

Metabolite Profile of Athrixia phylicoides DC. (Bush Tea) and Determination of Inhibitory Mechanism against Tyrosinase Enzyme from Mushroom
 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Preparation
2.2. Inhibition of Mushroom Tyrosinase Activity Assay
2.2.1. Thin-Layer Chromatography (TLC) Autography Assay
2.2.2. Tyrosinase-Based TLC Autography
2.2.3. Determination of Tyrosinase Inhibition Using Lineweaver–Burk Plots
2.3. Statistical Analysis
3. Results and Discussion
3.1. Tyrosinase-Based TLC Autography
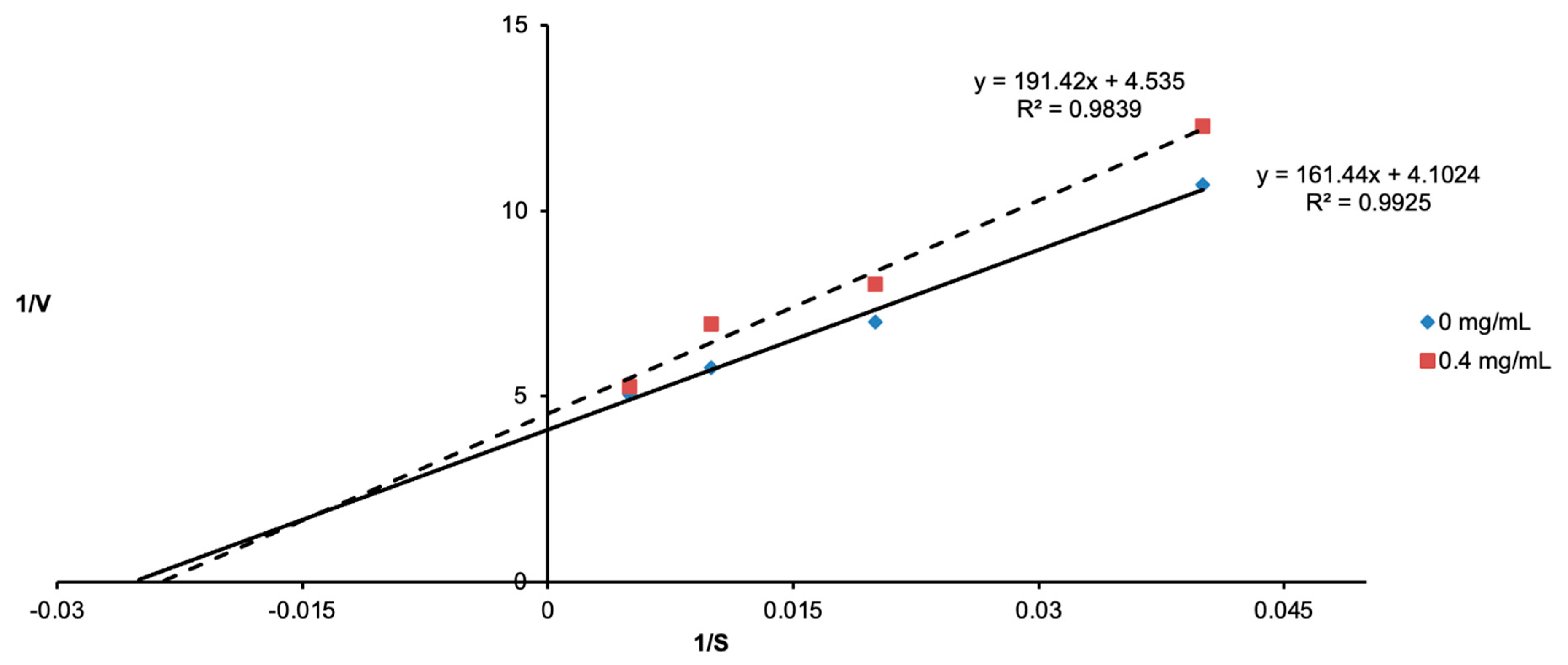
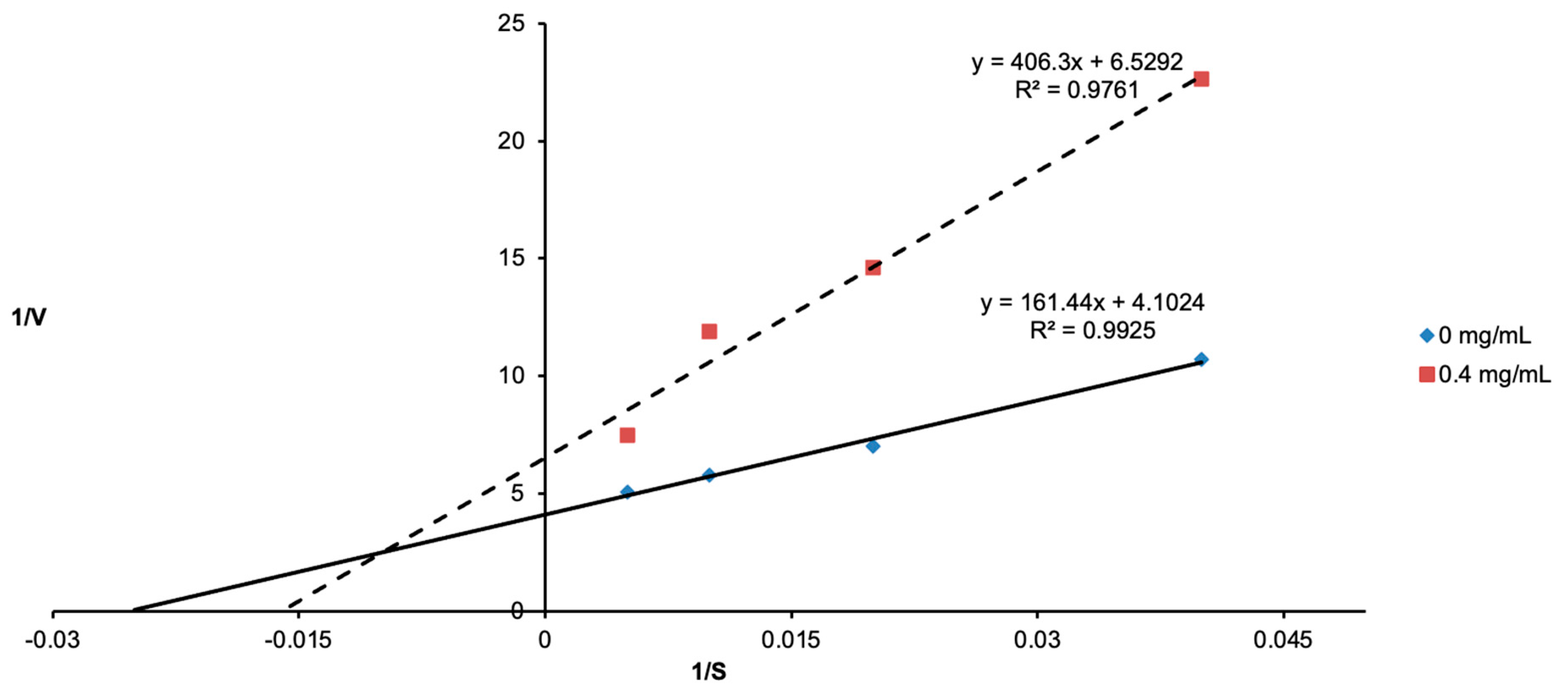
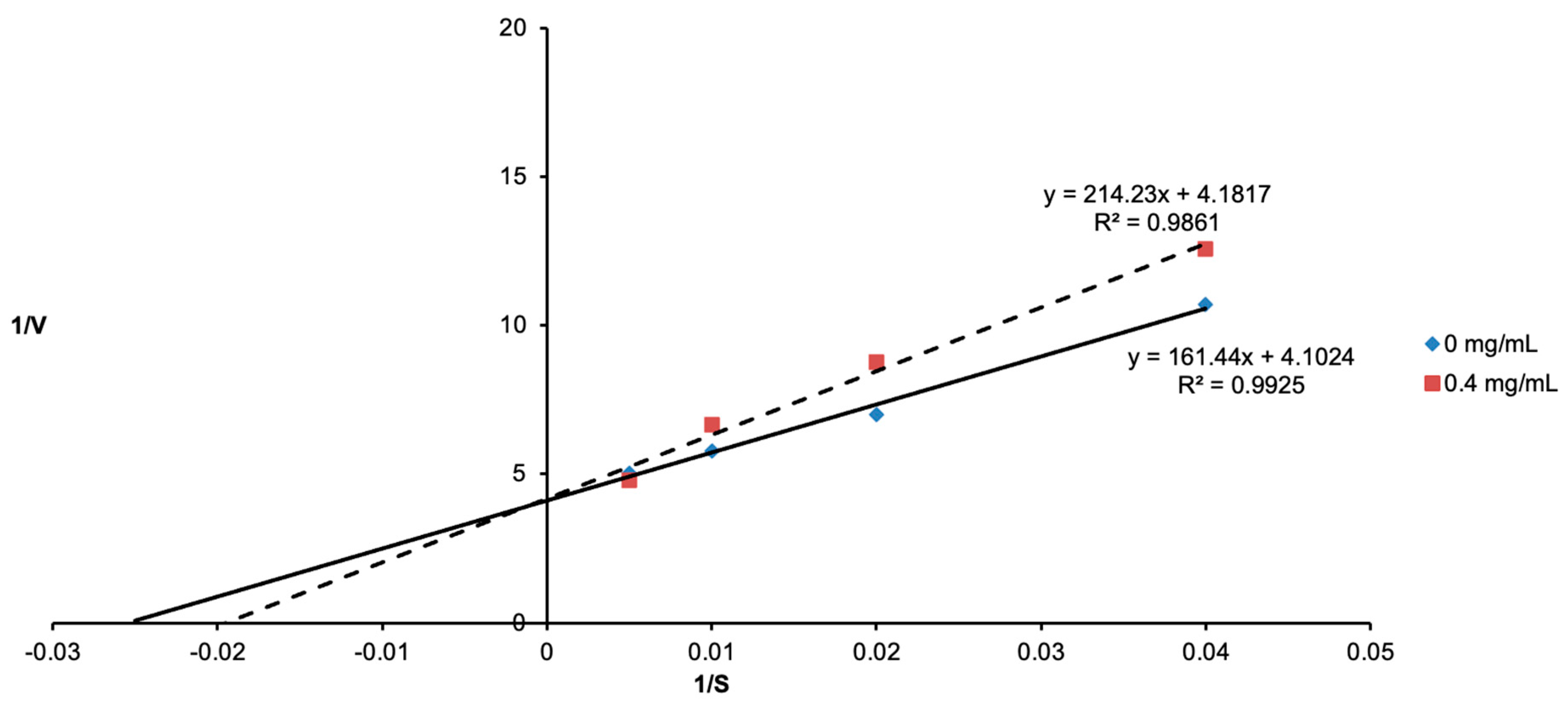
3.2. Determination of Tyrosinase Inhibition Using Lineweaver–Burk Plots
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Roberts, M.M.J. Indigenous Healing Plants, 1st ed.; Southern Book Publishers: Halfway House, South Africa, 1990. [Google Scholar]
- Hutchings, A.; Scott, A.H.; Lewis, G.; Cunningham, A. Zulu Medicinal Plants: An Inventory; University of Natal press: Pietermaritzburg, South Africa, 1996. [Google Scholar]
- Rampedi, I.; Olivier, J. The use and potential commercial development of Athrixia phylicoides. Acta Acad. 2005, 37, 165–183. [Google Scholar]
- Mudau, F.N.; Soundy, P.; du Toit, E.S.; Olivier, J. Variation in phenolic content of Athrixia phylicoides (L.) (bush tea) leaves with season and nitrogen application. S. Afr. J. Bot. 2006, 72, 398–402. [Google Scholar] [CrossRef] [Green Version]
- Olivier, J.; Symington, E.A.; Jonker, C.Z.; Rampedi, I.T.; van Eeden, T.S. Comparison of mineral composition of leaves and infusions of traditional and herbal teas. S. Afr. J. Sci. 2012, 108, 1–7. [Google Scholar] [CrossRef]
- Mohale, K.C.; Abrha, A.H.; Hlengilizwe, N.; Hintsa, A.T.; Umer, M.; Mudau, F.N.; Ali, S.W.; Afzal, M.I.; Martorell, M.; Salehi, B.; et al. Untargeted profiling of field cultivated bush tea (Athrixia phylicoides DC.) based on metabolite analysis. Cell. Mol. Biol. 2020, 66, 104–109. [Google Scholar]
- Thibane, V.S.; Ndhlala, A.R.; Finnie, J.F.; Van Staden, J. Cosmeceutical efficiency by some plant extracts used traditionally for skincare in inhibiting tyrosinase activity in a human epidermal melanocytes (HEM) cell line. S. Afr. J. Bot. 2019, 126, 256–260. [Google Scholar] [CrossRef]
- Kleynhans, R.; Singh, S.; Kishore, N.; Lall, N. Investigation towards propagation and cosmeceutical application of Athrixia phylicoides DC. S. Afr. J. Bot. 2017, 112, 319–321. [Google Scholar] [CrossRef]
- Joubert, E.; Gelderblom, W.C.A.; Louw, A.; de Beer, D. South African herbal teas: Aspalathus linearis, Cyclopia spp. and Athrixia phylicoides-A review. J. Ethnopharmacol. 2008, 119, 376–412. [Google Scholar] [CrossRef]
- Madzinga, M.; Kritzinger, Q.; Lall, N. Medicinal plants used in the treatment of superficial skin infections: From traditional medicine to herbal soap formulations. In Medicinal Plants for Holistic Health and Well-Being, 1st ed.; Lall, N., Ed.; Academic Press Publishers: Pretoria, South Africa, 2018; pp. 255–275. [Google Scholar]
- Maudu, M.E.; Mudau, F.N.; Mariga, I.K. Quality profiles of cultivated and wild bush tea (Athrixia phylicoides) harvested at various phenological stages. Int. J. Agric. Biol. 2012, 14, 144–148. [Google Scholar]
- Mohale, K.C.; Hintsa, A.T.; Emanuel, M.A.; Mudau, F.N. Metabolic profiling of cultivated bush tea (Athrixia phylicoides DC) in response to different pruning types. HortScience 2018, 53, 993–998. [Google Scholar] [CrossRef] [Green Version]
- Mudau, F.N.; Makunga, P.N. Effect of seasonal variation and growth conditions on carbohydrate partitioning in different organs and the quality of bush tea. HortScience 2018, 53, 999–1005. [Google Scholar]
- de Beer, D.; Joubert, E.; Malherbe, C.J.; Brand, D.J. Use of countercurrent chromatography during isolation of 6-hydroxyluteolin-7-O-β-glucoside, a major antioxidant of Athrixia phylicoides. J. Chromatogr. A 2011, 1218, 6179–6186. [Google Scholar] [CrossRef] [PubMed]
- Wagner, H.; Bladt, S. Plant Drug Analysis: A Thin Layer Chromatograph Atlas, 2nd ed.; Springer-Verlag Berlin Heidelberg: New York, NY, USA; Tokyo, Japan, 1996. [Google Scholar]
- Hsu, K.-D.; Chan, Y.-H.; Chen, H.-J.; Lin, S.-P.; Cheng, K.-C. Tyrosinase-based TLC autography for anti-melanogenic drug screening. Sci. Rep. 2018, 8, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mapunya, M.B.; Hussein, A.A.; Rodriguez, B.; Lall, N. Tyrosinase activity of Greyia flanaganii (Bolus) constituents. Phytomedicine 2011, 18, 1006–1012. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reichelt, K.V.; Hoffmann-Lucke, P.; Hartmann, B.; Weber, B.; Ley, J.P.; Krammer, G.E.; Swanepoel, K.M.; Engel, K.-H. Phytochemical characterization of South African bush tea (Athrixia Phylicoides DC.). S. Afr. J. Bot. 2012, 83, 1–8. [Google Scholar] [CrossRef]
- Lerotholi, L.J.; Chaudhary, S.K.; Chen, W.; Veale, C.G.L.; Combrinck, S.; Viljoen, A.M. Identification, isolation and determination of biomarkers for quality control of bush tea (Athrixia phylicoides). Planta Med. 2018, 84, 902–912. [Google Scholar]
- Fan, M.; Zhang, G.; Hu, X.; Xu, X.; Gong, D. Quercetin as a tyrosinase inhibitor: Inhibitory activity, conformational change and mechanism. Food Res. Int. 2017, 100, 226–233. [Google Scholar] [CrossRef]
- Tyteca, E.; Talebi, M.; Amos, R.; Park, S.H.; Taraji, M.; Wen, Y.; Szucs, R.; Pohl, C.A.; Dolan, J.W.; Haddad, P.R. Towards a chromatographic similarity index to establish localized quantitative structure-retention models for retention prediction: Use of retention factor ratio. J. Chromatogr. A 2017, 1486, 50–58. [Google Scholar] [CrossRef]
- Tshivhandekano, I.; Ngezimana, W.; Tshikalange, T.E.; Makunga, N.P.; Mudau, F.N. Nitrogen application influences quality, pharmacological activities and metabolite profile of Athrixia phylicoides DC. (Bush tea) cultivated under green house and field conditions. Acta Agric. Scand. B Soil Plant. Sci. 2018, 68, 388–400. [Google Scholar]
- Indrianingsih, A.W.; Tachibana, S.; Itoh, K. In vitro evaluation of antioxidant and α-glucosidase inhibitory assay of several tropical and subtropical plants. Procedia Environ. Sci. 2015, 28, 639–648. [Google Scholar] [CrossRef] [Green Version]
- Saghaie, L.; Pourfarzam, M.; Fassihi, A.; Sartippour, B. Synthesis and tyrosinase inhibitory properties of some novel derivatives of kojic acid. Res. Pharm. Sci. 2013, 8, 233–242. [Google Scholar]
- Cabanes, J.; Chazarra, S.; Garćia-Carmona, F. Kojic acid, a cosmetic skin whitening agent, is a slow binding inhibitor of catecholase activity of tyrosinase. J. Pharm. Pharmacol. 1994, 46, 982–985. [Google Scholar] [CrossRef] [PubMed]
- Chang, T.S. An updated review of tyrosinase inhibitors. Int. J. Mol. Sci. 2009, 10, 2440–2475. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Q.-X.; Kubo, I. Kinetics of mushroom tyrosinase inhibition in quercetin. J. Agric. Food Chem. 2002, 50, 4108–4112. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound | Retention Factor (Rf) |
|---|---|
| Quercetin a | 0.94 |
| Gallic acid a | 0.85 |
| Unknown 1 a | 0.73 |
| Unknown 2 a | 0.44 |
| Chlorogenic acid a | 0.19 |
| Kojic acid b | 0.19 |
| Compound | IC50 (µg/mL) | Inhibitory Strength (IC50I/IC50KA) |
|---|---|---|
| A. phylicoidesa | 20.65 ± 0.14 | 3.96 |
| Quercetin a | 51.07 ± 2.43 | 9.78 |
| Chlorogenic acid a | 15.25 ± 1.18 | 2.92 |
| Kojic acid b | 5.22 ± 1.44 | NA |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Thibane, V.S.; Ramphinwa, M.L.; Mchau, G.R.A.; Mudau, F.N. Metabolite Profile of Athrixia phylicoides DC. (Bush Tea) and Determination of Inhibitory Mechanism against Tyrosinase Enzyme from Mushroom. Cosmetics 2022, 9, 133. https://doi.org/10.3390/cosmetics9060133
Thibane VS, Ramphinwa ML, Mchau GRA, Mudau FN. Metabolite Profile of Athrixia phylicoides DC. (Bush Tea) and Determination of Inhibitory Mechanism against Tyrosinase Enzyme from Mushroom. Cosmetics. 2022; 9(6):133. https://doi.org/10.3390/cosmetics9060133
Chicago/Turabian StyleThibane, Vuyisile Samuel, Maanea Lonia Ramphinwa, Godwin Richard Ainamensa Mchau, and Fhatuwani Nixwell Mudau. 2022. "Metabolite Profile of Athrixia phylicoides DC. (Bush Tea) and Determination of Inhibitory Mechanism against Tyrosinase Enzyme from Mushroom" Cosmetics 9, no. 6: 133. https://doi.org/10.3390/cosmetics9060133