How | [117] | ||
| G. pedunculata Roxb. ex Buch.-Ham. | [127] | ||
| G. quadrifaria Baill. ex Pierre | [128] | ||
| G. staudtii Engl. | [128] | ||
| G. terpnophylla Thwaites | [113] | ||
| G. vieillardii Pierre | [129] | ||
| G. xanthochymus Hook.f. | [130] | ||
| Pentadesma butyracea Sabine | [131] | ||
| Symphonia globulifera L.f. | [132,133] | ||
| Eriocaulaceae | Oxygenated xanthones | Apical part | |
| Leiothrix curvifolia (Bong.) Ruhland | [134] | ||
| L. flavescens (Bong.) Ruhland | [134] | ||
| Fabaceae | Xanthone glycosides Prenylated xanthones | Shoot | |
| Baphia kirkii Baker | [135] | ||
| Cyclopia genistoides (L.) R.Br. | [136] | ||
| C. intermedia E.Mey. | [136] | ||
| C. maculata (Andrews) Kies | [136] | ||
| C. sessiliflora Eckl. & Zeyh. | [136] | ||
| Gentianaceae | Oxygenated xanthones Xanthone glycosides | Seed, root, leaf, rhizome | |
| Canscora decussata Schult. | [78] | ||
| Centaurium erythraea Raf. | [137,138,139] | ||
| C. cachanlahuen B.L.Rob. | [140] | ||
| C. linarifolium (Lamark) G. Beck Frasera caroliniensis Walter | [78,141] [142] | ||
| Gentiana acaulis L. | [143] | ||
| G. lutea L. G. rhodantha Franch. | [144,145,146] [147] | ||
| Gentianella turkestanorum (Gand.) Holub Hoppea fastigiata Griseb. | [148] [149] | ||
| Swertia chirata Buch.-Ham. ex Wall. | [150,151] | ||
| S. mileensis T.N.Ho & W.L.Shih | [152] | ||
| S. punicea Hemsl. | [153,154] | ||
| S. purpurascens Wall. | [155] | ||
| S. randaiensis Hayata | [156] | ||
| S. japonica Makino | [156] | ||
| S. swertopsis Makino | [156] | ||
| S. davidii Franch. | [154,157] | ||
| Hypericaceae | Prenylated xanthones Oxygenated xanthones Xantholignoids Miscellaneous xanthones | Fruit, stem bark, root, leaf | |
| Cratoxylum cochinchinense (Lour.) Blume | [158,159] | ||
| C. formosum (Jack) Benth. & Hook.f. ex Dyer | [117,159,160] | ||
| C. pruniflorum Kurz | [161] | ||
| Harungana madagascariensis Lam. ex Poir. | [71] | ||
| Hypericum androsaemum L. | [162] | ||
| H. canariensis L. | [163] | ||
| H. geminiflorum Hemsl. H. japonicum Thunb | [164] [165] | ||
| H. maculatum Crantz | [166] | ||
| H. oblongifolium Choisy H. patulum Thunb. | [167] [168,169,170,171] | ||
| H. perforatum L. | [13,172] | ||
| H. reflexum L.f. | [173] | ||
| H. riparium A.Chev. | [174] | ||
| H. roeperianum G.W.Schimp. ex A.Rich. H. sampsonii Hance | [175] [176,177,178] | ||
| H. subalatum Hayata | [179] | ||
| Psorospermum adamauense Engl. Vismia guaramirangae Huber | [180] [181] | ||
| Iridaceae | Xanthone glycosides | Apical part | |
| Iris adriatica Trinajstić ex Mitić | [182] | ||
| I. albicans Lange | [183] | ||
| I. florentina L. | [184] | ||
| I. germanica L. | [185] | ||
| I. nigricans Dinsm. | [186] | ||
| Lamiaceae | Oxygenated xanthones | Roots | |
| Premna microphylla Turcz. | [187] | ||
| Lecythidaceae | Oxygenated xanthones | Wood, bark | |
| Gustavia hexapetala (Aubl.) Sm. | [188] | ||
| Moraceae | Oxygenated xanthones Prenylated xanthones Xantholignoids Xanthone glycoside | Root, twig, bark | |
| Cudrania cochinchinensis (Lour.) Yakuro Kudo & Masam. Monnina obtusifolia Kunth Tovomita brasiliensis Mart. | [189] [78,117] [190] | ||
| Nyssaceae | Oxygenated xanthones | Flower, fruit, stem, leaf | |
| Camptotheca acuminata Decne. | [191] | ||
| Poaceae | Xantholignoids | Apical part | |
| Chionochloa flavicans Zotov | [192] | ||
| Polygalaceae Polygala caudata Rehder & E.H.Wilson | Oxygenated xanthones | Root | [193] |
| P. sibirica L. | [194] | ||
| P. tenuifolia Willd. | [195] | ||
| P. vulgaris L. | [196] | ||
| Polypodiaceae | Xanthone glycoside | Whole plant | |
| Pyrrosia sheareri (Baker) Ching | [154] | ||
| Rubiaceae | Xanthone glycoside | Leaf | |
| Coffea pseudozanguebariae Bridson | [32,197] | ||
| Thymeleaceae | Xanthone glycoside | Leaf, stem | |
| Gnidia involucrata Steud. ex A.Rich. | [198] | ||
| Zingiberaceae | Prenylated xanthones | Rhizome | |
| Hedychium gardnerianum Sheppard ex Ker Gawl. | [199] |
Table 2.
Xanthones in Fungi.
| Fungi (Genus) | Xanthones | References |
|---|---|---|
| Actinomadura | Oxygenated xanthones Prenylated xanthones | [200,201] |
| Apiospora | Oxygenated xanthones | [202] |
| Aspergillus | Oxygenated xanthones | [3,203,204,205,206,207,208,209,210,211] |
| Chaetomium | Miscellaneous xanthones | [204] |
| Emericella | Oxygenated xanthones Prenylated xanthones Miscellaneous xanthones | [204,208,212] |
| Gibberella | Oxygenated xanthones | [213] |
| Guanomyces | Oxygenated xanthones | [214] |
| Humicola | Oxygenated xanthones | [215] |
| Monodictys | Oxygenated xanthones Miscellaneous xanthones | [204] |
| Paecilomyces | Prenylated xanthones | [204] |
| Penicillium | Oxygenated xanthones Miscellaneous xanthones | [204,205,216] |
| Phomopsis | Oxygenated xanthones | [217,218] |
| Phoma | Oxygenated xanthones | [204] |
| Wardomyces | Oxygenated xanthones Miscellaneous xanthones | [204,219] |
| Xylaria | Oxygenated xanthones Miscellaneous xanthones | [219] |
Table 3.
Xanthones in Lichens.
| Lichens (Genus) | Xanthones | References |
|---|---|---|
| Calopadia | Miscellaneous xanthones | [220] |
| Diploicia | Bixanthones | [221] |
| Lecanora | Oxygenated xanthones Miscellaneous xanthones | [209,222,223,224] |
| Lecidella | Oxygenated xanthones | [224] |
| Myriolecis | Oxygenated xanthones Miscellaneous xanthones | [223] |
| Micarea | Oxygenated xanthones | [10,209,224,225] |
| Pertusaria | Miscellaneous xanthones | [226] |
| Phyllopsora | Miscellaneous xanthones | [227] |
| Pseudoparmelia | Oxygenated xanthones Miscellaneous xanthones | [228] |
| Pyrenula | Oxygenated xanthones | [229] |
| Sporopodium | Miscellaneous xanthones | [230] |
| Teloschistale | Bixanthones | [2] |
| Umbilicaria | Xanthone glycosides | [231] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Badiali, C.; Petruccelli, V.; Brasili, E.; Pasqua, G. Xanthones: Biosynthesis and Trafficking in Plants, Fungi and Lichens. Plants 2023, 12, 694. https://doi.org/10.3390/plants12040694
AMA Style
Badiali C, Petruccelli V, Brasili E, Pasqua G. Xanthones: Biosynthesis and Trafficking in Plants, Fungi and Lichens. Plants. 2023; 12(4):694. https://doi.org/10.3390/plants12040694
Chicago/Turabian StyleBadiali, Camilla, Valerio Petruccelli, Elisa Brasili, and Gabriella Pasqua. 2023. "Xanthones: Biosynthesis and Trafficking in Plants, Fungi and Lichens" Plants 12, no. 4: 694. https://doi.org/10.3390/plants12040694
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.