
Asplenium pseudocapillipes (Aspleniaceae), a New Fern Species from South Korea


1
Department of Forest Science, Chungbuk National University, Chungdae-Ro 1, Seowon-Gu, Cheongju 28644, Chungbuk, Korea
2
Department of Ecological and Environmental System, Kyungpook National University, Gyeongsang-Daero, Sangju-Si 37224, Gyeongsangbuk-Do, Korea
*
Authors to whom correspondence should be addressed.
Plants 2022, 11(22), 3089; https://doi.org/10.3390/plants11223089
Submission received: 30 September 2022
/
Revised: 4 November 2022
/
Accepted: 11 November 2022
/
Published: 14 November 2022
(This article belongs to the Special Issue Diversity and Phylogenetic Relationships of Vascular Plants)
Abstract
:A new allotetraploid species of the genus Asplenium, A. pseudocapillipes, originated from the hybridization between A. capillipes and A. tenuicaule, has been newly discovered in two limestone areas of South Korea. A molecular phylogenetic analysis using one chloroplast region (rbcL) and three single- or low-copy nuclear regions (AK1, gapCp, pgiC) and a cytological analysis, including genome size measurements, were conducted to characterize this new species. From these results, the maternal origin of A. pseudocapillipes was confirmed to be A. capillipes, which has never been reported in Korea. All three nuclear data showed that this new species had genotypes of both A. capillipes and A. tenuicaule. The quantitative characteristics of the leaves showed values intermediate between the two parental species. The absence of gemma accorded with its paternal origin from A. tenuicaule, and 32 spores per sporangium accorded with its maternal origin from A. capillipes. Although A. pseudocapillipes has 32 spores per sporangium, it is considered to be a sexually reproducing, not an apomitic, fern.
1. Introduction
Polyploidization is considered an important factor in plant evolution [1,2,3,4]. Polyploids can be generated by autopolyploidization and allopolyploidization. Autopolyploidization occurs by doubling the genome within a species, but allopolyploidization occurs by interspecific hybridization followed by genome doubling [5]. In ferns, the ratio of speciation events related to polyploidy is higher (31%) than that in angiosperms (15%) [6]. Because of the frequent polyploidization and hybridization that occurs in ferns, they can sometimes experience reticulate evolution [7]. As a result, it can form a species complex that has morphological continuity between species [8]. To examine this morphological complexity in related taxa, many researchers have attempted to apply genome size measurement or low-copy nuclear DNA markers for genotyping analysis in ferns [9,10,11,12,13,14,15,16]. Through these attempts, new species, that were previously unknown and unrecognized due to their morphological similarities, have actually been identified [9,14,17].
Comprising approximately 700 species, Asplenium L. (Aspleniaceae) is a species-rich fern genus [18,19]. Members of Asplenium show various ploidy levels [20,21] and form a species complex comprising basic diploids and many auto- and allopolyploids by frequent reticulate evolution [9,10,14,15]. Moreover, it is possible to produce sterile hybrids between species with different ploidy levels or between homoploids in the same complex [15]. As the presence of numerous morphologically intermediate species in the genus Asplenium make it more difficult to distinguish one from another, researchers have attempted to solve this problem using various methods, including cytological and molecular methods [9,10,15,16,22,23].
To investigate Asplenium species, we collected plant samples from various habitats in Korea. Among them, we found plants that had been collected from the limestone area and identified as A. tenuicaule Hayata in the field based on their morphological characteristics. However, they were different from typical A. tenuicaule because the plants had an acute segment that is not generally observed in this taxon. This newly observed Asplenium sp. was more similar to A. capillipes Makino, except for the absence of gemma in the acroscopic axil.
In this study, cytological and molecular phylogenetic analyses were conducted to investigate the identity of this enigmatic species. Our goals in this paper were (1) to verify whether the newly found Asplenium sp. originated from hybridization; (2) to identify the parent species if it was a hybrid taxon; and (3) to describe it in detail if it was recognized as a new species.
2. Results
2.1. Chloroplast Phylogeny Based on rbcl
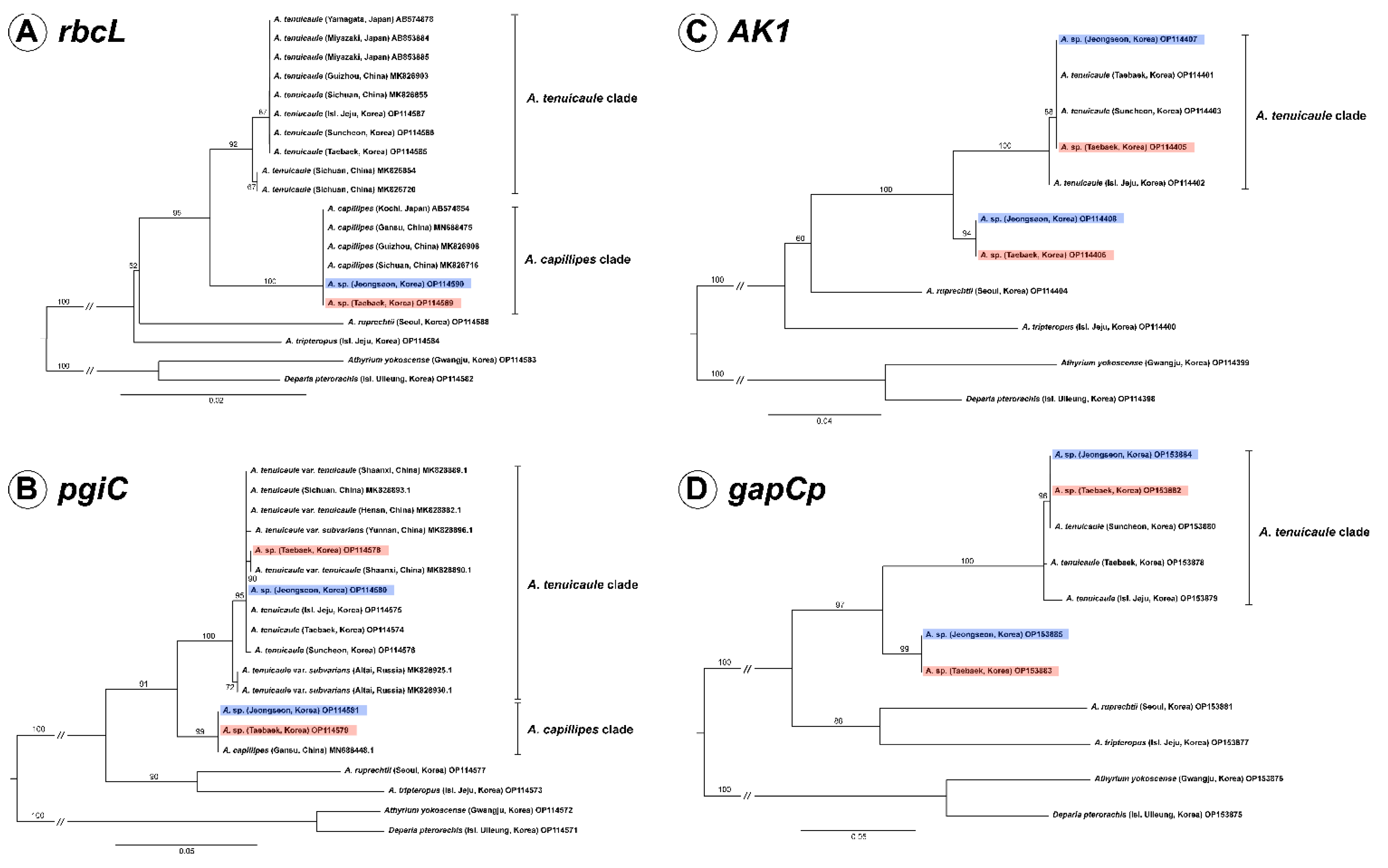
We determined the rbcL sequences of the new taxon Asplenium sp. found in Korea and its related species in this study and compared them together with the sequences of A. capillipes and A. tenuicaule reported from Japanese and Chinese samples. From the result, it was confirmed that the sequence of new taxon was identical to those of A. capillipes of Japan and China. The aligned sequence length of rbcL was 1272 bp with 1090 conserved sites and 63 parsimony-informative sites. The phylogenetic analysis showed that two individuals of Asplenium sp. were nested in the A. capillipes clade comprising Chinese and Japanese specimens with 100% ML bootstrap support, and the A. capillipes clade was identified as being sister to the A. tenuicaule clade (Figure 1A).
2.2. Nuclear Phylogenies Based on pgiC, AK1, and gapCp
After alignment, the lengths of the nuclear genes pgiC, AK1, and gapCp were found to be 694 bp, 866 bp, and 1221 bp, respectively. In pgiC, one copy of Asplenium sp. from each population (non-T type) was completely identical to the pgiC of Chinese A. capillipes, except for the 1 bp deletion of poly T in the Jeongseon population (CBNU2020-0171A). The other copy of Asplenium sp. in each population was almost identical to A. tenuicaule, therefore, we designated this copy as T type (Supplementary Figure S1). In the phylogeny based on pgiC, the T types of Asplenium sp. were nested in the A. tenuicaule clade, and the non-T types were positioned in the A. capillipes clade with an ML bootstrap support >99% (Figure 1B).
In the cases of AK1 and gapCp, each population of Asplenium sp. had two copies. One copy was identical to the sequence of A. tenuicaule, but the other was obviously different. The phylogeny based on AK1 and gapCp showed that the T types of Asplenium sp. and A. tenuicaule were also strongly supported as being monophyletic, and the non-T types of Asplenium sp. formed an independent clade with an ML bootstrap support >94% (Figure 1C,D).
2.3. Ploidy Analysis and Spore Observation
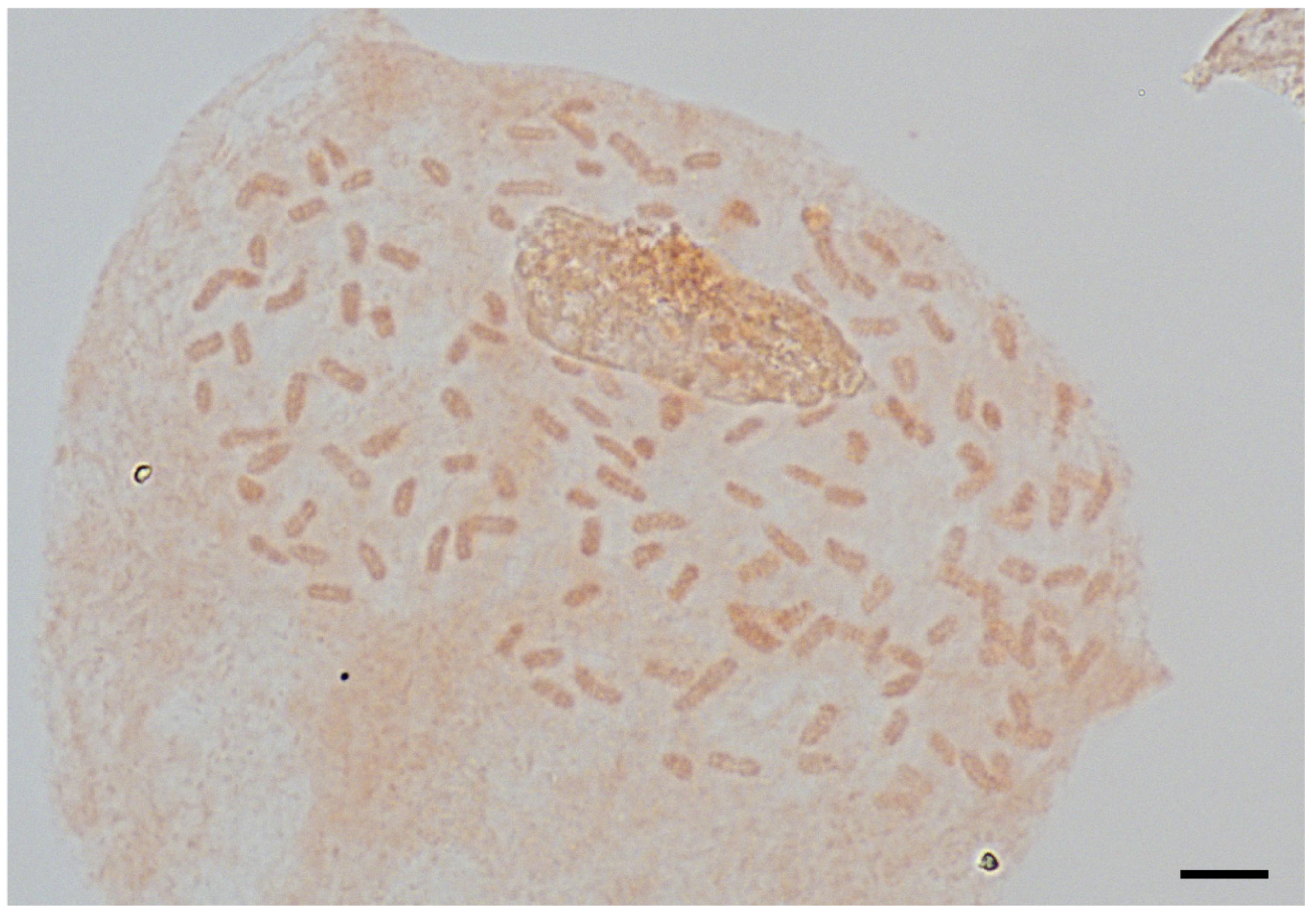
The mitotic chromosome number of Asplenium sp. was 2n = 144. Therefore, it was identified as being tetraploid (Figure 2). The genome size of Asplenium sp. with 16 individuals and A. tenuicaule with 4 individuals were measured and the mean C-values were 8.875 ± 0.06 pg and 4.079 ± 0.05 pg, respectively (Figure 3).
In addition, 32 spores were observed per sporangium (Figure 4E), with a mean size of 52.94 ± 3.02 μm ranging 46–59 μm, and they successfully germinated (Supplementary Figure S2).
Etymology: the species resembles A. capillipes.
From the results, we conclude Asplenium sp. should be treated as the new allotetraploid species Asplenium pseudocapillipes (Figure 4), which is described as follows.
2.4. Taxonomic Treatment
Asplenium pseudocapillipes S. H. Park, J. S. Kim & H. T. Kim, sp. nov. Type: South Korea, Taebaek-si, Hasami-dong, Deokhang Mountain, 770 m alt., 30 April 2020, (holotype CBNU2020-0106)
Diagnosis: Asplenium pseudocapillipes is allotetraploid, originating as the hybrid between A. capillipes and A. tenuicaule, and generally shows the intermediate form of both species (Table 1). This species has an acute segment apex and 32 spores per sporangium, which are similar to those of A. capillipes, but a gemma of the acroscopic axil does not form as in A. tenuicaule.
Plants are evergreen and epilithic. Plants (1.5-3-9(-12) cm tall. Rhizome shortly erect, apex scaly; scales dark brown to black, triangular to narrowly triangular. Fronds caespitose, herbaceous, green, subglabrous; stipe slender, sulcate adaxially, (0.3–)0.5–3(–5) cm; lamina triangular to narrowly triangular, (0.8-)1.5-6(-7) × (0.5-)0.7-2.5(-3) cm, 2 or almost three-pinnate, apex acute to acuminate, without gemma; pinnae (3-)4-9(-11) pairs, alternate or opposite, stalk slender, basal pinnae not reduced (or often slightly reduced), broadly ovate to triangular, 3-11(-15) × 2.5-7(-9) mm in basal pinna, pinnate to two-pinnatifid; ultimate segments (1-)1.5-3(-5) × 1-2.5(-3.5) mm, base cuneate, margin entire, apex mucronate to acute. Sori, one (or two) per ultimate segment or pinnule, basal to median on subtending vein, linear to narrowly oblong, (0.8-)1-1.8(-2.5) mm; indusia whitish to whitish-yellow, membranous, margins entire, opening toward costa, persistent. Spores 32 per sporangium, longer diameter averaging 52.94 μm, ranging 46-59 μm, excluding perispores. Tetraploid, 2n = 144.
Distribution and habitat: Two populations of A. pseudocapillipes have been reported from the limestone areas of Korea in Gangwon-do, Jeongsun-gun, and Taebaek-si. It usually grows by forming clusters on mossy rocks under moist forests.
3. Discussion
3.1. Discovering New Allotetraploid in Genus Asplenium
In this study, we found a new allotetraploid species, A. pseudocapillipes, from two populations in limestone areas of Korea. It was different from the hybrid species Asplenium × capillicaule Fraser-Jenk. from Nepal [25] and Japan [26] between A. capillipes and A. tenuicaule because Asplenium × capillicaule is known to have a gemma in acroscopic axils, which is similar to that of A. capillipes.
The A. pseudocapillipes found in Korea had two distinct copies of all three nuclear genes tested in the present study. The T types or non-T types (pgiC) of A. pseudocapillipes formed a clade with the counterparts of A. tenuicaule and A. capillipes, respectively. Maternally inherited rbcL was shown to be the maternal parent of A. pseudocapillipes. Based on the number of chromosomes, genome size, and fertile spores of A. pseudocapillipes, it is an allotetraploid between A. capillipes and A. tenuicaule because both parent species are diploids [18,24,26], excluding the tetraploid of A. capillipes in the Himalayas [27].
Allotetraploid A. pseudocapillipes may originate by two unreduced gametes of a homoploid hybrid (Figure 5A) or by a triploid bridge (Figure 5B). Because we have not yet found any sterile hybrids of diploid or triploid A. pseudocapillipes, it is not easy to establish a speciation model for this allotetraploid. The paternal species A. tenuicaule grows closely with A. pseudocapillipes in Korea, whereas the maternal species A. capillipes has not been reported in Korea yet. This implies that A. pseudocapillipes moved into Korea after speciation or that A. capillipes became extinct in Korea after speciation. However, A. capillipes is small and grows among moss with a high morphological similarity to A. tenuicaule [18] and A. pseudocapillipes. Therefore, this species may have been easily overlooked or was identified as A. tenuicaule even though we did not find any A. capillipes, which was identical to our original description, from herbarium specimens survey. If A. capillipes grew closely with A. tenuicaule and hybridization and polyploidization recently occurred, the relatively low genetic diversity of nuclear sequences between parents and offspring is to be expected.
In this study, we compared new allotetraploids to their parental species using morphological characters and cytological and molecular analyses. Further studies including gametogenesis and sporogenesis are required to deepen our understanding of this species.
3.2. Reproductive Mode of Asplenium Pseudocapillipes
During sporogenesis, most sexual reproductive leptosporangiate ferns produce 16 spore mother cells through four premeiotic mitoses and a total of 64 haploid spores per sporangium through meiosis [28]. In contrast, apomictic ferns have 32 spores with the same number of chromosomes as the parent sporophyte either by premeiotic endomitosis [29,30] or meiotic first division restitution [31]. Therefore, 32 spores per sporangium were considered to be presumptive evidence of apomictic ferns [29,30,32]. However, some species have been confirmed to be sexually reproductive ferns, even though they have 32 spores, such as Lindsaeaceae species [33] and Cystodium sorbifolium [34], or 16 spores, such as Alsophila species [35]. Lin, Kato, and Iwatsuki [34] deduced that the formation of eight spore mother cells was due to the reduction in premeiotic mitosis divisions from four to three and referred to these species types as “32-spored sexual type” or “lindsaeoid type”.
In Asplenium, there are some species with 32 spores, such as A. monodon [36], A. monanthes, A. palmeri [10], A. heterochroum, A. resilens [37], and A. cheilosorum [38], which are similar to A. pseudocapillipes found in the present study. Therefore, this poses the question of whether A. pseudocapillipes is agamosporous. To answer this question, we need to focus on the parents. A. capillipes is the maternal parent of A. pseudocapillipes, and is considered to be an apogamous species because it has 32 spores per sporangium [39]. However, half of the parental chromosomes in meiosis I, the normal meiotic division, and eight spore mother cells imply that this species is sexual in reproduction [24]. In contrast, A. tenuicaule, the paternal parent of A. pseudocapillipes, normally produces 64 spores in the sporangium. Therefore, it was suggested that the feature of “32-spored sexual type” found in A. pseudocapillipes was likely to be inherited from its paternal parent A. capillipes.
4. Materials and Methods
4.1. Plant Materials and Observation of Morphological Characteristics
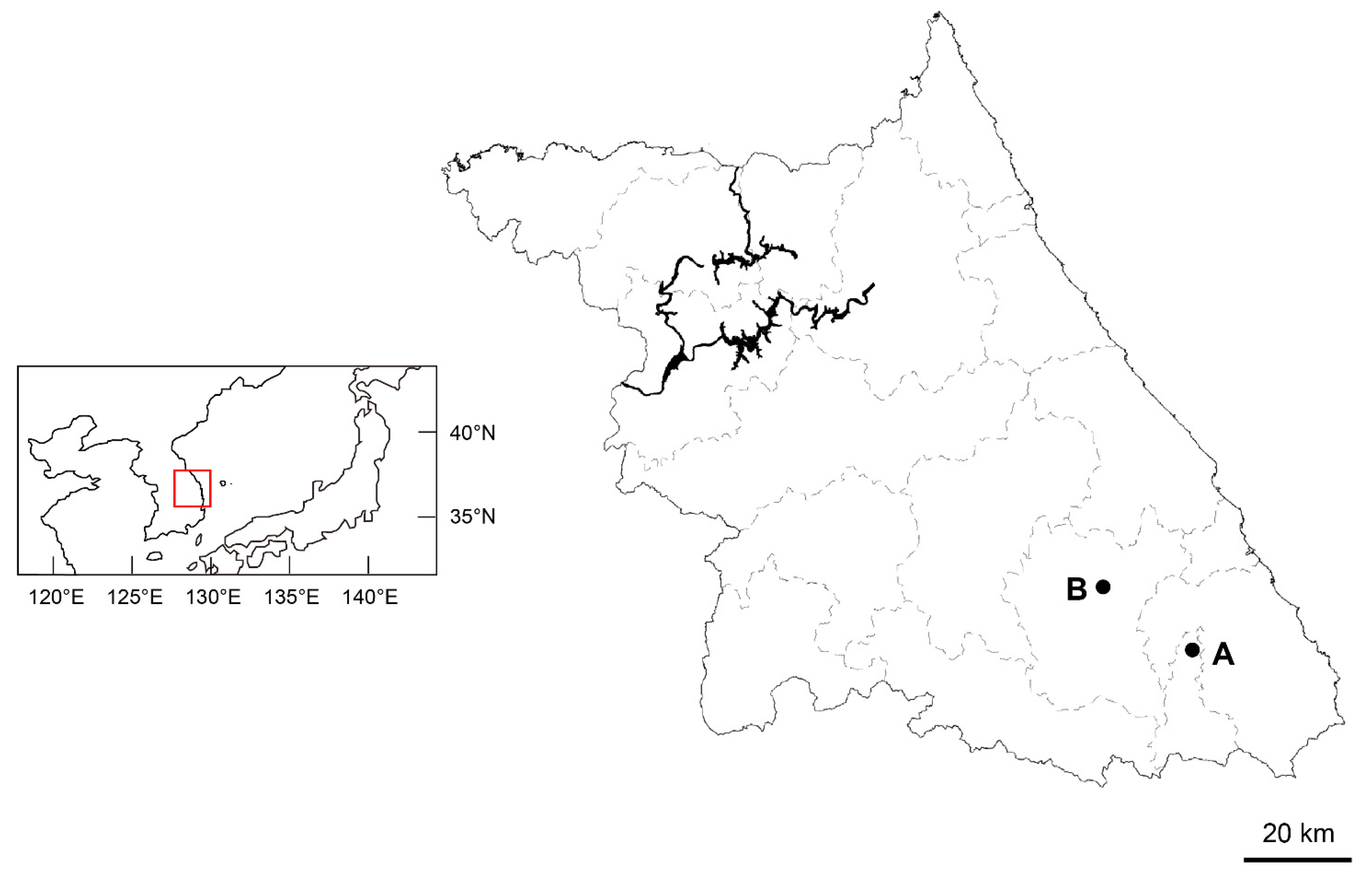
Except for the populations mentioned in the introduction, we found another population of Asplenium sp. and A. tenuicaule within a 10 m radius in the limestone areas of Gangwon-do, Korea (Figure 6). Living samples were collected from each population and transplanted into the greenhouse at Chungbuk National University (Cheongju, Chungbuk, Korea), and voucher specimens (Table 2) were deposited in the herbarium of the Chungbuk National University (CBNU). One individual from each of the two populations of the new taxon and three diploid Asplenium species (A. tenuicaule, A. ruprechtii Kurata, and A. tripteropus Nakai) were used for the molecular analysis. Two diploid Athyriaceae species (Athyrium yokoscense (Franch. & Sav.) Christ, and Deparia pterorachis (Christ) M. Kato were used as outgroups for the phylogenetic analysis (Table 2).
Rhizome scales and spores were observed using a light microscope (Olympus BX50, Tokyo, Japan). A total of 35 spores were randomly sampled under a light microscope (Olympus BX50) to measure the spore size based on the length of the long axis, excluding the perispore.
4.2. Chromosome Counting and Measurement of the Genome Size
For the observation of the mitotic chromosomes, the root tips were pretreated using 2 mM 8-hydroxyquinoline solution for 2 h and then fixed in Carnoy’s solution for 12 h. The fixed root tips were washed with 70% ethanol, macerated in 1 N HCl for 5 min, and then stained using the squashed method with 1% aceto-orcein. The slides were examined at 1000× magnification and then captured using a light microscope (Olympus BX50).
4.3. DNA Extraction, PCR Amplification, and Cloning
Genomic DNA was extracted from the leaves dried with silica gel using a DNeasy Plant Mini Kit (Qiagen, Hilden, Germany) following the manufacturer’s protocol. Based on maternally inherited chloroplast genomes and biparentally inherited nuclear genomes in the genus Asplenium, chloroplast rbcL and nuclear pgiC regions were amplified to clarify the parent species, given that the sequences of two putative parent species, namely A. capillipes and A. tenuicaule, have been previously reported (Figure 1A,B). Nuclear AK1 and gapCp regions were amplified to confirm the hybrid origin of the new taxon of Asplenium sp. Primer sets for rbcL, 1FN [40], and 1361R [41], AK1, AK_4F, and AK_R2 [42], gapCp, ESGAPCP8F1, and ESGAPCP11R1 [43], and pgiC, 14F, and 16R [44] were used for the PCR amplification. However, given that the pgiC amplification was not successful for Asplenium sp., the new primer set, 14F2 (5′-GAGTGTTTGGAATGTTTCCTTC-3′) and 16R3 (5′-GAGGAATGCCATCTATTGAA-3′), was newly designed for this study. The reaction mixture comprised 10 μL of AccuPower® PCR Premix (Bioneer, Daejeon, Korea), 1 μL of DNA, 1 μL of each primer (10 pM), and distilled water to a total volume of 20 μL. The PCR conditions were as follows: a total of 5 min denaturation at 95 °C, followed by 30 cycles at 95 °C for 45 s, 53 °C for 20 s, 72 °C for 60 s, followed by a final extension step at 72 °C for 10 min. Except for nuclear markers of Asplenium sp., the PCR products were purified using Expin™ PCR SV (GeneAll, Seoul, Korea) and sequenced using the AB1 3730xl System (Macrogen, Seoul, Korea). The PCR products of nuclear markers of Asplenium sp. were purified using the PureLink™ PCR purification kit (Invitrogen, Waltham, MA, USA) to remove primer dimers and small fragments of less than 300 bp. To obtain accurate sequences of nuclear markers, cloning was performed using the TOPO™ TA Cloning™ Kit (Invitrogen) following the manufacturer’s protocol. At least 10 colonies were randomly selected from each plate and grown in liquid medium. Plasmids were extracted using Exprep™ Plasmid SV (GeneAll) and sequenced with a universal M13 primer using the AB1 3730xl System (Macrogen).
4.4. Sequence Alignments and Phylogenetic Analyses
Each of the cloned nuclear genes was divided into two types, and a consensus sequence of each type was generated with a threshold of 50% in Geneious Prime software (ver. 2022.0.2) [45]. One chloroplast and three nuclear genes were aligned using MAFFT [46] and MUSCLE [47], respectively. For the phylogenetic analysis, the best-fit model of nucleotide substitution for each dataset was determined using ModelFinder [48] and a maximum likelihood analysis was performed using IQ-Tree [49] with 1000 ultrafast bootstraps [50].
4.5. Genome Size Measurement
The genome sizes of Asplenium sp. and A. tenuicaule were measured using a CyFlow® Ploidy Analyzer (Sysmex-Partec, Munster, Germany). Tetraploid Solanum tuberosum L., 1C = 1.82 pg [51], was used as the internal standard. Young fresh leaf tissues of the two Asplenium species and S. tuberosum were collected from living samples. They were washed using distilled water and chopped with razor blades in 500 μL of CyStain UV Precise P nuclei isolation buffer (Sysmex-Partec). After 10 min of incubation on ice, the suspension was then filtered through a nonsterile CellTrics® 30 μm filter and stained using 2 mL of UV Precise P staining buffer (Sysmex-Partec) containing DAPI. The stained suspension was loaded onto a flow cytometer, and the genome size was measured.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/plants11223089/s1, Figure S1: Comparison of pgiC gene sequence after editing the common Indels and outgroups in the original alignment among the Asplenium sp. and its putative parental species; Figure S2: Germinated spores of A. pseudocapillipes in the 1/2 MS medium 45 days after sowing. Scale bar = 0.1 mm.
Author Contributions
Conceptualization, H.T.K. and J.S.K.; field investigation, S.H.P., H.T.K. and J.S.K.; genome size measuring, chromosome counting, and sequencing, S.H.P.; data confirmation, H.T.K. and J.S.K.; writing—original draft preparation, S.H.P.; writing—review and editing, H.T.K. and J.S.K.; visualization, S.H.P.; supervision, H.T.K. and J.S.K.; project administration, H.T.K.; and funding acquisition, H.T.K. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by the National Institute of Biological Resources (NIBR) funded by the Korea Government (no. NIBR202212101).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Data generated in the present study were deposited in the NCBI database and available from the website (accessed on 1 December 2022, https://www.ncbi.nlm.nih.gov/nuccore/).
Conflicts of Interest
The authors declare no conflict of interest.
References
- Madlung, A. Polyploidy and its effect on evolutionary success: Old questions revisited with new tools. Heredity (Edinburgh) 2013, 110, 99–104. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Otto, S.P. The evolutionary consequences of polyploidy. Cell 2007, 131, 452–462. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Soltis, P.S.; Soltis, D.E. The role of hybridization in plant speciation. Annu. Rev. Plant Biol. 2009, 60, 561–588. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Soltis, P.S.; Marchant, D.B.; Van de Peer, Y.; Soltis, D.E. Polyploidy and genome evolution in plants. Curr. Opin. Genet. Dev. 2015, 35, 119–125. [Google Scholar] [CrossRef] [Green Version]
- Stebbins , G.L., Jr. Types of polyploids: Their classification and significance. Adv. Genet. 1947, 1, 403–429. [Google Scholar]
- Wood, T.E.; Takebayashi, N.; Barker, M.S.; Mayrose, I.; Greenspoon, P.B.; Rieseberg, L.H. The frequency of polyploid speciation in vascular plants. Proc. Natl. Acad. Sci. USA 2009, 106, 13875–13879. [Google Scholar] [CrossRef] [Green Version]
- Linder, C.R.; Rieseberg, L.H. Reconstructing patterns of reticulate evolution in plants. Am. J. Bot. 2004, 91, 1700–1708. [Google Scholar] [CrossRef] [Green Version]
- Barrington, D.S.; Haufler, C.H.; Werth, C.R. Hybridization, Reticulation, and Species Concepts in the Ferns. Am. Fern J. 1989, 79, 55–64. [Google Scholar] [CrossRef] [Green Version]
- Chang, Y.; Ebihara, A.; Lu, S.; Liu, H.; Schneider, H. Integrated taxonomy of the Asplenium normale complex (Aspleniaceae) in China and adjacent areas. J. Plant Res. 2018, 131, 573–587. [Google Scholar] [CrossRef]
- Dyer, R.J.; Savolainen, V.; Schneider, H. Apomixis and reticulate evolution in the Asplenium monanthes fern complex. Ann. Bot. 2012, 110, 1515–1529. [Google Scholar] [CrossRef] [Green Version]
- Ebihara, A.; Ishikawa, H.; Matsumoto, S.; Lin, S.J.; Iwatsuki, K.; Takamiya, M.; Watano, Y.; Ito, M. Nuclear DNA, chloroplast DNA, and ploidy analysis clarified biological complexity of the Vandenboschia radicans complex (Hymenophyllaceae) in Japan and adjacent areas. Am. J. Bot. 2005, 92, 1535–1547. [Google Scholar] [CrossRef] [PubMed]
- Hori, K.; Tono, A.; Fujimoto, K.; Kato, J.; Ebihara, A.; Watano, Y.; Murakami, N. Reticulate evolution in the apogamous Dryopteris varia complex (Dryopteridaceae, subg. Erythrovariae, sect. Variae) and its related sexual species in Japan. J. Plant Res. 2014, 127, 661–684. [Google Scholar] [PubMed]
- Hori, K. Hybrid origin of some species in the Deparia okuboana complex (Athyriaceae, Polypodiidae) verified with DNA analysis. Hikobia 2018, 17, 315–320. [Google Scholar]
- Liang, S.; Zhang, X.; Wei, R. Integrative taxonomy resolved species delimitation in a fern complex: A case study of the Asplenium coenobiale complex. Biodivers. Sci. 2019, 27, 1205–1220. [Google Scholar]
- Liang, S.Q.; Viane, R.L.; Zhang, X.C.; Wei, R. Exploring the reticulate evolution in the Asplenium pekinense complex and the A. varians complex (Aspleniaceae). J. Syst. Evol. 2021, 59, 125–140. [Google Scholar] [CrossRef]
- Ohlsen, D.J.; Perrie, L.R.; Shepherd, L.D.; Brownsey, P.J.; Bayly, M.J. Investigation of species boundaries and relationships in the Asplenium paleaceum complex (Aspleniaceae) using AFLP fingerprinting and chloroplast and nuclear DNA sequences. Aust. Syst. Bot. 2015, 27, 378–394. [Google Scholar] [CrossRef] [Green Version]
- Ebihara, A.; Matsumoto, S.; Ito, M. Taxonomy of the reticulate Vandenboschia radicans complex (Hymenophyllaceae) in Japan. Acta Phytotaxon. Geobot. 2009, 60, 26–40. [Google Scholar]
- Lin, Y.X.; Viane, R. Aspleniaceae; Wu, Z.Y., Raven, P.H., Hong, D.Y., Eds.; Flora of China Vol. 2–3 (Pteridophytes); Science Press: Beijing, China; Missouri Botanical Garden Press: St. Louis, MO, USA, 2013; pp. 267–316. [Google Scholar]
- PPGI. A community-derived classification for extant lycophytes and ferns. J. Syst. Evol. 2016, 54, 563–603. [Google Scholar] [CrossRef]
- Schneider, H.; Liu, H.M.; Chang, Y.F.; Ohlsen, D.; Perrie, L.R.; Shepherd, L.; Kessler, M.; Karger, D.N.; Hennequin, S.; Marquardt, J. Neo-and Paleopolyploidy contribute to the species diversity of Asplenium—The most species-rich genus of ferns. J. Syst. Evol. 2017, 55, 353–364. [Google Scholar] [CrossRef] [Green Version]
- Trewick, S.; Morgan-Richards, M.; Russell, S.; Henderson, S.; Rumsey, F.; Pinter, I.; Barrett, J.; Gibby, M.; Vogel, J. Polyploidy, phylogeography and Pleistocene refugia of the rockfern Asplenium ceterach: Evidence from chloroplast DNA. Mol. Ecol. 2002, 11, 2003–2012. [Google Scholar] [CrossRef]
- Chang, Y.; Li, J.; Lu, S.; Schneider, H. Species diversity and reticulate evolution in the Asplenium normale complex (Aspleniaceae) in China and adjacent areas. Taxon 2013, 62, 673–687. [Google Scholar] [CrossRef]
- Yatabe, Y.; Shinohara, W.; Matsumoto, S.; Murakami, N. Patterns of hybrid formation among cryptic species of bird-nest fern, Asplenium nidus complex (Aspleniaceae), in West Malesia. Bot. J. Linn. Soc. 2009, 160, 42–63. [Google Scholar] [CrossRef] [Green Version]
- Ichihara, Y. Ploidy level and reproductive mode of Asplenium capillipes (Aspleniaceae). J. Phytogeogr. Taxon. 2021, 69, 249–253. [Google Scholar] [CrossRef]
- Fraser-Jenkins, C.R. Taxonomic Revision of Three Hundred Indian Subcontinental Pteridophytes: With a Revised Census List; a New Picture of Fern-Taxonomy and Nomenclature in the Indian Subcontinent; Bishen Singh Mahendra Pal Singh: Dehradun, India, 2008. [Google Scholar]
- Ebihara, A. The Standard of Ferns and Lycophytes in Japan, Vol. 1; Gakken Publishers: Tokyo, Japan, 2016. [Google Scholar]
- BIR, S. Biosystematic studies on the asplenioid and athyrioid genera from the Himalayas: Results of chromosomal analysis on population basis. Indian Fern J. 1994, 11, 82–88. [Google Scholar]
- Walker, T. The cytogenetics of ferns. In The Experimental Biology of Ferns; Dyer, A.F., Ed.; Academic Press: London, UK; New York, NY, USA, 1979; pp. 87–132. [Google Scholar]
- Döpp, W. Die Apogamie bei Aspidium remotum Al. Br. Planta 1932, 17, 86–152. [Google Scholar] [CrossRef]
- Manton, I. Problems of Cytology and Evolution in the Pteridophyta; Cambridge University Press: London, UK, 1950. [Google Scholar]
- Braithwaite, A. A new type of apogamy in ferns. New Phytol. 1964, 63, 293–305. [Google Scholar] [CrossRef]
- Knobloch, I.W. A Preliminary Review of Spore Number and Apogamy within the Genus Cheilanthes. Am. Fern J. 1966, 56, 163–167. [Google Scholar] [CrossRef]
- Lin, S.-J.; Kato, M.; Iwatsuki, K. Sporogenesis, reproductive mode, and cytotaxonomy of some species of Sphenomeris, Lindsaea, and Tapeinidium (Lindsaeaceae). Am. Fern J. 1990, 80, 97–109. [Google Scholar] [CrossRef]
- Gastony, G.J. Spore morphology in the Dicksoniaceae. I. The genera Cystodium, Thyrsopteris, and Culcita. Am. J. Bot. 1981, 68, 808–819. [Google Scholar] [CrossRef]
- Huang, Y.-M.; Chiou, W.-L.; Lee, P.-H. Morphology of the gametophytes and young sporophytes of Cyatheaceae native to Taiwan. TAIWANIA-TAIPEI- 2001, 46, 274–283. [Google Scholar]
- Regalado, L.; Sánchez, C. Spore morphology as a taxonomic tool in the delimitation of three Asplenium L. species complexes (Aspleniaceae: Pteridophyta) in Cuba. Grana 2002, 41, 107–113. [Google Scholar] [CrossRef]
- Morzenti, V.M. Morphological and Cytological Data on Southeastern United States Species of the Asplenium heterochroum-resiliens Complex. Am. Fern J. 1966, 56, 167–177. [Google Scholar] [CrossRef]
- Mehra, P.N.; Bir, S.S. Cytological Observations on <i>Asplenium cheilosorum</i> Kunze. Cytologia 1960, 25, 17–27. [Google Scholar] [CrossRef] [Green Version]
- Mitui, K. Chromosome studies on Japanese ferns (4). J. Jap. Bot. 1970, 45, 84–90. [Google Scholar]
- De Groot, G.A.; During, H.J.; Maas, J.W.; Schneider, H.; Vogel, J.C.; Erkens, R.H. Use of rbcL and trnL-F as a two-locus DNA barcode for identification of NW-European ferns: An ecological perspective. PLoS ONE 2011, 6, e16371. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schuettpelz, E.; Pryer, K.M. Fern phylogeny inferred from 400 leptosporangiate species and three plastid genes. Taxon 2007, 56, 1037–1050. [Google Scholar]
- Hori, K.; Okuyama, Y.; Watano, Y.; Murakami, N. Recurrent hybridization without homoeologous chromosome paring in the Dryopteris varia complex (Dryopteridaceae). Chromosome Bot. 2018, 13, 9–24. [Google Scholar] [CrossRef] [Green Version]
- Schuettpelz, E.; Grusz, A.L.; Windham, M.D.; Pryer, K.M. The utility of nuclear gapCp in resolving polyploid fern origins. Syst. Bot. 2008, 33, 621–629. [Google Scholar] [CrossRef]
- Ishikawa, H.; Watano, Y.; Kano, K.; Ito, M.; Kurita, S. Development of primer sets for PCR amplification of the PgiC gene in ferns. J. Plant Res. 2002, 115, 0065–0070. [Google Scholar] [CrossRef]
- Kearse, M.; Moir, R.; Wilson, A.; Stones-Havas, S.; Cheung, M.; Sturrock, S.; Buxton, S.; Cooper, A.; Markowitz, S.; Duran, C.; et al. Geneious Basic: An integrated and extendable desktop software platform for the organization and analysis of sequence data. Bioinformatics 2012, 28, 1647–1649. [Google Scholar] [CrossRef] [Green Version]
- Katoh, K.; Misawa, K.; Kuma, K.i.; Miyata, T. MAFFT: A novel method for rapid multiple sequence alignment based on fast Fourier transform. Nucleic Acids Res. 2002, 30, 3059–3066. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Edgar, R.C. MUSCLE: A multiple sequence alignment method with reduced time and space complexity. BMC Bioinform. 2004, 5, 113. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kalyaanamoorthy, S.; Minh, B.Q.; Wong, T.K.; Von Haeseler, A.; Jermiin, L.S. ModelFinder: Fast model selection for accurate phylogenetic estimates. Nat. Methods 2017, 14, 587–589. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nguyen, L.-T.; Schmidt, H.A.; Von Haeseler, A.; Minh, B.Q. IQ-TREE: A fast and effective stochastic algorithm for estimating maximum-likelihood phylogenies. Mol. Biol. Evol. 2015, 32, 268–274. [Google Scholar] [CrossRef]
- Hoang, D.T.; Chernomor, O.; Von Haeseler, A.; Minh, B.Q.; Vinh, L.S. UFBoot2: Improving the ultrafast bootstrap approximation. Mol. Biol. Evol. 2018, 35, 518–522. [Google Scholar] [CrossRef]
- Valkonen, J.; Watanabe, K.; Pehu, E. Analysis of correlation between nuclear DNA content, chromosome number, and flowering capacity of asymmetric somatic hybrids of diploid Solanum brevidens and (di) haploid S. tuberosum. Jpn. J. Genet. 1994, 69, 525–536. [Google Scholar] [CrossRef]
Figure 1.
Maximum likelihood phylogeny based on plastid rbcL (A) and three nuclear markers, pgiC (B), AK1 (C), and gapCp (D) of Asplenium species and related species. Asplenium sp. is marked in red (Collection Site A) and blue (Collection Site B) boxes. Maximum likelihood bootstrap values are shown above the branches. The scale bar indicates a branch length corresponding to substitutions per site.
Figure 1.
Maximum likelihood phylogeny based on plastid rbcL (A) and three nuclear markers, pgiC (B), AK1 (C), and gapCp (D) of Asplenium species and related species. Asplenium sp. is marked in red (Collection Site A) and blue (Collection Site B) boxes. Maximum likelihood bootstrap values are shown above the branches. The scale bar indicates a branch length corresponding to substitutions per site.

Figure 2.
The mitotic chromosomes of Asplenium sp., 2n = 144. Microscopic photo. Scale bar = 10 μm.

Figure 3.
Results of genome size measurement using a flow cytometer. (A) Asplenium tenuicaule, (B) Asplenium sp.
Figure 3.
Results of genome size measurement using a flow cytometer. (A) Asplenium tenuicaule, (B) Asplenium sp.

Figure 4.
Asplenium pseudocapillipes sp. nov. (A,B) Habitat. (C) Plant. (D) Rhizome scale. (E) Thirty-two spores per sporangium.
Figure 4.
Asplenium pseudocapillipes sp. nov. (A,B) Habitat. (C) Plant. (D) Rhizome scale. (E) Thirty-two spores per sporangium.

Figure 5.
Two scenarios for the origin of A. pseudocapillipes. (A) Direct allopolyploidization via diploid hybrid. (B) Allotetraploidization via triploid bridge. Boxes represent each taxon and circles indicate gametes. Solid and dashed lines indicate known and putative taxa or gametes, respectively. The colors of the lines indicate whether it is native to Korea: black, native; gray, not native or unreported.
Figure 5.
Two scenarios for the origin of A. pseudocapillipes. (A) Direct allopolyploidization via diploid hybrid. (B) Allotetraploidization via triploid bridge. Boxes represent each taxon and circles indicate gametes. Solid and dashed lines indicate known and putative taxa or gametes, respectively. The colors of the lines indicate whether it is native to Korea: black, native; gray, not native or unreported.

Figure 6.
Collection sites for Asplenium sp. A: Taebaek-si. B: Jeongsun-gun.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
A comparison of A. pseudocapillipes and two parental species. Measurements of A. capillipes and A. tenuicaule were referred to in the Flora of China [18]. Asterisk (*) is a measurement of the spore length according to Ichihara [24].
| A. capillipes | A. pseudocapillipes | A. tenuicaule | |
|---|---|---|---|
| Plant height (cm) | 3–8 | 3–9 | 6–15 |
| Stipe (cm) | 0.3–2.5 | 0.5–3 | 1.5–5 |
| Lamina (cm) | 1.5–6 × 0.5–2.5 | 1.5–6 × 0.7–2.5 | 4–9 × 1.2–3 |
| Number of pinnae | 5–7 | 4–9 | 7–10 |
| Spore length (μm) | 38–42 45.7 * | 46–59 | 28–32 29.1 * |
| Ploidy | 2×, 4× | 4× | 2× |
| Number of spores per sporangium | 32 | 32 | 64 |
| Gemma on rachis | Present | Absent | Absent |
Table 2.
List of plant materials used for DNA analysis in this study.
| Voucher | Locality | Accession | |||
|---|---|---|---|---|---|
| rbcL | AK1 | gapCp | pgiC | ||
| Asplenium sp. | |||||
| CBNU2021-0084-A | Korea: Gangwon-do, Taebaek-si | OP114589 | OP114405 P114406 | OP153882 P153883 | OP114578 P114579 |
| CBNU2020-0171-A | Korea: Gangwon-do, Jeongsun-gun | OP114590 | OP114407 P114408 | OP153884 P153885 | OP114580 P114581 |
| Asplenium tenuicaule | |||||
| CBNU2020-0096-E | Korea: Jeollabuk-do, Suncheon-si | OP114586 | OP114403 | OP153880 | OP114576 |
| CBNU2020-0157-D | Korea: Jeju Island, Jeju-si | OP114587 | OP114402 | OP153879 | OP114575 |
| CBNU2021-0086-D | Korea: Gangwon-do, Taebaek-si | OP114585 | OP114401 | OP153878 | OP114574 |
| Asplenium ruprechtii | |||||
| CBNU2020-0180-A | Korea: Seoul, Gangbuk-gu | OP114588 | OP114404 | OP153881 | OP114577 |
| Asplenium tripteropus | |||||
| CBNU2020-0028-A | Korea: Jeju Island, Jeju-si | OP114584 | OP114400 | OP153877 | OP114573 |
| Athyrium yokoscense(outgroup) | |||||
| CBNU2021-0104-A | Korea: Jeollanam-do, Gwangju-si | OP114583 | OP114399 | OP153876 | OP114572 |
| Deparia pterorachis (outgroup) | |||||
| CBNU2021-0017-A | Korea: Gyeongsangbuk-do, Ulleung Island | OP114582 | OP114398 | OP153875 | OP114571 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Park, S.H.; Kim, J.S.; Kim, H.T. Asplenium pseudocapillipes (Aspleniaceae), a New Fern Species from South Korea. Plants 2022, 11, 3089. https://doi.org/10.3390/plants11223089
AMA Style
Park SH, Kim JS, Kim HT. Asplenium pseudocapillipes (Aspleniaceae), a New Fern Species from South Korea. Plants. 2022; 11(22):3089. https://doi.org/10.3390/plants11223089
Chicago/Turabian StylePark, Sang Hee, Jung Sung Kim, and Hyoung Tae Kim. 2022. "Asplenium pseudocapillipes (Aspleniaceae), a New Fern Species from South Korea" Plants 11, no. 22: 3089. https://doi.org/10.3390/plants11223089
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.