
Agro-Morphological Variability of Wild Vigna Species Collected in Senegal
 , , ,
, , , 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material
2.2. Site of Study and Meteorological Conditions
2.3. Experimental Design and Sowing Process
2.4. Data Collection and Analysis
3. Results
3.1. Qualitative Traits of the Wild Vigna Species
3.1.1. Morphological Variability of the Whole Set of Vigna Accessions
3.1.2. Multiple Correspondence Analysis
3.1.3. Relationship between the Wild Vigna Accessions
3.2. Diversity Description Based on Quantitative Traits
3.2.1. Descriptive Statistics Analysis
3.2.2. Analysis of Variance
3.2.3. Principal Component Analysis
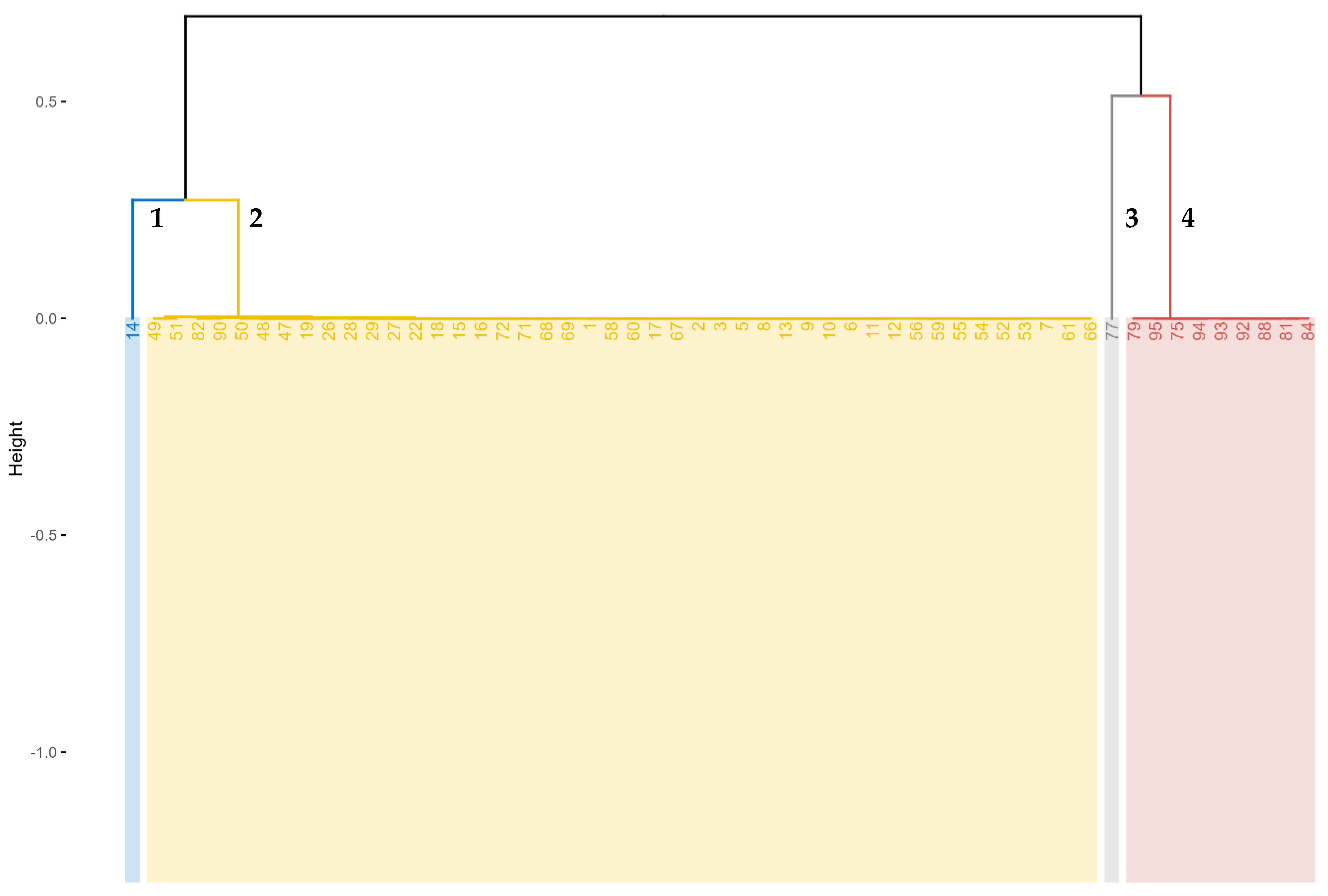
3.2.4. Cluster Analysis Showing the Grouping of the Accessions Based on the Quantitative Traits
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Oikeh, S.O.; Niang, A.; Abaidoo, R.; Houngnandan, P.; Futakuchi, K.; Koné, B.; Touré, A. Enhancing Rice Productivity and Soil Nitrogen Using Dual-Purpose Cowpea-NERICA® Rice Sequence in Degraded Savanna. J. Life Sci. 2012, 6, 1237–1250. [Google Scholar]
- Lonardi, S.; Muñoz-Amatriaín, M.; Liang, Q.; Shu, S.; Wanamaker, S.I.; Lo, S.; Tanskanen, J.; Schulman, A.H.; Zhu, T.; Luo, M.-C.; et al. The genome of cowpea (Vigna unguiculata [L.] Walp). Plant J. 2019, 98, 767–782. [Google Scholar] [CrossRef] [PubMed]
- Xiong, H.; Shi, A.; Mou, B.; Qin, J.; Motes, D.; Lu, W.; Ma, J.; Weng, Y.; Yang, W.; Wu, D. Genetic diversity and population structure of cowpea (Vigna unguiculata L. Walp). PLoS ONE 2016, 11, e0160941. [Google Scholar] [CrossRef] [PubMed]
- ANSD. Bulletin Mensuel des Statistiques Économiques d’Octobre 2020. Available online: https://www.ansd.sn (accessed on 18 October 2023).
- Ehlers, J.D.; Hall, A.E. Genotypic Classification of Cowpea Based on Responses to Heat and Photoperiod. Crop. Sci. 1996, 36, 673–679. [Google Scholar] [CrossRef]
- Elowad, H.O.; Hall, A.E. Influences of early and late nitrogen fertilization on yield and nitrogen fixation of cowpea under well-watered and dry field conditions. Field Crop. Res. 1987, 15, 229–244. [Google Scholar] [CrossRef]
- Hall, A.E. Breeding for adaptation to drought and heat in cowpea. Eur. J. Agron. 2004, 21, 447–454. [Google Scholar] [CrossRef]
- Ba, F.S.; Pasquet, R.S.; Gepts, P. Genetic diversity in cowpea [Vigna unguiculata (L.) Walp] as revealed by RAPD markers. Genet. Resour. Crop. E 2004, 51, 539–550. [Google Scholar] [CrossRef]
- Difo, V.H.; Onyike, E.; Ameh, D.A.; Njoku, G.C.; Ndidi, U.S. Changes in nutrient and antinutrient composition of Vigna racemosa flour in open and controlled fermentation. J. Food Sci. Technol. 2015, 52, 6043–6048. [Google Scholar] [CrossRef]
- Tomooka, N.; Kaga, A.; Isemura, T.; Vaughan, D. Vigna. In Wild Crop Relatives: Genomic and Breeding Resources: Legume Crops and Forages; Kole, C., Ed.; Springer: Berlin/Heidelberg, Germany, 2010; pp. 291–311. [Google Scholar] [CrossRef]
- Wilkes, G.; Williams, J.T. Current status of crop plant germplasm. Crit. Rev. Plant Sci. 1983, 1, 133–181. [Google Scholar] [CrossRef]
- Benjamin, U.; Olamide, F.; Oladipupo, D.; Yusuf, A.; Abdulhakeem, A. GSC Biological and Pharmaceutical Sciences Phenotypic variability studies in selected accessions of Nigerian wild cowpea (Vigna unguiculata L. Walp). GSC Biol. Pharm. Sci. 2018, 3, 19–27. [Google Scholar]
- Popoola, J.O.; Aremu, B.R.; Daramola, F.Y.; Ejoh, A.S.; Adegbite, A.E. Morphometric Analysis of some Species in the Genus Vigna (L.) Walp: Implication for Utilization for Genetic Improvement. J. Biol. Sci. 2015, 15, 156–166. [Google Scholar] [CrossRef]
- Joshua, N.N.; Namo, O.A.T. Agronomic evaluation of some landrace cowpeas (Vigna unguiculata (L.) Walp) and their wild relative (dekindtiana var. pubescens) for incorporation into cowpea breeding programme. Eur. J. Agric. For. Res. 2019, 7, 13–23. [Google Scholar]
- Li, C.; Fatokun, C.A.; Ubi, B.; Singh, B.B.; Scoles, G.J. Determining Genetic Similarities and Relationships among Cowpea Breeding Lines and Cultivars by Microsatellite Markers. Crop. Sci. 2001, 41, 189–197. [Google Scholar] [CrossRef]
- Kouakou, C.K.; Roy-Macauley, H.; Coudou, M.; Otto, M.C.; Rami, J.-F.; Cissé, N. Diversité génétique des variétés traditionnelles de niébé [Vigna unguiculata (L.) Walp] au Sénégal: étude préliminaire. Plant Genet. Resour. Newsl. 2007, 152, 33–44. [Google Scholar]
- Badiane, F.A.; Gowda, B.S.; Cissé, N.; Diouf, D.; Sadio, O.; Timko, M.P. Genetic relationship of cowpea (Vigna unguiculata) varieties from Senegal based on SSR markers. Evolution 2012, 11, 292–304. [Google Scholar] [CrossRef] [PubMed]
- Asare, A.T.; Gowda, B.S.; Galyuon, I.K.A.; Aboagye, L.L.; Takrama, J.F.; Timko, M.P. Assessment of the genetic diversity in cowpea (Vigna unguiculata L. Walp) germplasm from Ghana using simple sequence repeat markers. Plant Genet. Resour. Charact. Util. 2010, 8, 142–150. [Google Scholar] [CrossRef]
- Manifesto, M.M.; Schlatter, A.R.; Hopp, H.E.; Suárez, E.Y.; Dubcovsky, J. Quantitative Evaluation of Genetic Diversity in Wheat Germplasm Using Molecular Markers. Crop. Sci. 2001, 41, 682–690. [Google Scholar] [CrossRef]
- Kandel, B.P.; Shrestha, J. Characterization of rice (Oryza sativa L.) germplasm in Nepal: A mini review. Farming Manag. 2018, 3, 153–159. [Google Scholar] [CrossRef]
- Krichen, L.; Audergon, J.M.; Trifi-Farah, N. Relative efficiency of morphological characters and molecular markers in the establishment of an apricot core collection. Hereditas 2012, 149, 163–172. [Google Scholar] [CrossRef]
- Lee, O.N.; Park, H.Y. Assessment of genetic diversity in cultivated radishes (Raphanus sativus) by agronomic traits and SSR markers. Sci. Hortic. 2017, 223, 19–30. [Google Scholar] [CrossRef]
- Arteaga, S.; Yabor, L.; Torres, J.; Solbes, E.; Muñoz, E.; Díez, M.J.; Vicente, O.; Boscaiu, M. Morphological and agronomic characterization of Spanish landraces of Phaseolus vulgaris L. Agriculture 2019, 9, 149. [Google Scholar] [CrossRef]
- Menssen, M.; Linde, M.; Omondi, E.O.; Abukutsa-Onyango, M.; Dinssa, F.F.; Winkelmann, T. Genetic and morphological diversity of cowpea (Vigna unguiculata (L.) Walp) entries from East Africa. Sci. Hortic. 2017, 226, 268–276. [Google Scholar] [CrossRef]
- Tanhuanpää, P.; Manninen, O. High SSR diversity but little differentiation between accessions of Nordic timothy (Phleum pratense L.). Hereditas 2012, 149, 114–127. [Google Scholar] [CrossRef] [PubMed]
- Badiane, F.A.; Diouf, M.; Diouf, D. Cowpea. In Broadening the Genetic Base of Grain Legumes; Singh, M., Bisht, I.S., Dutta, M., Eds.; Springer: New Delhi, India, 2014; pp. 95–114. [Google Scholar] [CrossRef]
- Sarr, A.; Bodian, A.; Gbedevi, K.M.; Ndir, K.N.; Ajewole, O.O.; Gueye, B.; Foncéka, D.; Diop, E.A.; Diop, B.M.; Cissé, N.; et al. Genetic Diversity and Population Structure Analyses of Wild Relatives and Cultivated Cowpea (Vigna unguiculata (L.) Walp) from Senegal Using Simple Sequence Repeat Markers. Plant Mol. Biol. Rep. 2021, 39, 112–124. [Google Scholar] [CrossRef]
- IBPGR. Descriptors For Cowpea 377. 1983. Available online: https://www.scribd.com/document/556492902/Descriptors-for-Cowpea-377 (accessed on 18 October 2023).
- IPGRI. Key Characterization and Evaluation Descriptors: Methodologies for the Assessment of 22 Crops; Bioversity International: Rome, Italy, 2011; 602p. [Google Scholar]
- R Software. The Comprehensive R Archive Network. Available online: https://cran.r-project.org (accessed on 1 April 2021).
- Oyatomi, O.; Fatokun, C.; Boukar, O.; Abberton, M.; Ilori, C.; Maxted, N.; Dulloo, M.E.; Ford-Lloyd, B.V. Screening wild Vigna species and cowpea (Vigna unguiculata) landraces for sources of resistance to Striga gesnerioides. In Enhancing Crop Genepool Use: Capturing Wild Relatives and Landrace Diversity for Crop Improvement; Maxted, N., Dulloo, M.E., Ford-Lloyd, B.V., Eds.; CABI: Boston, MA, USA, 2016; pp. 27–31. [Google Scholar] [CrossRef]
- Popoola, J.O.; Adebambo, A.; Ejoh, S.; Agre, P.; Adegbite, A.E.; Omonhinmin, C.A. Morphological diversity and cytological studies in some accessions of Vigna vexillata (L.) A. Richard. Annu. Res. Rev. Biol. 2017, 19, 1–12. [Google Scholar] [CrossRef]
- Padulosi, S.; Ng, N.Q. Origin, taxonomy, and morphology of Vigna unguiculata (L.) Walp. In Advances in Cowpea Research; Singh, B.B., Dashiell, K.E., Jackai, L.E.N., Eds.; IITA-JIRCAS: Ibadan, Nigeria, 1997; Volumes 1–12. [Google Scholar]
- Damayanti, F.; Lawn, R.J.; Bielig, L.M. Genetic compatibility among domesticated and wild accessions of the tropical tuberous legume Vigna vexillata (L.) A. Rich. Crop. Pasture Sci. 2010, 61, 785–797. [Google Scholar] [CrossRef]
- Marubodee, R.; Ogiso-Tanaka, E.; Isemura, T.; Chankaew, S.; Kaga, A.; Naito, K.; Ehara, H.; Tomooka, N. Construction of an SSR and RAD-Marker Based Molecular Linkage Map of Vigna vexillata (L.) A. Rich. PLoS ONE 2015, 10, e0138942. [Google Scholar] [CrossRef] [PubMed]
- Popoola, J.O.; Adegbite, A.E.; Obembe, O.O. Cytological studies on some accessions of African Yam Bean (AYB) (Sphenostylis stenocarpa Hochst. Ex. A. Rich. Harms). Int. Res. J. Plant Sci. 2011, 2, 249–253. [Google Scholar]
- Nicotra, A.B.; Leigh, A.; Boyce, C.K.; Jones, C.S.; Niklas, K.J.; Royer, D.L.; Tsukaya, H. The evolution and functional significance of leaf shape in the angiosperms. Funct. Plant Biol. 2011, 38, 535–552. [Google Scholar] [CrossRef]
- Moalafi, S.; Sanka, G.; Apuyor, B. Genetic diversity in cultivated cowpea (Vigna unguiculata L.). Afr. J. Agric. Sci. 2010, 32, 841–850. [Google Scholar]
- Timko, M.P.; Singh, B.B. Cowpea, a Multifunctional Legume, in Genomics of Tropical Crop Plants, 1st ed.; Moore, P.H., Ming, R., Eds.; Genomics of Tropical Crop Plants. Plant Genetics and Genomics: Crops and Models; Springer: New York, NY, USA, 2008; Volume 1, pp. 227–258. [Google Scholar] [CrossRef]
- Burger, J.C.; Chapman, M.A.; Burke, J.M. Molecular insights into the evolution of crop plants. Am. J. Bot. 2008, 95, 113–122. [Google Scholar] [CrossRef]
- Adewale, B.D.; Okonji, C.; Oyekanmi, A.A.; Akintobi, D.A.C.; Aremu, C.O. Genotypic variability and stability of some grain yield components of Cowpea. Afr. J. Agric. Res. 2010, 5, 874–880. [Google Scholar]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameters | Descriptors | |
|---|---|---|
| Qualitative traits | ||
| Hypocotyl color | Green; Purple; Others | |
| Leaf color | Pale green; Intermediate green; Dark green | |
| Leaf texture | Cariaceous; Intermediate; Membranous | |
| Growth habit | Erect; Intermediate; Prostrate; Climbing | |
| Terminal leaflet shape | Globose; Sub-globose; Sub-hastate; Hastate; Others | |
| Plant pigmentation | None; Moderate | |
| Plant hairiness | Glabrescent; Short appressed hairs | |
| Flower color | Violet; Dark blue; Yellow | |
| Pod dehiscence | No shattering; Pods opened and twisted | |
| Pod texture | Smooth; Rough | |
| Seed texture | Smooth; Rough | |
| Seed coat color | Grey; Marbled; brown; red; green; black | |
| Eye color | White; Black | |
| Seed shape | Kidney; Ovoid; Crowder; Globose; Rhomboid; Others | |
| Quantitative traits | ||
| Parameters | Code | Unit |
| Terminal leaflet length | Tlfl | cm |
| Terminal leaflet width | Tlfw | cm |
| Time to 50% flowering | T50%fw | day |
| Time to 95% pod maturity | T95%Rp | day |
| Pod length | Pdl | cm |
| Pod width | Pdw | cm |
| Number of locules per pod | Nlpd | - |
| 100-seed weight | HSdw | g |
| Factor Axes | Dim 1 | Dim 2 | Dim 3 |
|---|---|---|---|
| Variance | 0.712 | 0.517 | 0.259 |
| % of var. | 32.385 | 23.497 | 11.773 |
| Cumulative % of var. | 32.385 | 55.882 | 67.665 |
| a. Analysis of Variance for 8 Quantitative Traits in 2021 | ||||||
|---|---|---|---|---|---|---|
| 2021 | Accessions Effect | Repetition Effect | Block Effect | |||
| Traits | Mean Sq | Pr(>F) | Mean Sq | Pr(>F) | Mean Sq | Pr(>F) |
| T50%fw | 760.100 | <2 × 10−16 | 29.000 | 0.063 | 11.100 | 0.369 |
| T95% Rp | 675.800 | <2 × 10−16 | 25.500 | 0.026 | 8.900 | 0.157 |
| Tlfl | 8.446 | <2 × 10−16 | 3.319 | 1.4 × 10−6 | 0.192 | 0.577 |
| Tlfw | 1.615 | <2 × 10−16 | 0.233 | 0.029 | 0.098 | 0.064 |
| Pdl | 14.050 | <2 × 10−16 | 0.123 | 0.384 | 0.085 | 0.889 |
| Pdw | 0.005 | <2 × 10−16 | 0.000 | 0.239 | 0.000 | 0.617 |
| Nlpd | 34.890 | <2 × 10−16 | 0.990 | 0.127 | 0.390 | 0.722 |
| HSdw | 1.051 | <2 × 10−16 | 0.020 | 0.310 | 0.015 | 0.604 |
| b. Analysis of Variance for 8 Quantitative Traits in 2022 | ||||||
| 2022 | Accessions Effect | Repetition Effect | Block Effect | |||
| Traits | Mean Sq | Pr(>F) | Mean Sq | Pr(>F) | Mean Sq | Pr(>F) |
| T50%fw | 656.800 | <2 × 10−16 | 177.600 | 7.3 × 10−6 | 9.600 | 0.816 |
| T95% Rp | 252.570 | <2 × 10−16 | 115.040 | 8.1 × 10−6 | 7.870 | 0.509 |
| Tlfl | 15.773 | <2 × 10−16 | 0.535 | 0.138 | 0.140 | 0.973 |
| Tlfw | 2.844 | <2 × 10−16 | 0.204 | 0.114 | 0.079 | 0.653 |
| Pdl | 15.197 | <2 × 10−16 | 0.134 | 0.041 | 0.029 | 0.828 |
| Pdw | 0.005 | <2 × 10−16 | 0.000 | 0.225 | 0.000 | 0.442 |
| Nlpd | 42.740 | <2 × 10−16 | 0.420 | 0.056 | 0.150 | 0.362 |
| HSdw | 0.479 | <2 × 10−16 | 0.012 | 0.577 | 0.027 | 0.211 |
| p-Values for Season Effects | ||||
|---|---|---|---|---|
| Code | Trait | Season | Accession | Season × Accession |
| T50%fw | Time to 50% flowering | 0.160 | <2 × 10−16 *** | 2.77 × 10−13 *** |
| T95% Rp | Time to 95% pod maturity | 0.188 | <2 × 10−16 *** | <2 × 10−16 *** |
| Tlfl | Terminal leaflet length | 2 × 10−16 *** | <2 × 10−16 *** | 2.33 × 10−14 *** |
| Tlfw | Terminal leaflet width | <2 × 10−16 *** | <2 × 10−16 *** | 8.24 × 10−10 *** |
| Pdl | Pod length | 0.0001 *** | <2 × 10−16 *** | 1.3 × 10−12 *** |
| Pdw | Pod width | 0.332 | <2 × 10−16 *** | 0.401 |
| Nlpd | Number of locules per pod | 0.0003 *** | <2 × 10−16 *** | 2.98 × 10−7 *** |
| HSdw | Weight of 100 seeds | 0.002 ** | <2 × 10−16 *** | <2 × 10−16 *** |
| Traits | T50fw | T95Rp | HSdw | Nlpd | Pdl | Pdw | Tlfl | Tlfw |
|---|---|---|---|---|---|---|---|---|
| T50fw | ||||||||
| T95Rp | 0.000 | |||||||
| HSdw | 0.152 | 0.100 | ||||||
| Nlpd | 0.000 | 0.000 | 0.010 | |||||
| Pdl | 0.000 | 0.000 | 0.023 | 0.000 | ||||
| Pdw | 0.205 | 0.140 | 0.000 | 0.013 | 0.029 | |||
| Tlfl | 0.000 | 0.000 | 0.010 | 0.000 | 0.000 | 0.013 | ||
| Tlfw | 0.003 | 0.005 | 0.741 | 0.056 | 0.032 | 0.685 | 0.138 |
| Cluster 1 | Cluster 2 | Cluster 3 | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Traits | Mean in Category | Overall Mean | p-Value | Traits | Mean in Category | Overall Mean | p-Value | Traits | Mean in Category | Overall Mean | p-Value |
| Pdw | 0.700 | 0.405 | 0 | T50fw | 101.500 | 75.701 | 0 | Pdl | 8.495 | 7.625 | 0 |
| HSdw | 4.795 | 1.853 | 0 | T95Rp | 97.190 | 91.249 | 0.013 | Nlpd | 14.508 | 13.133 | 0 |
| Tlfw | 5.998 | 4.365 | 0.05 | Tlfw | 3.592 | 4.365 | 0.001 | Tlfl | 10.285 | 9.542 | 0 |
| Tlfl | 5.161 | 9.542 | 0.025 | Tlfl | 6.710 | 9.542 | 0 | Tlfw | 4.503 | 4.365 | 0.015 |
| Pdl | 2.485 | 7.625 | 0.009 | Nlpd | 8.214 | 13.133 | 0 | Pdw | 0.400 | 0.405 | 0.046 |
| Nlpd | 2 | 13.133 | 0 | Pdl | 4.310 | 7.625 | 0 | T95Rp | 89.719 | 91.249 | 0.007 |
| T50fw | 69.913 | 75.701 | 0 | ||||||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dramé, D.; Bodian, A.; Fonceka, D.; Tossim, H.-A.; Diangar, M.M.; Nguepjop, J.R.; Sambakhe, D.; Sidybe, M.; Diouf, D. Agro-Morphological Variability of Wild Vigna Species Collected in Senegal. Agronomy 2023, 13, 2761. https://doi.org/10.3390/agronomy13112761
Dramé D, Bodian A, Fonceka D, Tossim H-A, Diangar MM, Nguepjop JR, Sambakhe D, Sidybe M, Diouf D. Agro-Morphological Variability of Wild Vigna Species Collected in Senegal. Agronomy. 2023; 13(11):2761. https://doi.org/10.3390/agronomy13112761
Chicago/Turabian StyleDramé, Demba, Amy Bodian, Daniel Fonceka, Hodo-Abalo Tossim, Mouhamadou Moussa Diangar, Joel Romaric Nguepjop, Diarietou Sambakhe, Mamadou Sidybe, and Diaga Diouf. 2023. "Agro-Morphological Variability of Wild Vigna Species Collected in Senegal" Agronomy 13, no. 11: 2761. https://doi.org/10.3390/agronomy13112761