
Two Decades of Desiccation Biology: A Systematic Review of the Best Studied Angiosperm Resurrection Plants


1
Department of Molecular and Cell Biology, University of Cape Town, Rondebosch, Cape Town 7701, South Africa
2
Department of Horticulture, Michigan State University, East Lansing, MI 48824, USA
3
Plant Resiliency Institute, Michigan State University, East Lansing, MI 48824, USA
*
Author to whom correspondence should be addressed.
Plants 2021, 10(12), 2784; https://doi.org/10.3390/plants10122784
Submission received: 19 October 2021
/
Revised: 6 December 2021
/
Accepted: 6 December 2021
/
Published: 16 December 2021
(This article belongs to the Special Issue Drought and Desiccation Tolerance in Plants)
Abstract
:Resurrection plants have an extraordinary ability to survive extreme water loss but still revive full metabolic activity when rehydrated. These plants are useful models to understand the complex biology of vegetative desiccation tolerance. Despite extensive studies of resurrection plants, many details underlying the mechanisms of desiccation tolerance remain unexplored. To summarize the progress in resurrection plant research and identify unexplored questions, we conducted a systematic review of 15 model angiosperm resurrection plants. This systematic review provides an overview of publication trends on resurrection plants, the geographical distribution of species and studies, and the methodology used. Using the Preferred Reporting Items for Systematic reviews and Meta–Analyses protocol we surveyed all publications on resurrection plants from 2000 and 2020. This yielded 185 empirical articles that matched our selection criteria. The most investigated plants were Craterostigma plantagineum (17.5%), Haberlea rhodopensis (13.7%), Xerophyta viscosa (reclassified as X. schlechteri) (11.9%), Myrothamnus flabellifolia (8.5%), and Boea hygrometrica (8.1%), with all other species accounting for less than 8% of publications. The majority of studies have been conducted in South Africa, Bulgaria, Germany, and China, but there are contributions from across the globe. Most studies were led by researchers working within the native range of the focal species, but some international and collaborative studies were also identified. The number of annual publications fluctuated, with a large but temporary increase in 2008. Many studies have employed physiological and transcriptomic methodologies to investigate the leaves of resurrection plants, but there was a paucity of studies on roots and only one metagenomic study was recovered. Based on these findings we suggest that future research focuses on resurrection plant roots and microbiome interactions to explore microbial communities associated with these plants, and their role in vegetative desiccation tolerance.
1. Introduction
Even though water is essential to life, plants are often faced with shortages of this valuable resource due to their sessile nature. As a result, many have evolved sophisticated strategies for resisting, avoiding or tolerating water shortages [1,2]. One of the most successful adaptations to such extreme drought is desiccation tolerance—the ability to survive water loss to 10% relative water content (RWC), equivalent to 0.1 g H2O/g dry weight, and revive full metabolism when rehydrated [3]. Such plants are commonly called resurrection plants [4]. Desiccation tolerance is common in seeds, spores, and pollen, but very rare in vegetative tissues of plant, occurring in only ~240 angiosperms. Interestingly, resurrection plants are extremely diverse, representing at least 10 families [5] across both monocotyledon and dicotyledon lineages.
Resurrection plants have received growing research attention in the past 20 years and a handful of species have emerged as models for understanding desiccation tolerance. Among these are the monocot resurrection plants Eragrostis nindensis, Oropetium thomaeum, Sporobolus stapfianus, Tripogon loliiformis, Xerophyta humilis and Xerophyta viscosa* (multiple populations were later reclassified as X. schlechteri) and the dicots Boea hygrometrica, Craterostigma plantagineum, Craterostigma pumilum, Craterostigma wilmsii, Haberlea rhodopensis, Lindernia brevidens, Myrothamnus flabellifolia, Ramonda serbica and Ramonda nathaliae. These species are distributed across the globe, with representatives found in both the Southern and Northern hemispheres, but the highest density of resurrection plants occurs in arid tropical and subtropical regions in Africa, South America and Australia. Fewer resurrection plants are found in the Northern hemisphere, but B. hygrometrica and Paraboea rufescens occur in Asia, and H. rhodopensis and Ramonda species are endemic to Europe [6,7,8]. The ability to tolerate desiccation has enabled resurrection plants to thrive in extremely arid microclimatic conditions where other plants perish. Resurrection plants grow predominantly in sites with shallow rocky soil, high temperatures, and limited rainfall [5,9].
Resurrection plants have evolved protective mechanisms that allow them to cope with environmental stress, including a robust antioxidant defense system, sophisticated gene expression programs in which late embryogenesis abundant, heat shock proteins, and other stress responsive genes are transcribed and where necessary translated, and subtle metabolic modulations involving numerous phytohormones and phytochemicals [3,10,11,12,13]. Despite the intricacy of desiccation tolerance mechanisms in plants, scientists have made significant improvements in developing an understanding these processes.
Over the past two decades “omics” methodologies have been increasingly applied to resurrection plants to acquire insights into the biochemical processes and molecular mechanisms of desiccation tolerance. Multiple genomic, transcriptomic, proteomic, metabolomic, and physiological/biochemical studies of desiccation tolerant plants have been published in recent years. The advances in “omics” techniques enable exploration of new genes, transcripts, metabolites, proteins, and microbes that contribute to desiccation tolerance. Recently, genomic and transcriptomic analyses of B. hygrometrica, C. plantagineum, E. nindensis, H. rhodopensis, L. brevidens, O. thomaeum, T. loliiformis, and X. schlechteri, have identified gene families and transcripts involved in desiccation responses [14,15,16,17,18,19,20,21]. Metabolomic analysis of C. plantagineum, L. brevidens, M. flabellifolia, and X. schlechteri have provided insight into the central role of sugars (sucrose in particular), selected amino and organic acids, and phenolic antioxidants [22,23,24]. However, the bulk of these studies have been conducted on leaf tissues only, with the role of the root being largely ignored. Furthermore, there are limited studies addressing the microbial communities associated with resurrection plants, despite growing awareness of the importance of plant microbiota in plant host performance. Given that microbes have the potential to improve resilience to numerous biotic and abiotic stresses [25,26], it is likely that these interactions play a prominent role in desiccation tolerance.
This review describes the changing landscape of resurrection plant research over the past 20 years and identifies under–explored areas of research. We systematically summarized research efforts on resurrection plants to identify the 15 most studied resurrection plants–based on publication number—and compiled all genomic, transcriptomic, proteomic, metabolomic, metagenomic and physiological studies on these species from 2000 to 2020. In total, we summarized 185 research studies on resurrection plants to address the following questions: (i) what are the best studied resurrection plants? (ii) where are these species native to and how does that relate to where have they been studied? (iii) what types of methodologies have been applied to each species? and (iv) what are under–explored avenues of research in resurrection plants? To the best of our knowledge, this serves as the first systematic review conducted on such a diverse set of resurrection plants.
2. Materials and Methods
2.1. Literature Search and Selection Criteria
We used a multi–step protocol called Preferred Reporting Items for Systematic reviews and Meta–Analyses (PRISMA) to collate publications for this systematic review [27,28]. A literature search was conducted using EBSCOhost, Scopus, google scholar, ProQuest, PubMed, SciFinder and Europe PubMed Central (Europe PMC). The search was conducted in January 2021 using wildcards including “resurrection plants” or “desiccation tolerant plants”, “desiccation stress” or “water deficit conditions” and “dehydration and rehydration”. There were no filters applied in the search. Additional research articles that matched our search criteria were obtained from reference lists of the published reviews and original articles. This literature search generated a total of 6522 studies and all citations were exported to EndNote 20.1 for macOS.
A screening process was conducted in the EndNote software by manually evaluating article titles and abstracts. Initial screening was performed to discard duplicates, reviews, mini–reviews, editorials, commentaries, books, book chapters, theses, dissertations, opinions, conference abstracts, presentations, meta–analyses, protocols, manuals, notes, and news. Next, selection and exclusion criteria were applied to the title and abstract of each article. Studies were excluded based on (i) research articles dated before the year 2000; (ii) scientific journals without impact factor; (iii) studies of resurrection plants for medicinal use; (iv) studies that did not focus on desiccation tolerance and defense mechanisms against abiotic stress; and (v) resurrection plants under the divisions of Bryophyta, Marchantiophyta, and Pteridophyta. The selection criteria for inclusion of scientific articles included (i) original full–text articles published in English from 2000 to 2020; (ii) studies involving angiosperm resurrection plants regardless of the location or country; (iii) research articles that focused on desiccation tolerance mechanisms and (iv) studies based on genomics, transcriptomics, proteomics, metabolomics, metagenomics and physiology. All records of the literature search and the number of included full–text articles were retained in accordance with the PRISMA framework (Figure 1).
2.2. Literature Analysis and Data Acquisition
From this set of literature, we summarized overall trends on the number and type of publications on resurrection plants from 2000 to 2020, including the number of studies conducted on each species, the country where these studies were conducted relative to the native range of the species, and the types of studies performed—with a particular focus on “omics” methodologies. Initially, we compared publication trends over time by computing the number of studies published annually from 2000 to 2020 (see Supplementary Table S1 for a detailed list of full-text surveyed articles). Next, we computed the total number of studies for each focal species. It is noteworthy that some populations of Xerophyta viscosa have recently been reclassified as X. schlechteri. In addition, the majority of papers (reference: 2, 10, 16, 18, 50, 77 and 85) on X. schlechteri refer to it as Xerophyta viscosa. This has been taken into account in our dataset. To understand the geographical distribution of focal species relative to research efforts, we categorized studies into three groups: (i) native—conducted by researchers working within the native range of the focal species, (ii) collaborative—conducted by researchers working both within and beyond the native range of the focal species, and (iii) international—conducted by researchers working outside of the focal species native range. To understand research methodologies, we classified studies into six categories: (i) studies involving analysis of genes, their function and expression using different techniques were assigned to genomic studies; (ii) articles that were investigating ribonucleic acid (RNA) transcripts were classified as transcriptomic studies; (iii) articles that reported the role of proteins were categorized as proteomic studies; (iv) studies that involved profiling and quantification of primary and secondary metabolites were captured as metabolomic studies; (v) studies involving the analysis of microbes associated with resurrection plants were classified as metagenomics; and (vi) physiological studies included studies looking at a range of responses from RWC, dry mass, germination, stomatal conductance, photosynthesis rate, antioxidant enzyme activity, to ultrastructural image–based technologies.
Data were extracted and arranged in Microsoft Excel 2019 and analyzed in R version 1.2.5033 (R Studio Inc., Boston, MA, USA, 2019). We computed the number of studies conducted on each species annually, if they were native, collaborative or international study, and what the methodologies were. The R Packages gplots, ggplot2 and plotly were used for data visualization.
3. Results
After a rigorous screening and filtering, the literature search yielded 185 articles (Figure 1). We used these articles to identify the best studied resurrection plant species, to summarize the geographical distribution of focal species relative to research efforts, and to quantify the research methodologies used. A total of 15 angiosperm resurrection plants were selected for inclusion, namely B. hygrometrica, C. plantagineum, C. pumilum, C. wilmsii, E. nindensis, H. rhodopensis, L. brevidens, M. flabellifolia, O. thomaeum, R. nathaliae, R. serbica, S. stapfianus, T. loliiformis, X. humilis, and X. schlechteri.
3.1. Well Studied Angiosperm Resurrection Plants
To identify the best studied resurrection plants, we computed the total number of studies focusing on each species. Some articles analyzed more than one resurrection species and these studies were counted for each plant species. Therefore, the sum of all studies reported on each plant (211) exceeds the total number of studies (185). The largest proportion (17.5%, 37/211) of studies were conducted on C. plantagineum. Following that, 13.7% of studies were conducted on H. rhodopensis, 11.9% on X. schlechteri, 8.5% on M. flabellifolia, 8.1% on B. hygrometrica, 7.1% on X. humilis, 7.6% on R. serbica, 6.2% on S. stapfianus, 4.7% on E. nindensis, 3.3% on L. brevidens, 2.8% on C. wilmsii, 2.4% on R. nathaliae, and T. loliiformis, and 1.9% on C. pumilum and O. thomaeum (see Figure 2).
3.2. Distribution of Resurrection Plants
Resurrection plants occur and have been studied across the globe, from Africa to Asia, Oceania, Europe, and North America (Table 1). Researchers working in South Africa have published more studies (56) on resurrection plants than any other nation. These studies investigated the focal species M. flabellifolia (12), Craterostigma spp. (3), Xerophyta spp. (39) and multiple species within Poaceae (10). Bulgaria was the country with the second most studies—primarily focused on H. rhodopensis (24) and R. serbica (1), followed by China with 17 studies of B. hygrometrica (Figure 3).
Most studies (55.1%, 102/185) were conducted by teams working within the native range of the focal species (Figure 2). Six countries explored their native resurrection plants, namely Australia, Bulgaria, China, Kosovo, Serbia, and South Africa. Other studies (30.8%, 57/185) were conducted by teams working outside the native range of the focal species. These international studies were distributed across the world, with major contributions from European countries, including Germany, Hungary and Italy (Figure 3). Other international studies were conducted in China, India, Kenya, and the United States of America (USA). Germany, in particular, studied many non–native resurrection species including Craterostigma spp. (35), L. brevidens (6), M. flabellifolia (2) and grasses in Poaceae (2). We also identified a number of collaborative studies (14%, 26/185) involving scientists working both within and beyond the native range of the focal species. Many of these involved participants from Asia, Europe, and USA who established collaborations with researchers in South Africa (Figure 3).
3.3. Publication Trends and Methodologies of Investigation
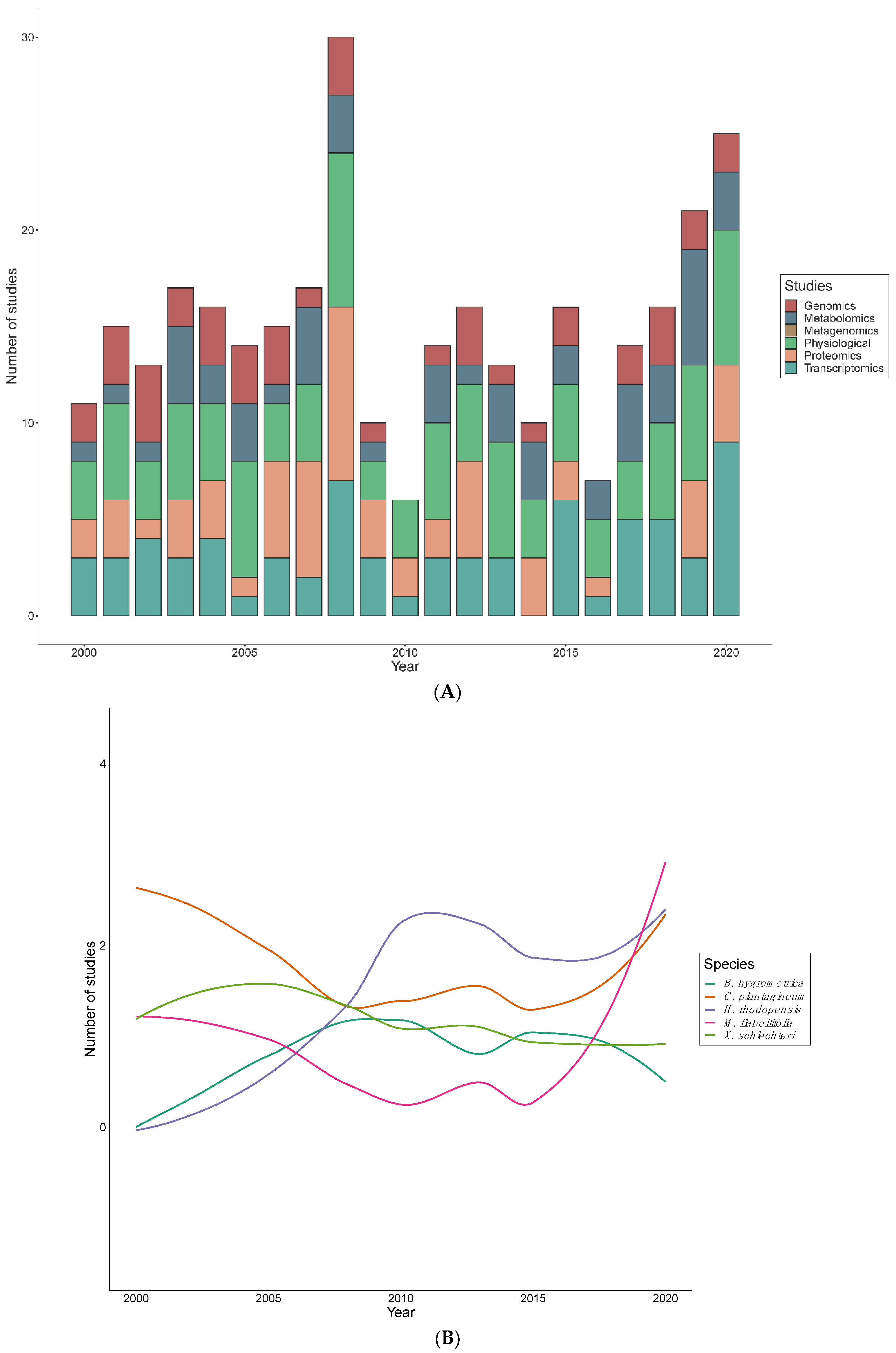
Publication rates have been relatively erratic, with 2008, and 2018–2020 showing the highest numbers of publications (Figure 4A). An average of seven articles were published per year, despite the decline in the years 2009, 2010 and 2016. In the past two decades, C. plantagineum was most intensively studied from 2000 and 2005, while the investigation of H. rhodopensis and M. flabellifolia took an upsurge in 2009 and 2019, respectively (Figure 4B). In contrast, X. schlechteri studies remained steady from the year 2000, but slightly decreased in 2015–2018. A wide range of high–throughput technologies have been employed in order to explore desiccation tolerance mechanisms in resurrection plants (Table 2). These methodologies have been applied to individual species in differing proportions (Figure 4A). Physiological studies were the most common approach to studying resurrection plants while genomic studies are the least reported. Proteomic and transcriptomic studies followed a comparable publication pattern, however, there was a significant increase of transcriptomics in 2020. Metabolomic studies have fluctuated between 2000 to 2020, however, a significant reduction occurred in 2009, 2010, and 2016. Out of 185 records, there was only one metagenome study recovered–conducted on Ramonda species.
4. Discussion
Resurrection plants are a distinctive group of species that could provide scientists with key insights into the mechanisms of extreme stress tolerance, which may ultimately be leveraged to improve tolerance in drought–sensitive crops. It is not surprising that resurrection plants have received increasing research attention over the past years as appreciation for their phenomenal defense mechanisms has become more widespread. Our study was based on the analysis of 185 empirical studies of angiosperm resurrection plants published over the past two decades. Our systematic review identifies the best studied resurrection plants, the geographical distribution of species and research efforts across the world, and changes in the methodologies used for investigation.
Our findings revealed that out of the 15 model species, C. plantagineum was the most studied, with 17.5% of all publications in the past 20 years devoted to this species. The publication trends of C. plantagineum were at the maximum between 2000 and 2005. However, in some studies C. plantagineum was explored concurrently with other resurrection plants. For instance, Moore et al. [29] simultaneously investigated the functional role of arabinose polymers as plasticizers and maintaining flexibility in the cell wall of C. plantagineum, M. flabellifolia, E. nindensis and Xerophyta spp. These multi–species approaches enable researchers to identify conserved aspects of desiccation tolerance across disparate clades. Surprisingly, 95% studies of C. plantagineum were international studies, with German researchers leading the work on this species, although it is native to Southern Africa. Another resurrection species, H. rhodopensis, was extensively studied with 13.7% of publications addressing desiccation tolerance mechanisms in this species. The desiccation tolerance features of H. rhodopensis have been recently reviewed by Liu et al. [12]. Interestingly, H. rhodopensis was also frequently investigated alongside other species. For example, a study by Vassileva et al. [62] assessed leaf micromorphology of both H. rhodopensis and R. serbica, and both species are native to Bulgaria. Xerophyta spp. were also widely explored, constituting 19% of all studies. Although C. pumilum, L. brevidens, O. thomaeum, R. nathaliae, and T. loliiformis have been classified as best studied resurrection species by others, our findings showed that these were among the least studied species in our dataset. This is partially explained by the fact that some of the work on O. thomaeum, is not related to desiccation tolerance mechanisms and was therefore not included in the current analyses [31,63]. Despite the relatively low number of studies on O. thomaeum, there is a chromosome-level genome assembly and associated RNA-seq data [16,20] for the species. This is in contrast to some of the other species, such as R. nathaliae, for which there are no genomic resources available. We suggest that more research is required to understand how these species respond to desiccation and to identify mechanistic deviations relative to other angiosperms. The families of Poaceae and Cyperaceae contain many of the known desiccation tolerant species [64], however, within those families only few species have been extensively studied. Despite some similarities among angiosperm resurrection plants, substantial variations in mechanisms of tolerance have been identified [5,65], including differences in the production of metabolites, expression of genes, synthesis of proteins and physical and ultrastructural features. Thus, there is still room to explore the intricate and diverse mechanisms of stress tolerance exhibited by these species.
We also investigated the distribution of resurrection plant research across the world. Resurrection plants are common in arid tropical and subtropical regions, occurring mostly on mountains with exposed and rocky surfaces [9,66]. Our results showed that most studies of resurrection plants were conducted within the native range of the species, likely because access to plant material is much easier when working within the native range of the focal species. In particular, most studies were executed in South Africa which is likely due to the remarkable diversity of resurrection plants native to Southern Africa. In fact, seven out of the fifteen best studied species are native to Southern Africa, which is known as an epicenter of diversity of resurrection plants [64,65,67,68]. Despite the species richness in Africa, resurrection plant research still lags behind in other African countries, possibly due to lack of resources or funding. Outside of Africa, Bulgaria has been a major contributor to resurrection plant research with many studies of H. rhodopensis and R. serbica, both of which are native to the Balkan and Rhodope mountains of Eastern Europe [6,69]. Similarly, 99% of the studies conducted on B. hygrometrica were carried out within the species native range of China. It is noteworthy that limited collaborative studies were discovered. A number of the resurrection plants native to the Global South were studied in European countries without instigating collaboration. This could be attributed to the narrow distribution of resurrection plants in the Global North as well as differences in funding and resources available for research in the Global North relative to the Global South [70]. Despite these imbalances, South Africa remains a leader in the research of angiosperm resurrection plants. That being said, multiple collaborative efforts were identified linking researchers in South Africa to international scientists in Asia, Europe, and North America. These collaborative efforts may facilitate the exploration of desiccation tolerance mechanisms and minimize costs for those residing in indigent countries.
High–throughput methodologies have been increasingly applied to resurrection plants and provide comprehensive insight into the mechanisms of desiccation tolerance. We found that a greater proportion of studies focused on physiological and transcriptomic techniques, relative to other methodologies. The physiological studies correlate with average research publications per year. This could be attributed to the fact that physiological studies typically do not involve advanced technologies compared to other methodologies and are therefore easier to initiate. For instance, light microscope, scanning electron microscope (SEM), and transmission electron microscopes were used to examine physiological [68,71] and ultrastructural [24,65,72] aspects of resurrection plants.
In the past years, scientists have explored resurrection plant genomes, yet only 5 of the 15 best studied resurrection plants have a publicly available whole genome assembly [73]. Although genome sequencing of desiccation tolerant plants has sought to identify a “footprint” of vegetative desiccation tolerance in these species [3,74], such signatures have not been forthcoming. There have been some genetic studies of Ramonda spp. [75], however, the focus was not directed to the functional role of genes under desiccation conditions. There are various emerging genetic technologies such as genome-editing (clustered regularly interspaced short palindromic repeats) applied for gene identification in microorganisms and plant species, but this approach is rarely employed in resurrection plant research. However, Costa et al. [76] suggested a futuristic approach of further studying desiccation tolerance genes by breeding plants with improved tolerance to drought. Nevertheless, Hilhorst et al. [11] postulated that more genomic research is anticipated in the near future.
The rapid increase in omics research in recent years correlates with the advances in technologies. It is worth noting that high–throughput technologies are expensive and mostly performed in well–developed countries, while lower income countries lag behind. Our analyses show that most expensive high–throughput technologies were applied in studies conducted in the USA, Australia, China, and European countries. For instance, all genome sequencing studies have emanated from the Global North, with the exception of China–a notable outlier relative to other countries in the Global South. Transcriptome studies were also predominately conducted in the Global North including those on B. hygrometrica [21], C. plantagineum [77], L. brevidens [20], E. nindensis [19], O. thomaeum [16] and S. stapfianus [34]. Similarly, proteome studies were mostly performed in the Global North on C. plantagineum [77], R. serbica and B. hygrometrica [78], although two papers on X. viscosa [51,79] have come from South Africa. Metabolomic studies, on the other hand show equal distribution been the Global North, on S. stapfianus [34], C. plantagineum [77], H. rhodopensis [80] and B. hygrometrica [81], and the Global South on, M. flabellifolia [82] and X. schlechteri [24]. In addition, the execution of these high–throughput techniques was also noted in collaborative studies between well resourced (mostly Northern) and under resourced (mostly Southern) countries, such as those by Plancot et al. [55], Costa et al. [16], Vidović et al. [83] and Pardo et al. [19]. These patterns further reinforce the notion that economic barriers limit participation in omics studies by teams working exclusively in the Global South. Omics research is underrepresented in Africa, not only in plant biology, but also in the biomedical research [70] and these challenges are mainly associated with socio–economic factors. In this regard, we urge researchers to develop more collaborations between countries with a rich diversity of desiccation tolerant plants and countries with economic resources. Such collaborations will not only aid in understanding desiccation tolerance and facilitating the biotechnological roll out of drought tolerant crops, but will also expand participation and bring diverse expertise to resurrection plant research. This is critical for food security especially in economically depressed African nations, where 95% of agriculture relies on rainfall and where, due to climate change, droughts are predicted to have become so severe that by 2050 all conventional farming practices will be abandoned [84,85].
Collectively, molecular studies have revealed that there are core transcripts, proteins and metabolites produced by different resurrection plants. However, there are also considerable mechanistic differences among genera and species [12,74]. Thus, it is imperative to continue in depth systems studies on as many resurrection plants as possible to fully understand the spectrum of ‘’tools” used, and the manner in use thereof, in achieving desiccation tolerance. With this insight, the use of individual resurrection species as models for specific crops becomes more feasible. Interestingly, some individual species show natural variation and plasticity in desiccation tolerance. Bentley et al. [82] reported differences in the metabolites of M. flabellifolia from different geographical regions. This could be due to numerous environmental factors including the soil/bedrock type and respective microbial interactions, light intensities, temperatures, rainfall and biotic stresses experienced in different regions. The nature of such fine tuning could explain “leeways” in vegetative desiccation tolerance and is of interest to the medical and cosmetic industries. M. flabellifolia itself has several medicinal [82] and cosmetic [86] applications. We suggest that investigation of a species across its geographic range could provide insight on the genetics, metabolites, microbes, and environmental factors that impact desiccation tolerance.
The role of roots in desiccation tolerance is largely unexplored. The few studies that report on roots include a physiological study on roots of X. schlechteri [87] and H. rhodopensis [88], one phenology and carbohydrate metabolism in C. plantagineum [89], one metabolomic and transcriptomic study in T. loliiformis [15] and one study on the significant role of phytohormones in dehydrated H. rhodopensis [90]. While diverse in nature, these studies confirm the central role of sucrose and antioxidants in desiccation tolerance, but also reveal stark differences in root longevity. Roots of C. plantagineum senesce after rehydration and new ones are generated [89]. In the other two species, roots are maintained, and indeed senescence is actively suppressed in T. lolliformis [15]. Numerous questions remain and we propose that root systems studies are sorely needed.
Similarly, there has been limited research published on the microbiome associated with resurrection plants. The only metagenomic study recovered from all 185 records in our analyses explored rhizospheric bacterial diversity associated with Ramonda species [91]. Rakic et al. [8] also investigated the role of mycorrhizal fungi associated with R. nathaliae roots in facilitating mineral stress associated with serpentine soils, but this was not directly related to their desiccation tolerance.
As indicated before, plant–microbiome interactions play an important role in alleviating environmental stress, improving nutrient uptake and facilitating plant growth [25]. Therefore, metagenomics studies aimed at the identification and characterization of microbiota inhabiting the rhizosphere and roots of resurrection plants could provide insight into the functional role of plant microbiomes and their influence on desiccation tolerance mechanisms. Further investigation should be conducted to understand the role of plant associated microbiota under desiccating conditions.
5. Conclusions and Future Research
Resurrection plants are excellent models for investigating plant responses to environmental stresses and adaptive mechanisms of water stress tolerance. Our analyses showed that research efforts have been skewed to C. plantagineum, H. rhodopensis, B. hygrometrica, and Xerophyta spp. We also identified a link between the geographical range of the focal resurrection plants and the national affiliations of the researchers studying them, with most resurrection species being studied within their native range. The integration of multi–omics studies provide new opportunities to understand desiccation tolerance mechanisms. Despite extensive research on the intricate mechanisms of vegetative desiccation tolerance, our results showed that the biggest gaps lie belowground, and more research is needed to understand root molecular physiology, metagenomics and plant microbe interactions.
This systematic study identifies knowledge gaps pertaining to resurrection plants and points towards socio–economic barriers impacting research outputs. We recommend that (i) despite the available literature on the well-studied angiosperm resurrection plants (O. thomaeum, S. stapfianus, T. loliiformis, X. humilis, C. pumilum, C. wilmsii, L. brevidens, R. serbica, and R. nathaliae) more research is still needed on these species; (ii) researchers establish collaborations across geographic and socio economic space to apply new methodologies to resurrection plants native to Global South; (iii) studies aim to identify the complex interplay between, and regulatory features associated with, the genome, transcriptome, proteome, metabolome and consequent physiological outplay; and. Within these, there are still big gaps in our knowledge. For example, there are no studies on epigenetic regulation of vegetative desiccation tolerance, although some hint that this plays a significant role (reviewed in [92]); (iv) that more focus should be given to studies aimed at exploring the belowground dynamics of desiccation tolerance.
Supplementary Materials
The following are available online at https://www.mdpi.com/article/10.3390/plants10122784/s1, Table S1. A list of articles used in this study as well as captured data.
Author Contributions
Conceptualization, S.M.T., R.A.M. and J.M.F.; methodology, S.M.T. and R.A.M.; literature search and extraction, S.M.T.; data curation, S.M.T.; visualization, S.M.T. and R.A.M.; writing—original draft preparation S.M.T.; writing—review and editing S.M.T., R.A.M. and J.M.F. All authors have read and agreed to the published version of the manuscript.
Funding
S.M.T. was funded by National Research Foundation Grant–holder’s scholarship and University of Cape Town Postgraduate funding, R.A.M. was supported by NSF Postdoctoral Research Fellowship in Biology (IOS-1906094) and by the Plant Resiliency Institute at Michigan State University and J.M.F. acknowledges funding from the Department of Science and Innovation, National Research Foundation South African Research Chair grant number 98406.
Data Availability Statement
Data used in this study are available in the Supplementary Materials.
Acknowledgments
We thank all authors who conducted the original/primary research that this review focused on. We express our gratitude to the reviewers and editor of this review article.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Farrant, J.M.; Vander Willigen, C.; Loffell, D.A.; Bartsch, S.; Whittaker, A. An investigation into the role of light during desiccation of three angiosperm resurrection plants. Plant Cell Environ. 2003, 26, 1275–1286. [Google Scholar] [CrossRef] [Green Version]
- Mundree, S.; Whittaker, A.; Thomson, J.; Farrant, J. An aldose reductase homolog from the resurrection plant Xerophyta viscosa Baker. Planta 2000, 211, 693–700. [Google Scholar] [CrossRef] [PubMed]
- Oliver, J.M.; Farrant, M.J.; Hilhorst, W.M.H.; Mundree, S.; Williams, B.; Bewley, D.J. Desiccation Tolerance: Avoiding Cellular Damage During Drying and Rehydration. Annu. Rev. Plant Biol. 2020, 71, 435–460. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gaff, D.F. Desiccation-tolerant flowering plants in southern Africa. Science 1971, 174, 1033–1034. [Google Scholar] [CrossRef] [PubMed]
- Marks, R.A.; Farrant, J.M.; Nicholas McLetchie, D.; Van Buren, R. Unexplored dimensions of variability in vegetative desiccation tolerance. Am. J. Bot. 2021, 108, 346–358. [Google Scholar] [CrossRef]
- Georgieva, K.; Szigeti, Z.; Sarvari, E.; Gaspar, L.; Maslenkova, L.; Peeva, V.; Peli, E.; Tuba, Z. Photosynthetic activity of homoiochlorophyllous desiccation tolerant plant Haberlea rhodopensis during dehydration and rehydration. Planta 2007, 225, 955–964. [Google Scholar] [CrossRef]
- Mitra, J.; Xu, G.; Wang, B.; Li, M.; Deng, X. Understanding desiccation tolerance using the resurrection plant Boea hygrometrica as a model system. Front. Plant Sci. 2013, 4, 446. [Google Scholar] [CrossRef] [Green Version]
- Rakic, T.; Ilijevic, K.; Lazarevic, M.; Grzetic, I.; Stevanovic, V.; Stevanovic, B. The resurrection flowering plant Ramonda nathaliae on serpentine soil–coping with extreme mineral element stress. Flora-Morphol. Distrib. Funct. Ecol. Plants 2013, 208, 618–625. [Google Scholar] [CrossRef]
- Porembski, S.; Barthlott, W. Granitic and gneissic outcrops (inselbergs) as centers of diversity for desiccation-tolerant vascular plants. Plant Ecol. 2000, 151, 19–28. [Google Scholar] [CrossRef]
- Farrant, J.M.; Cooper, K.; Hilgart, A.; Abdalla, K.O.; Bentley, J.; Thomson, J.A.; Dace, H.J.W.; Peton, N.; Mundree, S.G.; Rafudeen, M.S. A molecular physiological review of vegetative desiccation tolerance in the resurrection plant Xerophyta viscosa (Baker). Planta 2015, 242, 407–426. [Google Scholar] [CrossRef] [Green Version]
- Hilhorst, H.W.; Costa, M.-C.D.; Farrant, J.M. A footprint of plant desiccation tolerance. Does it exist? Mol. Plant 2018, 11, 1003–1005. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, J.; Moyankova, D.; Djilianov, D.; Deng, X. Common and Specific Mechanisms of Desiccation Tolerance in Two Gesneriaceae Resurrection Plants. Multiomics Evidences. Front. Plant Sci. 2019, 10, 1067. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- VanBuren, R.; Wai, C.M.; Zhang, Q.; Song, X.; Edger, P.P.; Bryant, D.; Michael, T.P.; Mockler, T.C.; Bartels, D. Seed desiccation mechanisms co-opted for vegetative desiccation in the resurrection grass Oropetium thomaeum. Plant Cell Environ. 2017, 40, 2292–2306. [Google Scholar] [CrossRef] [PubMed]
- Apostolova, E.; Gozmanova, M.; Nacheva, L.; Ivanova, Z.; Toneva, V.; Minkov, I.; Baev, V.; Yahubyan, G. MicroRNA profiling the resurrection plant Haberlea rhodopensis unveils essential regulators of survival under severe drought. Biol. Plant. 2020, 64, 541–550. [Google Scholar] [CrossRef]
- Asami, P.; Rupasinghe, T.; Moghaddam, L.; Njaci, I.; Roessner, U.; Mundree, S.; Williams, B. Roots of the Resurrection Plant Tripogon loliiformis Survive Desiccation Without the Activation of Autophagy Pathways by Maintaining Energy Reserves. Front. Plant Sci. 2019, 10, 459. [Google Scholar] [CrossRef]
- Costa, M.-C.D.; Artur, M.A.S.; Maia, J.; Jonkheer, E.; Derks, M.F.L.; Nijveen, H.; Williams, B.; Mundree, S.G.; Jiménez-Gómez, J.M.; Hesselink, T.; et al. A footprint of desiccation tolerance in the genome of Xerophyta viscosa. Nat. Plants 2017, 3, 17038. [Google Scholar] [CrossRef] [Green Version]
- Giarola, V.; Jung, N.U.; Singh, A.; Satpathy, P.; Bartels, D. Analysis of pcC13-62 promoters predicts a link between cis-element variations and desiccation tolerance in Linderniaceae. J. Exp. Bot. 2018, 69, 3773–3784. [Google Scholar] [CrossRef]
- Mbinda, W.; Dixelius, C.; Oduor, R. Induced Expression of Xerophyta viscosa XvSap1 Gene Enhances Drought Tolerance in Transgenic Sweet Potato. Front. Plant Sci. 2019, 10, 1119. [Google Scholar] [CrossRef] [Green Version]
- Pardo, J.; Man Wai, C.; Chay, H.; Madden, C.F.; Hilhorst, H.W.M.; Farrant, J.M.; VanBuren, R. Intertwined signatures of desiccation and drought tolerance in grasses. Proc. Natl. Acad. Sci. USA 2020, 117, 10079–10088. [Google Scholar] [CrossRef] [Green Version]
- VanBuren, R.; Bryant, D.; Edger, P.P.; Tang, H.; Burgess, D.; Challabathula, D.; Spittle, K.; Hall, R.; Gu, J.; Lyons, E.; et al. Single-molecule sequencing of the desiccation-tolerant grass Oropetium thomaeum. Nature 2015, 527, 508–511. [Google Scholar] [CrossRef] [Green Version]
- Zhu, Y.; Wang, B.; Phillips, J.; Zhang, Z.-N.; Du, H.; Xu, T.; Huang, L.-C.; Zhang, X.-F.; Xu, G.-H.; Li, W.-L.; et al. Global Transcriptome Analysis Reveals Acclimation-Primed Processes Involved in the Acquisition of Desiccation Tolerance in Boea hygrometrica. Plant Cell Physiol. 2015, 56, 1429–1441. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bentley, J.; Farrant, J. Field and acclimated metabolomes of a resurrection plant suggest strong environmental regulation in the extreme end of the species’ range. S. Afr. J. Bot. 2020, 135, 127–136. [Google Scholar] [CrossRef]
- Passon Née Gleichenhagen, M.; Weber, F.; Jung, N.; Bartels, D. Profiling of phenolic compounds in desiccation-tolerant and non-desiccation-tolerant Linderniaceae. Phytochem. Anal. PCA 2020, 32, 521–529. [Google Scholar] [CrossRef] [PubMed]
- Radermacher, A.L.; du Toit, S.F.; Farrant, J.M. Desiccation-Driven Senescence in the Resurrection Plant Xerophyta schlechteri (Baker) N.L. Menezes: Comparison of Anatomical, Ultrastructural, and Metabolic Responses Between Senescent and Non-Senescent Tissues. Front. Plant Sci. 2019, 10, 1396. [Google Scholar] [CrossRef]
- Abdelaal, K.; AlKahtani, M.; Attia, K.; Hafez, Y.; Király, L.; Künstler, A. The Role of Plant Growth-Promoting Bacteria in Alleviating the Adverse Effects of Drought on Plants. Biology 2021, 10, 520. [Google Scholar] [CrossRef]
- Khan, N.; Bano, A.; Rahman, M.A.; Guo, J.; Kang, Z.; Babar, M.A. Comparative Physiological and Metabolic Analysis Reveals a Complex Mechanism Involved in Drought Tolerance in Chickpea (Cicer arietinum L.) Induced by PGPR and PGRs. Sci. Rep. 2019, 9, 2097. [Google Scholar] [CrossRef]
- Moher, D.; Liberati, A.; Tetzlaff, J.; Altman, D.G.; Group, P. Preferred reporting items for systematic reviews and meta-analyses: The PRISMA statement. PLoS Med. 2009, 6, e1000097. [Google Scholar] [CrossRef] [Green Version]
- Page, M.J.; McKenzie, J.E.; Bossuyt, P.M.; Boutron, I.; Hoffmann, T.C.; Mulrow, C.D.; Shamseer, L.; Tetzlaff, J.M.; Akl, E.A.; Brennan, S.E.; et al. The PRISMA 2020 statement: An updated guideline for reporting systematic reviews. BMJ 2020, 372, 71. [Google Scholar]
- Moore, J.; Nguema-Ona, E.; Vicré-Gibouin, M.; Sørensen, I.; Willats, W.; Driouich, A.; Farrant, J. Arabinose-rich polymers as an evolutionary strategy to plasticize resurrection plant cell walls against desiccation. Planta 2013, 237, 739–754. [Google Scholar] [CrossRef]
- Willigen, C.V.; Farrant, J.M.; Pammenter, N.W. Anomalous Pressure Volume Curves of Resurrection Plants Do Not Suggest Negative Turgor. Ann. Bot. 2001, 88, 537–543. [Google Scholar] [CrossRef] [Green Version]
- Siddabathula, N.; Arigela, R.; Singh, R.; Prasanna, P.V. Taxonomy of endemic dwarf grass Oropetium villosulum Stapf ex Bor, an overlooked cohabitant of Oropetium thomaeum (L.f.) Trin. NeBIO 2019, 10, 168–174. [Google Scholar]
- Le, T.; Blomstedt, C.; Kuang, J.; Tenlen, J.; Gaff, D.; Hamill, J.; Neale, A. Desiccation-tolerance specific gene expression in leaf tissue of the resurrection plant Sporobolus stapfianus. Funct. Plant Biol. 2007, 34, 589–600. [Google Scholar] [CrossRef] [PubMed]
- Oliver, M.J.; Guo, L.; Alexander, D.C.; Ryals, J.A.; Wone, B.W.M.; Cushman, J.C. A Sister Group Contrast Using Untargeted Global Metabolomic Analysis Delineates the Biochemical Regulation Underlying Desiccation Tolerance in Sporobolus stapfianus. Plant Cell 2011, 23, 1231–1248. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yobi, A.; Schlauch, K.A.; Tillett, R.L.; Yim, W.C.; Espinoza, C.; Wone, B.W.M.; Cushman, J.C.; Oliver, M.J. Sporobolus stapfianus: Insights into desiccation tolerance in the resurrection grasses from linking transcriptomics to metabolomics. BMC Plant Biol. 2017, 17, 67. [Google Scholar] [CrossRef] [Green Version]
- Myers, M.Y.; Farrant, J.M.; Roden, L.C. Preliminary characterization of floral response of Xerophyta humilis to desiccation, vernalisation, photoperiod and light intensity. Plant Growth Regul. 2010, 62, 213–216. [Google Scholar] [CrossRef]
- Frank, W.; Munnik, T.; Kerkmann, K.; Salamini, F.; Bartels, D. A role for phospholipase D during early events of dehydration: Water-deficit triggers phospholipase D activity in the resurrection plant Craterostigma plantagineum. Eur. J. Endocrinol. 2000, 12, 111–124. [Google Scholar]
- Charuvi, D.; Nevo, R.; Shimoni, E.; Naveh, L.; Zia, A.; Adam, Z.; Farrant, J.M.; Kirchhoff, H.; Reich, Z. Photoprotection conferred by changes in photosynthetic protein levels and organization during dehydration of a homoiochlorophyllous resurrection plant. Plant Physiol. 2015, 167, 1554–1565. [Google Scholar] [CrossRef] [Green Version]
- Cooper, K.; Farrant, J.M. Recovery of the resurrection plant Craterostigma wilmsii from desiccation: Protection versus repair. J. Exp. Bot. 2002, 53, 1805–1813. [Google Scholar] [CrossRef] [Green Version]
- Phillips, J.; Fischer, E.; Baron, M.; Dries, N.; Facchinelli, F.; Kutzer, M.; Rahmanzadeh, R.; Remus, D.; Bartels, D. Lindernia brevidens: A novel desiccation-tolerant vascular plant, endemic to ancient tropical rainforests. Plant J. Cell Mol. Biol. 2008, 54, 938–948. [Google Scholar] [CrossRef]
- Moore, J.P.; Westall, K.L.; Ravenscroft, N.; Farrant, J.M.; Lindsey, G.G.; Brandt, W.F. The predominant polyphenol in the leaves of the resurrection plant Myrothamnus flabellifolius, 3, 4, 5 tri-O-galloylquinic acid, protects membranes against desiccation and free radical-induced oxidation. Biochem. J. 2005, 385, 301–308. [Google Scholar] [CrossRef] [Green Version]
- Gashi, B.; Abdullai, K.; Sota, V.; Kongjika, E. Micropropagation and in vitro conservation of the rare and threatened plants Ramonda serbica and Ramonda nathaliae. Physiol. Mol. Biol. Plants 2015, 21, 123–136. [Google Scholar] [CrossRef] [Green Version]
- Appleby, N.; Edwards, D.; Batley, J. New technologies for ultra-high throughput genotyping in plants. In Plant Genomics; Methods in Molecular Biology; Somers, D.J., Ed.; Humana Press: New York, NY, USA, 2009; pp. 19–39. [Google Scholar]
- Liu, X.; Giarola, V.; Quan, W.; Song, X.; Bartels, D. Identification and characterization of CTP:phosphocholine cytidylyltransferase CpCCT1 in the resurrection plant Craterostigma plantagineum. Plant Sci. 2020, 302, 110698. [Google Scholar] [CrossRef]
- Qiu, J.-R.; Xiang, X.-Y.; Wang, J.-T.; Xu, W.-X.; Chen, J.; Xiao, Y.; Jiang, C.-Z.; Huang, Z. MfPIF1 of Resurrection Plant Myrothamnus flabellifolia Plays a Positive Regulatory Role in Responding to Drought and Salinity Stresses in Arabidopsis. Int. J. Mol. Sci. 2020, 21, 3011. [Google Scholar] [CrossRef] [PubMed]
- Raza, A.; Razzaq, A.; Mehmood, S.; Hussain, M.; Wei, S.; He, H.; Zaman, Q.; Xuekun, Z.; Hasanuzzaman, M. Omics: The way forward to enhance abiotic stress tolerance in Brassica napus L. GM Crop. Food 2021, 12, 251–281. [Google Scholar] [CrossRef]
- Guo, J.; Huang, Z.; Sun, J.; Cui, X.; Liu, Y. Research Progress and Future Development Trends in Medicinal Plant Transcriptomics. Front. Plant Sci. 2021, 12, 691838. [Google Scholar] [CrossRef]
- Lowe, R.; Shirley, N.; Bleackley, M.; Dolan, S.; Shafee, T. Transcriptomics technologies. PLoS Comput. Biol. 2017, 13, e1005457. [Google Scholar] [CrossRef] [Green Version]
- Ma, C.; Wang, H.; Macnish, A.J.; Estrada-Melo, A.C.; Lin, J.; Chang, Y.; Reid, M.S.; Jiang, C.Z. Transcriptomic analysis reveals numerous diverse protein kinases and transcription factors involved in desiccation tolerance in the resurrection plant Myrothamnus flabellifolia. Hortic. Res. 2015, 2, 15034. [Google Scholar] [CrossRef] [Green Version]
- Vidovic, M.; Cukovic, K. Isolation of high-quality RNA from recalcitrant leaves of variegated and resurrection plants. 3Biotech 2020, 10, 3. [Google Scholar] [CrossRef]
- Wang, L.; Shang, H.; Liu, Y.; Zheng, M.; Wu, R.; Phillips, J.; Bartels, D.; Deng, X. A role for a cell wall localized glycine-rich protein in dehydration and rehydration of the resurrection plant Boea hygrometrica. Plant Biol. 2009, 11, 837–848. [Google Scholar] [CrossRef]
- Abdalla, K.; Rafudeen, M. Analysis of the nuclear proteome of the resurrection plant Xerophyta viscosa in response to dehydration stress using iTRAQ with 2DLC and tandem mass spectrometry. J. Proteom. 2012, 75, 2361–2374. [Google Scholar] [CrossRef]
- Aslam, B.; Basit, M.; Nisar, M.A.; Khurshid, M.; Rasool, M.H. Proteomics: Technologies and Their Applications. J. Chromatogr. Sci. 2017, 55, 182–196. [Google Scholar] [CrossRef] [Green Version]
- Röhrig, H.; Colby, T.; Schmidt, J.; Harzen, A.; Facchinelli, F.; Bartels, D. Analysis of desiccation-induced candidate phosphoproteins from Craterostigma plantagineum isolated with a modified metal oxide affinity chromatography procedure. Proteomics 2008, 8, 3548–3560. [Google Scholar] [CrossRef] [PubMed]
- Piasecka, A.; Kachlicki, P.; Stobiecki, M. Analytical methods for detection of plant metabolomes changes in response to biotic and abiotic stresses. Int. J. Mol. Sci. 2019, 20, 379. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Plancot, B.; Vanier, G.; Maire, F.; Bardor, M.; Lerouge, P.; Farrant, J.; Moore, J.; Driouich, A.; Vicré-Gibouin, M.; Afonso, C.; et al. Structural characterization of arabinoxylans from two African plant species Eragrostis nindensis and Eragrostis tef using various mass spectrometric methods. Rapid Commun. Mass Spectrom. RCM 2014, 28, 908–916. [Google Scholar] [CrossRef] [PubMed]
- Zhao, C.; Zhang, Y.; Du, J.; Guo, X.; Wen, W.; Gu, S.; Wang, J.; Fan, J. Crop Phenomics: Current Status and Perspectives. Front. Plant Sci. 2019, 10, 714. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wasaya, A.; Zhang, X.; Fang, Q.; Yan, Z. Root phenotyping for drought tolerance: A review. Agronomy 2018, 8, 241. [Google Scholar] [CrossRef] [Green Version]
- Anand, G.; Goel, V.; Dubey, S.; Sharma, S. Tailoring the rhizospheric microbiome of Vigna radiata by adaptation to salt stress. Plant Growth Regul. 2021, 93, 79–88. [Google Scholar] [CrossRef]
- Kaushal, M.; Mahuku, G.; Swennen, R. Metagenomic Insights of the Root Colonizing Microbiome Associated with Symptomatic and Non-Symptomatic Bananas in Fusarium Wilt Infected Fields. Plants 2020, 9, 263. [Google Scholar] [CrossRef] [Green Version]
- Parween, T.; Bhandari, P.; Jan, S.; Raza, S. Interaction between pesticide and soil microorganisms and their degradation: A molecular approach. In Plant, Soil and Microbes; Hakeem, K.A.M., Ed.; Springer: Berlin/Heidelberg, Germany, 2016; pp. 23–43. [Google Scholar]
- Piombo, E.; Abdelfattah, A.; Droby, S.; Wisniewski, M.; Spadaro, D.; Schena, L. Metagenomics approaches for the detection and surveillance of emerging and recurrent plant pathogens. Microorganisms 2021, 9, 188. [Google Scholar] [CrossRef]
- Vassileva, V.; Moyankova, D.; Dimitrova, A.; Mladenov, P.; Djilianov, D. Assessment of leaf micromorphology after full desiccation of resurrection plants. Plant Biosyst. Int. J. Deal. All Asp. Plant Biol. 2018, 153, 108–117. [Google Scholar] [CrossRef]
- Patra, J.K.; Mishra, R.R.; Rout, S.D.; Thatoi, H.N. An assessment of Nutrient Content of Different Grass Species of Similipal Tiger Reserve, Orissa. World J. Agric. Sci. 2011, 7, 34–41. [Google Scholar]
- Gaff, D.F.; Oliver, M. The evolution of desiccation tolerance in angiosperm plants: A rare yet common phenomenon. Funct. Plant Biol. 2013, 40, 315–328. [Google Scholar] [CrossRef] [PubMed]
- Farrant, J.M. A comparison of mechanisms of desiccation tolerance among three angiosperm resurrection plant species. Plant Ecol. 2000, 151, 29–39. [Google Scholar] [CrossRef]
- Jovanovic, Z.; Rakic, T.; Stevanovic, B.; Radovic, S. Characterization of oxidative and antioxidative events during dehydration and rehydration of resurrection plant Ramonda nathaliae. Plant Growth Regul. 2011, 64, 231–240. [Google Scholar] [CrossRef]
- Moore, J.P.; Farrant, J.M.; Lindsey, G.G.; Brandt, W.F. The South African and Namibian populations of the resurrection plant Myrothamnus flabellifolius are genetically distinct and display variation in their galloylquinic acid composition. J. Chem. Ecol. 2005, 31, 2823–2834. [Google Scholar] [CrossRef] [PubMed]
- Willigen, C.; Pammenter, N.; Mundree, S.; Farrant, J. Some physiological comparisons between the resurrection grass, Eragrostis nindensis, and the related desiccation-sensitive species, E. curvula. Plant Growth Regul. 2001, 35, 121–129. [Google Scholar] [CrossRef]
- Mihailova, G.; Solti, Á.; Sarvari, E.; Keresztes, Á.; Rapparini, F.; Velitchkova, M.; Simova, L.; Aleksandrov, V.; Georgieva, K. Freezing tolerance of photosynthetic apparatus in the homoiochlorophyllous resurrection plant Haberlea rhodopensis. Environ. Exp. Bot. 2020, 178, 104157. [Google Scholar] [CrossRef]
- Hamdi, Y.; Zass, L.; Othman, H.; Radouani, F.; Allali, I.; Hanachi, M.; Okeke, C.J.; Chaouch, M.; Tendwa, M.B.; Samtal, C. Human OMICs and Computational Biology Research in Africa: Current Challenges and Prospects. OMICS J. Integr. Biol. 2021, 25, 213–233. [Google Scholar] [CrossRef] [PubMed]
- Rapparini, F.; Neri, L.; Mihailova, G.; Petkova, S.; Georgieva, K. Growth irradiance affects the photoprotective mechanisms of the resurrection angiosperm Haberlea rhodopensis Friv. in response to desiccation and rehydration at morphological, physiological and biochemical levels. Environ. Exp. Bot. 2015, 113, 67–79. [Google Scholar] [CrossRef]
- Karbaschi, P.; Williams, B.; Taji, A.; Mundree, S. Tripogon loliiformis elicits a rapid physiological and structural response to dehydration for desiccation tolerance. Funct. Plant Biol. 2016, 43, 1–13. [Google Scholar] [CrossRef]
- Lyall, R.; Gechev, T. Multi-Omics Insights into the Evolution of Angiosperm Resurrection Plants. Annu. Plant Rev. Online 2020, 3, 77–110. [Google Scholar]
- Farrant, J.M.; Moore, J.P.; Hilhorst, H.W.M. Editorial: Unifying Insights into the Desiccation Tolerance Mechanisms of Resurrection Plants and Seeds. Front. Plant Sci. 2020, 11, 1089. [Google Scholar] [CrossRef] [PubMed]
- Siljak-Yakovlev, S.; Stevanovic, V.; Tomasevic, M.; Brown, S.C.; Stevanovic, B. Genome size variation and polyploidy in the resurrection plant genus Ramonda: Cytogeography of living fossils. Environ. Exp. Bot. 2008, 62, 101–112. [Google Scholar] [CrossRef]
- Costa, M.-C.D.; Cooper, K.; Hilhorst, H.W.M.; Farrant, J.M. Orthodox Seeds and Resurrection Plants: Two of a Kind? Plant Physiol. 2017, 175, 589–599. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, X.; Legay, S.; Sergeant, K.; Zorzan, S.; Leclercq, C.C.; Charton, S.; Giarola, V.; Liu, X.; Challabathula, D.; Renaut, J.; et al. Molecular insights into plant desiccation tolerance: Transcriptomics, proteomics and targeted metabolite profiling in Craterostigma plantagineum. Plant J. 2021, 107, 377–398. [Google Scholar] [CrossRef] [PubMed]
- Jiang, G.; Wang, Z.; Shang, H.; Yang, W.; Hu, Z.; Phillips, J.; Deng, X. Proteome analysis of leaves from the resurrection plant Boea hygrometrica in response to dehydration and rehydration. Planta 2007, 225, 1405–1420. [Google Scholar] [CrossRef]
- Ingle, R.A.; Schmidt, U.G.; Farrant, J.M.; Thomson, J.A.; Mundree, S.G. Proteomic analysis of leaf proteins during dehydration of the resurrection plant Xerophyta viscosa. Plant Cell Environ. 2007, 30, 435–446. [Google Scholar] [CrossRef]
- Moyankova, D.; Mladenov, P.; Berkov, S.; Peshev, D.; Georgieva, D.; Djilianov, D. Metabolic profiling of the resurrection plant Haberlea rhodopensis during desiccation and recovery. Physiol. Plant. 2014, 152, 675–687. [Google Scholar] [CrossRef]
- Sun, R.-Z.; Lin, C.-T.; Zhang, X.-F.; Duan, L.-X.; Qi, X.-Q.; Gong, Y.-H.; Deng, X. Acclimation-induced metabolic reprogramming contributes to rapid desiccation tolerance acquisition in Boea hygrometrica. Environ. Exp. Bot. 2018, 148, 70–84. [Google Scholar] [CrossRef]
- Bentley, J.; Moore, J.; Farrant, J. Metabolomic Profiling of the Desiccation-Tolerant Medicinal Shrub Myrothamnus flabellifolia Indicates Phenolic Variability Across Its Natural Habitat: Implications for Tea and Cosmetics Production. Molecules 2019, 24, 1240. [Google Scholar] [CrossRef] [Green Version]
- Vidovic, M.; Franchin, C.; Morina, F.; Veljovic-Jovanovic, S.; Masi, A.; Arrigoni, G. Efficient protein extraction for shotgun proteomics from hydrated and desiccated leaves of resurrection Ramonda serbica plants. Anal. Bioanal. Chem. 2020, 412, 8299–8312. [Google Scholar] [CrossRef]
- Dai, A. Increasing drought under global warming in observations and models. Nat. Clim. Chang. 2013, 3, 52–58. [Google Scholar] [CrossRef]
- Thornton, P.; Jones, P.; Alagarswamy, G.; Andresen, J.; Herrero, M. Adapting to climate change: Agricultural system and household impacts in East Africa. Agric. Syst. 2010, 103, 73–82. [Google Scholar] [CrossRef]
- Farrant, J. Plants that Rise from the Dead Breathe Life into Research. Available online: https://lagenceodyc.com/wp-content/uploads/2019/02/Brand_content_Nature_Armani_20190207-compressed.pdf (accessed on 5 December 2021).
- Kamies, R.; Rafudeen, M.; Farrant, J. The use of aeroponics to investigate antioxidant activity in the roots of Xerophyta viscosa. Plant Growth Regul. 2010, 62, 203–211. [Google Scholar] [CrossRef]
- Péli, E.R.; Mihailova, G.; Petkova, S.; Tuba, Z.; Georgieva, K. Differences in physiological adaptation of Haberlea rhodopensis Friv. leaves and roots during dehydration–rehydration cycle. Acta Physiol. Plant. 2012, 34, 947–955. [Google Scholar] [CrossRef]
- Norwood, M.; Toldi, O.; Richter, A.; Scott, P. Investigation into the ability of roots of the poikilohydric plant Craterostigma plantagineum to survive dehydration stress. J. Exp. Bot. 2003, 54, 2313–2321. [Google Scholar] [CrossRef]
- Djilianov, D.L.; Dobrev, P.I.; Moyankova, D.P.; Vankova, R.; Georgieva, D.T.; Gajdošová, S.; Motyka, V. Dynamics of Endogenous Phytohormones during Desiccation and Recovery of the Resurrection Plant Species Haberlea rhodopensis. J. Plant Growth Regul. 2013, 32, 564–574. [Google Scholar] [CrossRef]
- Đokić, L.; Savić, M.; Narančić, T.; Vasiljević, B. Metagenomic analysis of soil microbial communities. Arch. Biol. Sci. 2010, 62, 559–564. [Google Scholar] [CrossRef]
- Farrant, J.M.; Hilhorst, H.W.M. What is dry? Exploring metabolism and molecular mobility at extremely low water contents. J. Exp. Bot. 2021, 72, 1507–1510. [Google Scholar] [CrossRef]
Figure 1.
Systematic reviews and Meta–Analyses (PRISMA) flow chart outlining the exclusion and selection procedure used in the current meta-analysis with corresponding records from the database of angiosperm resurrection plants.
Figure 1.
Systematic reviews and Meta–Analyses (PRISMA) flow chart outlining the exclusion and selection procedure used in the current meta-analysis with corresponding records from the database of angiosperm resurrection plants.

Figure 2.
The number of studies published on each resurrection plant species. Species are ordered phylogenetically. Studies are categorized as either native (conducted by researchers working in the native range of the species), collaborative (involving researchers from both within and beyond the native range of the species) or international (studies conducted by a team working outside of the native range of the species).
Figure 2.
The number of studies published on each resurrection plant species. Species are ordered phylogenetically. Studies are categorized as either native (conducted by researchers working in the native range of the species), collaborative (involving researchers from both within and beyond the native range of the species) or international (studies conducted by a team working outside of the native range of the species).

Figure 3.
Map showing where the model resurrection plants are native to relative to where they have been studied. Circles indicate the native area of the species and are scaled by the number of studies conducted in their native range. Arrows point to the location where international and collaborative studies have been conducted and are scaled by the number of studies. Dashed arrows are for collaborative studies and solid arrows are for international studies. Plants in the same genus (or family for grasses) are consolidated for simplicity and studies are grouped by continent.
Figure 3.
Map showing where the model resurrection plants are native to relative to where they have been studied. Circles indicate the native area of the species and are scaled by the number of studies conducted in their native range. Arrows point to the location where international and collaborative studies have been conducted and are scaled by the number of studies. Dashed arrows are for collaborative studies and solid arrows are for international studies. Plants in the same genus (or family for grasses) are consolidated for simplicity and studies are grouped by continent.

Figure 4.
The number of studies exploring desiccation tolerance mechanisms of resurrection plants in the past two decades using various techniques. (A) Research articles published per year from 2000–2020 focused on Genomic (genetics), Transcriptomic (RNA), Proteomic (proteins), Metabolomic (metabolites), Metagenomic (microbes), and Physiological (biochemical) techniques to understand the mechanisms of desiccation tolerance in resurrection plants. (B) Analysis of publication trends of five most widely analyzed angiosperm resurrection plants, namely B. hygrometrica, C. plantagineum, H. rhodopensis, M. flabellifolia and X. schlechteri studies per year.
Figure 4.
The number of studies exploring desiccation tolerance mechanisms of resurrection plants in the past two decades using various techniques. (A) Research articles published per year from 2000–2020 focused on Genomic (genetics), Transcriptomic (RNA), Proteomic (proteins), Metabolomic (metabolites), Metagenomic (microbes), and Physiological (biochemical) techniques to understand the mechanisms of desiccation tolerance in resurrection plants. (B) Analysis of publication trends of five most widely analyzed angiosperm resurrection plants, namely B. hygrometrica, C. plantagineum, H. rhodopensis, M. flabellifolia and X. schlechteri studies per year.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
The best studied angiosperm resurrection plants in the past two decades and their native range.
Table 1.
The best studied angiosperm resurrection plants in the past two decades and their native range.
| Resurrection Plant | Continent | Region | Reference |
|---|---|---|---|
| Monocot | |||
| E. nindensis | Africa | Southern Africa | [29,30] |
| O. thomaeum | Asia and East Africa | India, Northeast tropical and East tropical Africa | [31] |
| S. stapfianus | Africa | South Africa | [32,33,34] |
| T. loliioformis | Australia | Australia | [15] |
| X. humilis | Africa | Southern Africa | [29,35] |
| X. schlechteri | Africa | Lesotho, South Africa, Swaziland | [10,24,29] |
| Dicot | |||
| B. hygrometrica | Asia | China | [7] |
| C. plantagineum | Africa | Kenya, South Africa | [36] |
| C. pumilum | Africa | East Africa | [37] |
| C. wilmsii | Africa | South Africa | [38] |
| H. rhodopensis | Europe | Bulgaria | [6] |
| L. brevidens | Africa | Kenya | [39] |
| M. flabellifolia | Africa | Namibia, South Africa, Zimbabwe | [40] |
| R. nathaliae | Europe | Serbia, Bulgaria | [41] |
| R. serbica | Europe | Serbia, Bulgaria | [8] |
Table 2.
Tools and technologies used for investigation of desiccation tolerance mechanisms in resurrection plants.
Table 2.
Tools and technologies used for investigation of desiccation tolerance mechanisms in resurrection plants.
| Technology–Based Approach | Genomics | Transcriptomics | Proteomics | Metabolomics | Physiology | Metagenomics |
|---|---|---|---|---|---|---|
| DNA sequencing, genetic profile, genetic mapping, structural & functional genomics | RNA sequencing, expression profiling, transcriptional regulation | Protein identification, quantification, Translation modification | Metabolites profiling, identification & quantification | Morphological, biochemical & phenotypical characterization | Bacterial and fungal and viral nucleic acids analysis | |
| Methodology and Quantitative techniques |
|
|
|
|
|
|
| High–throughput techniques |
|
|
|
|
|
|
| References | [42,43,44,45] | [15,46,47,48,49,50] | [51,52,53] | [23,54,55] | [24,39,56,57] | [58,59,60,61] |
Key: cDNA-AFLP (cDNA-amplified fragment length polymorphism); qRT-PCR (real-time quantitative PCR); RNA-PET-seq (paired end sequencing); sRNA-seq (small RNA sequencing); SDS-PAGE (sodium dodecyl sulphate–polyacrylamide gel electrophoresis); iTRAQ (isobaric tag for relative and absolute quantitation); LC-MS (liquid chromatography-mass spectrometry); NMR (nuclear magnetic resonance).
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Tebele, S.M.; Marks, R.A.; Farrant, J.M. Two Decades of Desiccation Biology: A Systematic Review of the Best Studied Angiosperm Resurrection Plants. Plants 2021, 10, 2784. https://doi.org/10.3390/plants10122784
AMA Style
Tebele SM, Marks RA, Farrant JM. Two Decades of Desiccation Biology: A Systematic Review of the Best Studied Angiosperm Resurrection Plants. Plants. 2021; 10(12):2784. https://doi.org/10.3390/plants10122784
Chicago/Turabian StyleTebele, Shandry M., Rose A. Marks, and Jill M. Farrant. 2021. "Two Decades of Desiccation Biology: A Systematic Review of the Best Studied Angiosperm Resurrection Plants" Plants 10, no. 12: 2784. https://doi.org/10.3390/plants10122784
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.