
Mapping and Validation of Alectra vogelii Resistance in the Cowpea Landrace B301


1
Department of Biology, University of Virginia, Charlottesville, VA 22904, USA
2
Corn, Soybean, and Wheat Quality Research Unit, United Stats Department of Agriculture–Agricultural Research Service (USDA-ARS), Wooster, OH 44691, USA
*
Author to whom correspondence should be addressed.
Agronomy 2022, 12(11), 2654; https://doi.org/10.3390/agronomy12112654
Submission received: 27 September 2022
/
Revised: 21 October 2022
/
Accepted: 25 October 2022
/
Published: 27 October 2022
(This article belongs to the Section Crop Breeding and Genetics)
Abstract
:Cowpea is the most important food legume in West and Central Africa and a valuable economic commodity in the region. Among the major biotic constraints to cowpea production are root parasitic weeds of which Alectra vogelii (Benth.) is of increasing importance. The cowpea landrace B301 was previously identified as a source of Alectra resistance, but neither the genes nor genomic loci conferring this resistance have been mapped. Therefore, to map and identify genetic markers linked to Alectra resistance for use in the molecular improvement of cowpea, we developed an F2 population from a cross of the susceptible variety 524B with B301. The population was phenotyped for resistance to A. vogelii and genotyped with a cowpea high density single nucleotide polymorphism (SNP) microarray. Putative resistance loci were mapped in F2 populations by categorical trait–multiple interval mapping and validated by selective genotyping. Selective genotyping indicated that the resistance loci on Vu04 (Rav1) and Vu11 (Rav2) were significantly associated with resistance (p ≤ 0.01). Using marker assisted backcrossing, the two resistance loci were introgressed independently into the susceptible 524B genetic background. Phenotyping and genotyping of the segregating backcross families delineated Rav1 to a 10 cM on chromosome 4 and Rav2 to a 6.7 cM interval in chromosome 11. These two loci are desirable for breeding Alectra resistant cowpea varieties due to their simple inheritance and ability to independently confer complete immunity to the parasite.
1. Introduction
Cowpea (Vigna unguiculata L. Walp.) is a legume crop that is widely grown in sub-Saharan Africa, South America, Asia, and the Southern parts of the United States and Europe [1]. Cowpea is especially valuable in West and Central Africa, where it is considered the most important food legume in part due to its high stress tolerance and ability to produce in hot and arid conditions [2]. Due to its ability to fix nitrogen, cowpea can also be grown in relatively depleted soils. In addition to its widespread use as a grain crop, cowpea pods and leaves may be consumed as green vegetables and its haulm is used as feed for cattle. Although cowpea is highly tolerant to abiotic heat and drought stresses, it is susceptible to many biotic constraints including fungi, bacteria, viruses, nematodes, insect pests, and parasitic weeds including Striga gesnerioides (Willd.) Vatke and Alectra vogelii (Benth.).
Alectra vogelii is prevalent in East and Southern Africa as well as the Guinea and Sudan Savanna regions of West Africa [1,2,3]. Alectra is particularly threatening to cowpea production in Botswana, Kenya, Tanzania, Zambia, and Zimbabwe [4,5]. In addition to cowpea, A. vogelii parasitizes groundnut, mungbean, bambara, soybean, and common bean [6,7,8,9,10,11]. Alectra infestation can result in complete yield loss, especially in conjunction with Striga infection [5,12]. Parasitism usually occurs two to three weeks prior to emergence of the parasite from the soil, during which time up to 75% of host damage occurs [1,2]. Symptoms include chlorosis, stunting, necrotic spots, leaf desiccation, and reduced flower and pod numbers. Alectra is most damaging in marginal environments with poor, sandy soil and low rainfall [1].
Alectra is difficult to control and widely dispersed by water, wind, animals, and grain distribution [13,14]. It has been suggested that Alectra plants produce even greater numbers of seed than Striga spp. and seedbanks can persist in the soil for up to 15 years [15,16]. Several races of Alectra exist throughout Africa based on the differential response of cowpea varieties to parasites collected from different regions, and there are key differences between Alectra populations from West and Southern Africa [9,17]. For instance, several cowpea lines from Botswana are resistant to Alectra from West Africa, but susceptible to Alectra from their native Botswana [17]. Variation in host virulence has been reported across species including cowpea, groundnut, mungbean, and bambara. Alectra populations collected from Mali, Nigeria, and Cameroon comprised a single race based on host virulence and differed from populations collected from Botswana, Kenya, Malawi, and South Africa. In total, five distinct A. vogelii races have been identified [9]. Several approaches have been employed to control the parasite including crop rotations, weeding, herbicide application, and by inducing suicidal germination. However, these methods are ineffective or too expensive for growers [10]. Thus, identifying and breeding new, Alectra resistant cowpea cultivars adapted to local consumer preferences is necessary to improve yields and ensure widespread production of the crop.
Several cowpea lines with resistance to Alectra have been identified and breeding lines have been developed through traditional selection techniques [4,10,16,17,18,19,20]. Among Alectra resistant cowpea lines, the landrace B301 (TVu-13950) and breeding line IT81D-994 are the most thoroughly characterized. Interesting, both B301 and IT81D-994 are also resistant to several races of Striga [21]. While no resistance genes or QTL for Alectra resistance have been mapped in cowpea, analysis of genetic segregation and allelism tests in backcross and F2 populations derived from B301 suggested the presence of duplicate dominant genes for resistance, which were designated Rav1 and Rav2 [10]. Segregation of Striga and Alectra resistance in F2 and backcross populations derived from B301 and a susceptible parent, indicated that the dominant Striga resistance gene in B301 was non-allelic and unlinked with either Alectra resistance gene [4]. Resistance to Alectra in IT81D-994, was non-allelic to the Rav1 and Rav2 genes identified in B301 by Singh et al. (1993), leading to its designation as Rav3 [21].
Previously, mapping efforts were undertaken to identify simple sequence repeat (SSR) markers linked with Alectra resistance. Bulked segregant analysis (BSA) using 20 SSR markers identified two markers on chromosome Vu03, CLM0356 and RB16, that segregated with Alectra resistance in an F2 population derived from B301 and susceptible parent, Banjar [22]. However, it is unclear how reliable or tightly linked these markers are to the resistance gene since the overall genome coverage of the 20 SSRs was low, and the phenotypic effect of the resistance locus was uncharacterized. Consequently, identifying reliable genetic markers linked to Alectra resistance genes is desirable for future breeding efforts.
To elucidate the inheritance of Alectra resistance in B301 and identify useful genetic markers for molecular breeding, we performed a comprehensive genetic analysis of an F2 mapping population segregating for Alectra resistance using the Illumina Cowpea iSelect Consortium Array [23]. Putative resistance loci were validated in a second, larger F2 population by selective genotyping of susceptible plants with polymerase chain reaction (PCR) based genetic markers. Marker assisted backcrossing (MABC) was used to incorporate the two major putative resistance loci into a uniform susceptible background for validation. We report here the identification of genomic regions associated with Alectra resistance and provide genetic markers for rapid development of high quality and Alectra resistant cowpea lines.
2. Materials and Methods
2.1. Parental Germplasm
The Alectra susceptible cowpea line 524B was used as the maternal parent and crossed with the resistant cowpea landrace, B301. 524B is a high-quality blackeye type breeding line developed at the University of California Riverside. Seed was kindly provided by Philip Roberts at the University of California Riverside. B301, a cowpea landrace collected from Botswana, has been widely used as a source of S. gesnerioides resistance in cowpea and is resistant to several Alectra races [17]. B301 was generously provided by Lucky Omoigui at the University of Agriculture, Makurdi. The F1 seed from the 524B × B301 cross were collected, planted, and the resulting plants were self-fertilized to produce two separate F2 mapping populations. Initially, 97 F2 individuals (population 1) were screened for resistance to Alectra and genotyped. The identified QTL were then validated by selective genotyping of susceptible individuals from a larger F2 population (n = 164), independently derived from the same cross (524B × B301) and referred to here as population 2.
2.2. Analysis of Alectra vogelii Resistance in Cowpea
All experiments involving viable A. vogelii seeds, developing parasites, and host–parasite interactions, were performed in an APHIS-approved quarantine facility. Pot trials were performed using previously published methods found suitable for evaluating resistance and susceptibility of cowpea to Striga or Alectra [4,10,24]. Pot trials were conducted in D15H Deepots (Stuewe & Sons Inc., Tangent, OR, USA) containing soil composed of approximately 70% sand and 30% Metro-Mix 300 (Sun Gro Horticulture Ltd., Agawam, MA, USA). The Alectra seeds were combined with fine sand (<250 microns) in a ratio approximating 2000 seeds g−1 by weight and 1 g of the mixture was added to each pot. The pots were watered every other day for seven days to maintain soil moisture and precondition the Alectra seeds. After seven days, a single cowpea seed was planted in each pot. The plants were watered to maintain soil moisture without supplemental nutrition and grown under 12 h light:12 h dark photoperiods at a constant day/night temperature of 30 °C. Cowpea plants were inspected 10 weeks after planting for the presence of Alectra plants that had emerged above the soil-line, following which each cowpea was carefully uprooted to record the presence of additional attached, but un-emerged parasites (Figure 1). Cowpea plants with visible Alectra parasitism (attachment) were considered susceptible, while plants without the presence of the parasite (emerged or un-emerged) were considered resistant.
Initially, Cowpea lines 524B, B301, and IT81D-994 were screened for resistance to Alectra populations collected from farmers’ fields in Botswana, Nigeria, and Tanzania (Table 1). IT81D-994 was obtained from Benoit Batieno (Institut de l’Environnement et Recherches Agricoles (INERA), Burkina Faso). From this analysis, the A. vogelii population collected from Kikombo in the Dodoma region of Tanzania, was selected for subsequent F2 screening experiments and mapping analysis based on its high level of virulence, good viability, and ability to overcome the Alectra resistance of IT81D-994. Each F2 screening included four replicates of each parent. The second F2 population was evaluated after eight weeks.
2.3. DNA Extraction and Genotyping
Two weeks after planting the parental and F2 cowpea, young leaf tissue samples were collected from each plant and DNA was extracted using a modified Cetyl trimethylammonium bromide (CTAB) protocol [25]. The DNA concentrations were adjusted to approximately 50 ng µL−1 and the 97 F2 plants from the first screening population and three replicates of each parent were genotyped with the Illumina Cowpea iSelect Consortium Array. The SNPs were called using a previously developed cluster file [23] and filtered in GenomeStudio 2.0 (Illumina, Inc., San Diego, CA, USA). Filtering conditions were based on guidelines from the Illumina genotyping technical notes (http://www.illumina.com/documents/products/technotes/technote_infinium_genotyping_data_analysis.pdf accessed on 27 August 2019). Polymorphic SNPs were identified, and a genetic linkage map was constructed. Chi-squared analysis was performed based on an expected F2 genotypic segregation of 1:2:1 to identify loci with significant segregation distortion for their exclusion from selective genotyping (p ≤ 0.05).
2.4. Linkage Mapping and QTL Analysis
Linkage mapping was performed using MSTmap [26]. The Kosambi mapping function was applied, and grouping was performed at p-value = 10−6. Chromosome number and orientation were assigned to correspond with the cowpea reference genome (Vigna unguiculata v1.2, NSF, UCR, USAID, DOE-JGI, http://phytozome.jgi.doe.gov/ accessed on 20 September 2022).
Using Windows QTL Cartographer, a categorical trait multiple interval mapping (CT-MIM) approach was applied [27]. The CT-MIM model, based on the categorical assignment of F2 individuals to resistant and susceptible phenotypic classes, was initially developed using default parameters: regression forward selection of markers, walk speed of 1 cM, and Bayesian information criterion (BIC): g(n) = ln(n), where g(n) is a function of sample n. Each model was refined iteratively by optimizing QTL position, searching for new QTL, and testing existing QTL based on the previously described criteria. The additive (a) and dominance (d) effects were used to calculate the degree of dominance (d/a) for each QTL [28]. Genetic linkage and QTL data were visualized using MapChart 2.3 [29].
The QTL identified in F2 population 1 were validated using a selective genotyping approach, except for a QTL detected on Vu05 due to the presence of extreme segregation distortion at the locus (X2 = 45.6) in the population, which prevented accurate validation since the assumption of equal frequencies of resistant and susceptible parental alleles in the non-selected class was violated [30,31]. SNP markers within and flanking the regions of interest were converted to PCR-based markers (Supplementary Table S1) using a three primer, allele-specific PCR (AS-PCR) assay [32]. The AS-PCR SNP identifiers correspond to those assigned by Muñoz-Amatriaín et al. (2016). Susceptible individuals in F2 population 2 were genotyped using the AS-PCR markers and the amplicons were separated by 6% (w/v) polyacrylamide gel electrophoresis (PAGE), stained with ethidium bromide, and viewed by ultraviolet light. Graphical genotypes were visualized using GGT 2 [33]. The allele frequency differences (pS–pNS) between selected susceptible individuals and the theoretical allele frequency of non-selected F2 plants were calculated, where pS is the allele frequency of the susceptible parental allele at the ith marker among selected susceptible F2 progeny and pNS is the expected allele frequency (0.5) among non-selected F2 individuals. The standard errors of marker allele frequency differences (σp) were calculated as σp = (pSqS/2NS)1/2, where qS is the frequency of the resistant parental genotype and NS is the number of susceptible F2 individuals. Absolute values of the allele frequency differences greater than 2σp or 3σp indicate significance of the resistance QTL at 95% and 99% confidence, respectively [34,35].
2.5. Marker Assisted Backcrossing and QTL Validation
To further validate the two resistance loci detected by CT-MIM and selective genotyping in a more uniform genetic background, marker assisted backcrossing was performed. 524B × B301 F1 plants were backcrossed to the susceptible parent and the subsequent generation (BC1) was genotyped with markers flanking and spanning the Vu04 (Rav1) and Vu11 (Rav2) Alectra resistance QTL (Supplementary Table S2). Individuals with the desirable resistance locus were backcrossed a second time and the process was repeated until BC3 individuals were obtained containing only the Rav1 or Rav2 resistance locus. Individuals for which recombination was detected between flanking markers were maintained and progressed to the next generation to further delineate the mapping interval. Finally, BC3 individuals were self-fertilized to develop segregating BC3S1 families. Four BC3S1 families for Rav1 (n = 60) and two BC3S1 families for Rav2 (n = 49) were evaluated for resistance or susceptibility to parasitic infection. Susceptible individuals from the Rav1 and Rav2 families were genotyped by AS-PCR with five and four markers, respectively (Supplementary Table S2).
3. Results
3.1. Phenotypic Characterization of Alectra Resistance
Cowpea cultivars 524B, B301, and IT81D-994 responded differentially to five Alectra populations collected from Botswana, Nigeria, and Tanzania, indicating the existence of at least three identifiable races of Alectra. The blackeye cultivar 524B was susceptible to all five populations tested, whereas B301 was resistant to the populations from Nigeria and Tanzania, but susceptible to the Botswanan population. IT81D-994 was resistant to all populations except the Alectra collected from Kikombo, Tanzania (Table 1). In all susceptible interactions, Alectra emergence occurred 5–10 weeks after planting. To study the inheritance of Alectra resistance in B301, we first screened an F2 population of 97 individuals derived from a cross of 524B × B301 (population 1). We subsequently phenotyped a second independently derived 524B × B301 F2 population of 164 individuals (population 2). Chi-squared tests indicated segregation did not significantly differ from an expected 15:1 ratio based upon the assumption of unlinked, duplicate dominant genes in either population (p ≤ 0.05, Table 2). All the 524B controls were susceptible to Alectra and none of the B301 plants displayed any Alectra parasitism (Table 2).
3.2. SNP Genotyping, Linkage Mapping, and QTL Analysis
To map Alectra resistance QTL, population 1 was genotyped using a cowpea high density SNP microarray. After monomorphic and low-quality SNPs were filtered out, 19,089 polymorphic markers were used for further analyses. The markers were assigned to 11 linkage groups, corresponding to the 11 cowpea chromosomes (designated Vu01–Vu11) and the total genome size was 895 cM (Supplementary Table S3). Chi-squared analysis of genotypic segregation detected the presence of segregation distortion on chromosomes Vu01, Vu03, Vu04, Vu05, Vu08, and Vu09 (p ≤ 0.05). Only the segregation distortion at the Vu05 locus corresponded with a detected Alectra resistance QTL (Table 3).
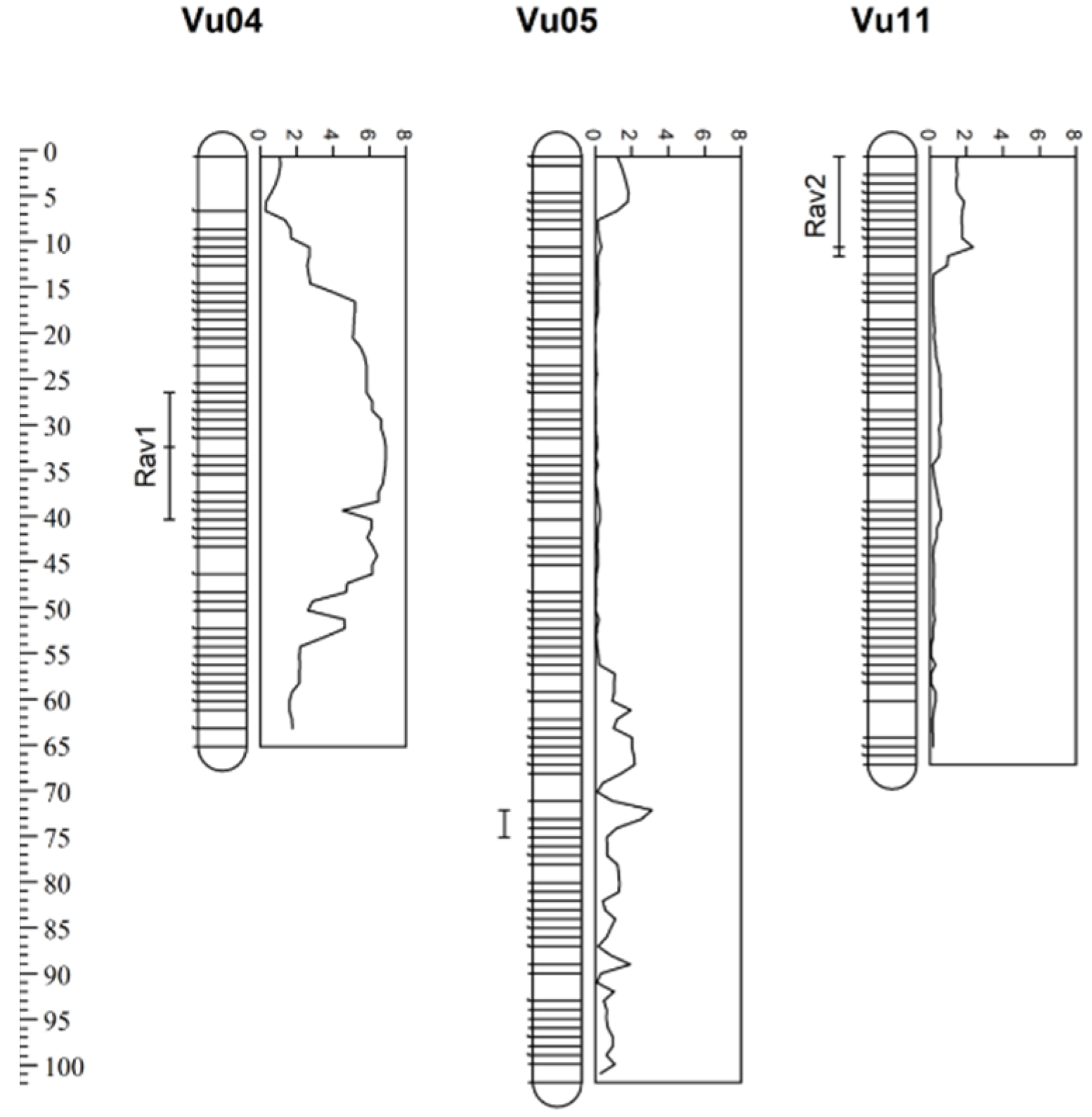
Using the CT-MIM function, three resistance QTL were identified. The first resistance locus on Vu04 was significant at an LOD of 6.9, with additive and dominance effects of 4.4 and −6.6, respectively. The QTL was slightly underdominant (d/a = −1.5). The second QTL corresponded with the locus on Vu05. The QTL was also slightly underdominant (d/a = −1.1) and the LOD was 3.1, the additive effect was 10.7, and the dominance effect was −12.2. However, none of the 97 individuals were homozygous for the 524B allele at this locus, indicating extreme segregation distortion. A third QTL was detected near the top of Vu11, which had a LOD of 2.4, additive effect of 7.5, and dominance effect of −1.3 (Table 3, Figure 2, Supplementary Figure S1). The Vu11 QTL displayed partial dominance (d/a = −0.2).
3.3. Selective Genotyping
Selective genotyping of susceptible individuals from population 2 (n = 164 individuals) validated the presence of QTL on Vu04 and Vu11. The allele frequency differences were greater than 3σp for all markers tested within the two QTL intervals. Presence of the resistant parental genotype, B301, was not detected in any of the 20 susceptible F2 plants within the Vu04 QTL, while the homozygous resistant genotype was not present among any of the susceptible individuals at the top of Vu11, which was consistent with the dominance variance detected in population 1 (Table 4, Supplementary Figure S2). The Vu05 QTL could not be validated by selective genotyping due to extreme segregation distortion favoring the homozygous resistant and heterozygous genotypes across both the resistant and susceptible classes in population 1. None of the 97 F2 individuals in population 1 genotyped with the iSelect array contained homozygous 524B SNP genotypes at the Vu05 QTL. Consequently, F2 individuals from population 2 were not genotyped with markers corresponding with this locus, and the QTL was not validated (Table 3 and Table 4).
3.4. Marker Assisted Backcrossing
Phenotypic evaluation of BC3S1 families for Alectra resistance validated the presence and functionality of Rav1 and Rav4 resistance loci. Three of the four Rav1 backcross families segregated for resistance and approximated the 3:1 segregation ratio expected with the presence of a single dominant gene based on X2 test (p < 0.05). One Rav1 BC3S1 family was fixed for the 524B susceptible alleles at the flanking SNP markers 2_07872 and 2_04604, while the other three families segregated for at least one of the two loci, indicating that the Rav1 resistance gene is present between these two markers (Supplementary Table S2). Two Rav2 BC3S1 families similarly segregated for resistance confirming that the resistance gene is located near the top of chromosome 11. Combined with the F2 mapping data, the resistance locus was delimited to a 6.7 cM interval between SNP markers 2_41050 and 2_46383 (Supplementary Table S2; Figure S2). These findings were consistent with the results obtained from CT-MIM and selective genotyping.
4. Discussion
Characterization of the underlying genetic basis of Alectra resistance in cowpea landrace B301 indicates that although multiple resistance genes are present, as was previously reported [10,21], the genetic variance is primarily explained by a dominant resistance locus on Vu04 (Rav1) and a recessive resistance locus on Vu11 (Rav2) rather than two duplicate dominant genes. The genetic complexity of Alectra resistance conferred by B301 is not unexpected. Horizontal (multigenic) resistance to parasitic weeds was previously reported for sunflower’s (Helianthus annuus) resistance response to the parasitic plant, Orobanche cumana [36,37,38].
To validate QTL that were detected by CT-MIM, a selective genotyping (i.e., trait-based analysis) approach was used. Selective genotyping is most desirable when characterizing a single trait and when phenotyping costs are significantly lower than the costs associated with genotyping a large population [31,35]. Selective genotyping confirmed the presence of A. vogelii resistance QTL on Vu04 and Vu11. However, the presence of the QTL on Vu05 could not be validated due to high segregation distortion.
The Rav1 (Vu4) and Rav2 (Vu11) resistance intervals each contain several candidate genes for Alectra resistance. Following validation and further delineation by MABC, Rav1 was delimited to a 10.3 cM interval (Supplementary Table S3). A BLASTN search of the RSG3-301 S. gesnerioides resistance gene coding sequence (CDS) against predicted cowpea CDS was performed and both Vu04 and Vu11 were hotspots for potential RSG3-301 paralogs (e-value < 4 × 10−7). Among the 38 significant blast hits, nine mapped to Vu04 and 28 mapped to Vu11. A single hit mapped to Vu03. One of the nine hits on Vu04, Vigun04g080050, mapped to the Rav1 interval. This gene is predicted to encode a disease resistance RPP13-like protein 1 NB-ARC domain-containing disease resistance protein that is a potential paralog of the S. gesnerioides resistance gene RSG3-301 (e-value ≤ 7 × 10−87) [39] (Vigna unguiculata v1.2, NSF, UCR, USAID, DOE-JGI, http://phytozome.jgi.doe.gov/ accessed on 20 September 2022). However, 937 other predicted genes fall within the Rav1 interval and additional fine mapping is needed.
The partially dominant resistance QTL on Vu11, was conservatively delineated to a 6.7 cM interval, based on the genotypic recombination of susceptible F2 plants combined with genotyping/phenotyping results obtained from BC3S1 families (Table 3, Supplementary Figure S2). Rav2 was detected by CT-MIM and validated by selective genotyping with greater than 99% confidence. None of the susceptible F2 individuals contained the homozygous susceptible genotype at the Rav2 locus, although 11 of the 20 susceptible plants were heterozygous throughout the QTL interval, indicating Rav2 did not confer complete immunity in its heterozygous state (Table 3 and Table 4, Supplementary Figure S2). The degree of dominance (d/a) was also minimal, which is consistent with the presence of a partially recessive resistance locus (Table 3). The Rav2 interval contains 231 annotated genes. No RSG3-301 paralogs were detected within this interval, although this region is tightly linked to the RSG3-301 gene and 27 potential paralogous genes [40]. Further investigation is needed to elucidate which of these genes is responsible for the Alectra resistance response of B301.
In addition to Rav1 and Rav2, initial QTL analysis suggested the presence of a resistance QTL on Vu05. However, this locus did not individually confer immunity to Alectra, as shown by the presence of susceptible F2 individuals with the homozygous B301 genotype throughout the QTL interval (Supplementary Figure S2). Extreme segregation distortion prevented validation of Vu05 QTL using a one-tailed selective genotyping approach. Since false discovery rate is influenced by segregation distortion, especially in smaller mapping populations, we do not believe Vu05 QTL is worth pursuing further and is likely a false discovery [41]. In fact, the homozygous susceptible genotype was not found among any of the 97 F2 plants at the locus, suggesting deleterious effects, unrelated to Alectra resistance, were associated with the locus.
Several BC3S1 families were developed to isolate Rav1 and Rav2 in uniform genetic backgrounds. Evaluation of the segregating families confirmed the presence of the dominant Rav1 and recessive Rav2 resistance loci and indicated they act independently of one another. By using flanking markers or markers centrally located within these genomic regions, breeders should be able to reliably select for Alectra resistance using marker assisted selection (MAS) as we have done for the BC3S1 families. No deleterious effects were detected with either Rav1 or Rav2 locus based on phenotypic observation, though further investigation is needed to confirm the absence of deleterious alleles associated with these loci. Rav1 appears especially valuable given its dominant gene action and overall effectiveness of its resistance. The AS-PCR markers reported in this study will facilitate the development of improved local varieties with resistance to Alectra and the stacking of multiple Alectra resistance genes (Supplementary Table S1). The resistance QTL reported in this study need further testing against multiple races in uniform genetic backgrounds to elucidate their overall effectiveness and determine the level and spectrum of resistance conferred by each of these genes. Unfortunately, B301 was found susceptible to some Alectra populations from its native Botswana, so additional sources of Alectra resistance must be identified and mapped (Table 1). IT81D-994 seems like a logical complementary candidate given its resistance to Alectra in Botswana.
In summary, we identified two QTL that confer resistance to Alectra in cowpea and report several genetic markers suitable for the molecular breeding of improved local lines for resistance to this noxious agricultural parasite. Based on the simple inheritance of Rav1 and Rav2 resistance loci and their ability to confer complete immunity to multiple Alectra races, these loci can be used to alleviate the impact of Alectra on cowpea production throughout Africa and can be rapidly incorporated into breeding lines by traditional or molecular breeding techniques.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/agronomy12112654/s1, Figure S1: Categorical trait multiple interval mapping (CT-MIM) of Alectra vogelii resistance in an F2 population (n = 97) generated from a cross between cowpea lines 524B and B301; Figure S2: Graphical genotypes of Alectra vogelii susceptible F2 individuals developed from a cross between 524B × B301; Table S1: Primer sequences and loci of allele-specific PCR (AS-PCR) markers used for validating Alectra vogelii resistance genes/QTL in B301; Table S2: Phenotype and Genotype of segregating BC3S1 families; Table S3: Genetic mapping data derived from the cross 524B × B301.
Author Contributions
Conceptualization, E.W.O. and M.P.T.; methodology, E.W.O. and M.P.T.; software, E.W.O.; formal analysis, E.W.O.; investigation, E.W.O.; resources, M.P.T.; writing—original draft preparation, E.W.O.; writing—review and editing, M.P.T.; visualization, E.W.O.; supervision, M.P.T.; project administration, M.P.T.; funding acquisition, M.P.T. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by a grant from the Kirkhouse Trust SCIO to MPT (GF147029). MPT was also supported in part by funds from the National Science Foundation Plant Genome Research Program (IOS-1238057).
Data Availability Statement
Not applicable.
Acknowledgments
The authors wish to thank Macsamuel Ugbaa and Lucky Omoigui (University of Makurdi, Nigeria), Beatrice Mwaipopo, and Geleta Fite (Department of Agricultural Research, Gaborone, Botswana) for providing the Alectra seed used in these analyses. Many thanks also to Matthew Tolerico for assistance with plant growth and PCR genotyping. We thank Emily Farber and Suna Onengut-Gumuscu and Stephen Rich at the Center for Public Health Genomics (University of Virginia School of Medicine, Charlottesville, VA) for assistance in running the iSelect arrays.
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
References
- Singh, B.B. Advances in Cowpea Research; IITA: Ibadan, Nigeria, 1997. [Google Scholar]
- Singh, B.B. Challenges and Opportunities for Enhancing Sustainable Cowpea Production; Fatokun, C.A., Tarawali, S.A., Singh, B.B., Kormawa, P.M., Tamo, M., Eds.; International Institute of Tropical Agriculture: Ibadan, Nigeria, 2000; pp. 154–163. [Google Scholar]
- Singh, B.B.; Olufajo, O.O.; Ishiyaku, M.E.; Adeleke, R.A.; Ajeigiie, H.A.; Mohammed, S.G. Registration of six improved germplasm lines of cowpea with combined resistance to Striga gesnerioides and Alectra vogelii. Crop Sci. 2006, 46, 2332–2333. [Google Scholar] [CrossRef] [Green Version]
- Atokple, I.D.K.; Singh, B.B.; Emechebe, A.M. Independent inheritance of Striga and Alectra resistance in cowpea genotype B301. Crop Sci. 1993, 33, 714–715. [Google Scholar] [CrossRef]
- Riches, C.R. The Biology and Control of Alectra vogelii Benth.(Scrophylariaceae) in Botswana; University of Reading: Nusajaya, Malaysia, 1988. [Google Scholar]
- Botha, P. The parasitism of Alectra vogelii Benth. with special reference to the germination of its seeds. J. S. Afr. Bot. 1948, 14, 63–80. [Google Scholar]
- Salako, E.A. Observations on the effect of Alectra vogelii infestation on the yield of groundnut. Trop. Pest Manag. 1984, 30, 209–211. [Google Scholar] [CrossRef]
- Visser, J.H.; Dorr, I.; Kollmann, R. Compatibility of Alectra vogelii with different leguminous host species. J. Plant Physiol. 1990, 135, 737–745. [Google Scholar] [CrossRef]
- Riches, C.R.; Hamilton, K.A.; Parker, C. Parasitism of grain legumes by Alectra species (Scrophulariaceae). Ann. Appl. Biol. 1992, 121, 361–370. [Google Scholar] [CrossRef]
- Singh, B.B.; Emechebe, A.M.; Atokple, I.D.K. Inheritance of Alectra resistance in cowpea genotype B301. Crop Sci. 1993, 33, 70–72. [Google Scholar] [CrossRef]
- Hussien, T.; Mishra, B.B.; Gebrekidan, H. A new parasitic weed (Alectra vogelii) similar to Striga on groundnut in Ethiopia. Trop. Sci. 2006, 46, 139–140. [Google Scholar] [CrossRef]
- Emechebe, A.; Singh, B.B.; Leleji, O.; Atokple, I.; Adu, J. Cowpea-striga problems and research in Nigeria. In Proceedings of the Combating Striga in Africa: Proceedings of the International Workshop Held in Ibadan, Ibadan, Nigeria, 22–24 August 1988; pp. 18–28. [Google Scholar]
- Parker, C.; Riches, C.R. Parasitic Weeds of the World: Biology and Control; CAB International: Wallingford, UK, 1993. [Google Scholar]
- Berner, D.K.; Cardwell, K.F.; Faturoti, B.O.; Ikie, F.O.; Williams, O.A. Relative roles of wind, crop seeds, and cattle in dispersal of Striga spp. Plant Dis. 1994, 78, 402–406. [Google Scholar] [CrossRef]
- Kroschel, J. Striga—How will it affect African agriculture in the future?—An ecological perspective. Agroecol. Plant Prot. Hum. Environ. Views Concepts 1998, 16, 137–158. [Google Scholar]
- Karanja, J.; Nguluu, S.; Wambua, J.; Gatheru, M. Response of cowpea genotypes to Alectra vogelii parasitism in Kenya. Afr. J. Biotechnol. 2013, 12, 6591–6598. [Google Scholar]
- Polniaszek, T.I.; Parker, C.; Riches, C.R. Variation in the virulence of Alectra vogelii populations on cowpea. Trop. Pest Manag. 1991, 37, 152–154. [Google Scholar] [CrossRef]
- Omoigui, L.; Kamara, A.; Ishiyaku, M.; Boukar, O. Comparative responses of cowpea breeding lines to Striga and Alectra in the dry savanna of northeast Nigeria. Afr. J. Agric. Res. 2012, 7, 747–754. [Google Scholar] [CrossRef]
- Fite, G.L.; Bruce, T.; Foyer, C.; Halford, N.; Keys, A.; Kunert, K.; Lawlor, D.; Parry, M.; Russell, G. Cowpea landraces of Botswana: A potential resistance source for Alectra vogelii. Asp. Appl. Biol. 2010, 96, 111–117. [Google Scholar]
- Kabambe, V.; Mazuma, E.; Bokosi, J.; Kazila, E. Release of cowpea line IT99K-494-6 for yield and resistance to the parasitic weed Alectra vogelii (Benth) in Malawi. Afr. J. Agric. Res. 2014, 8, 196–203. [Google Scholar]
- Atokple, I.D.K.; Singh, B.B.; Emechebe, A.M. Genetics of resistance to Striga and Alectra in cowpea. J. Hered. 1995, 86, 45–49. [Google Scholar] [CrossRef]
- Omoigui, L.; Ugba, M.; Bello, L.; Gowda, B.; Timlo, M.; Motagi, B. SSR markers linked with Alectra vogelii resistance in cowpea (Vigna unguiculata (L.). In Proceedings of the 5th International conference on Next Generation Genomics and Integrated Breeding for Crop Improvement (NGGIBCI-V), Hyderabad, India, 18–20 February 2015; ICRISAT: Patancheru, India, 2015. [Google Scholar]
- Munoz-Amatriain, M.; Mirebrahim, H.; Xu, P.; Wanamaker, S.I.; Luo, M.; Alhakami, H.; Alpert, M.; Atokple, I.; Batieno, B.J.; Boukar, O.; et al. Genome resources for climate-resilient cowpea, an essential crop for food security. Plant J. 2017, 89, 1042–1054. [Google Scholar] [CrossRef] [Green Version]
- Ohlson, E.W.; Timko, M.P. Race structure of cowpea witchweed (Striga gesnerioides) in West Africa and its implications for Striga resistance breeding of cowpea. Weed Sci. 2020, 68, 125–133. [Google Scholar] [CrossRef] [Green Version]
- Doyle, J.; Doyle, J.L. Genomic plant DNA preparation from fresh tissue-CTAB method. Phytochem. Bull. 1987, 19, 11–15. [Google Scholar]
- Wu, Y.; Bhat, P.; Close, T.J.; Lonardi, S. Efficient and accurate construction of genetic linkage maps from noisy and missing genotyping data. In International Workshop on Algorithms in Bioinformatics; Springer: Berlin/Heidelberg, Germany, 2007; pp. 395–406. [Google Scholar]
- Li, J.; Wang, S.; Zeng, Z.B. Multiple-interval mapping for ordinal traits. Genetics 2006, 173, 1649–1663. [Google Scholar] [CrossRef] [Green Version]
- deVicente, M.C.; Tanksley, S.D. QTL analysis of transgressive segregation in an interspecific tomato cross. Genetics 1993, 134, 585–596. [Google Scholar] [CrossRef] [PubMed]
- Voorrips, R.E. MapChart: Software for the graphical presentation of linkage maps and QTLs. J. Hered. 2002, 93, 77–78. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lander, E.S.; Botstein, D. Mapping mendelian factors underlying quantitative traits using RFLP linkage maps. Genetics 1989, 121, 185–199. [Google Scholar] [CrossRef] [PubMed]
- Darvasi, A.; Soller, M. Selective genotyping for determination of linkage between a marker locus and a quantitative trait locus. Theor. Appl. Genet. 1992, 85, 353–359. [Google Scholar] [CrossRef] [PubMed]
- Gaudet, M.; Fara, A.G.; Sabatti, M.; Kuztninsky, E.; Mugnozza, G.S. Single-reaction for SNP genotyping on agarose gel by allele-specific PCR in black poplar (Populus nigra L.). Plant Mol. Biol. Report. 2007, 25, 1–9. [Google Scholar] [CrossRef]
- van Berloo, R. GGT 2.0: Versatile software for visualization and analysis of genetic data. J. Hered. 2008, 99, 232–236. [Google Scholar] [CrossRef]
- Steel, R.G.D.; Torrie, J.H. Principles and Procedures of Statistics, a Biometrical Approach; McGraw-Hill Kogakusha, Ltd.: New York, NY, USA, 1980. [Google Scholar]
- Lebowitz, R.J.; Soller, M.; Beckmann, J.S. Trait-based analyses for the detection of linkage between marker loci and quantitative trait loci in crosses between inbred lines. Theor. Appl. Genet. 1987, 73, 556–562. [Google Scholar] [CrossRef]
- Molinero-Ruiz, M.; García-Ruiz, R.; Melero-Vara, J.M.; Domínguez, J. Orobanche cumana race F: Performance of resistant sunflower hybrids and aggressiveness of populations of the parasitic weed. Weed Res. 2009, 49, 469–478. [Google Scholar] [CrossRef]
- Molinero-Ruiz, M.; Melero-Vara, J.M.; García-Ruiz, R.; Domínguez, J. Pathogenic diversity within field populations of Orobanche cumana and different reactions on sunflower genotypes. Weed Res. 2006, 46, 462–469. [Google Scholar] [CrossRef]
- Molinero-Ruiz, M.; Pérez-Vich, B.; Pineda-Martos, R.; Melero-Vara, J.M. Indigenous highly virulent accessions of the sunflower root parasitic weed Orobanche cumana. Weed Res. 2008, 48, 169–178. [Google Scholar] [CrossRef]
- Altschul, S.F.; Madden, T.L.; Schaffer, A.A.; Zhang, J.; Zhang, Z.; Miller, W.; Lipman, D.J. Gapped BLAST and PSI-BLAST: A new generation of protein database search programs. Nucleic Acids Res. 1997, 25, 3389–3402. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, J.; Timko, M.P. Gene-for-gene resistance in Striga-cowpea associations. Science 2009, 325, 1094. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Wang, S.; Li, H.; Deng, Q.; Zheng, A.; Li, S.; Li, P.; Li, Z.; Wang, J. Effects of missing marker and segregation distortion on QTL mapping in F2 populations. Theor. Appl. Genet. 2010, 121, 1071–1082. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Alectra vogelii parasitism of susceptible cowpea line 524B eight weeks after planting. Shown is the growth of the parasite that occurs following successful attachment and development. The soil has been removed to allow visualization of un-emerged parasites.
Figure 1.
Alectra vogelii parasitism of susceptible cowpea line 524B eight weeks after planting. Shown is the growth of the parasite that occurs following successful attachment and development. The soil has been removed to allow visualization of un-emerged parasites.

Figure 2.
Cowpea linkage groups and Alectra vogelii resistance quantitative trait loci (QTL) with logarithm of the odds (LOD) scores. Significant QTL, Rav1 and Rav2, were identified by categorical trait multiple interval mapping (CT-MIM). Error bars indicate a 1-LOD confidence interval and scale is provided in centiMorgans (cM).
Figure 2.
Cowpea linkage groups and Alectra vogelii resistance quantitative trait loci (QTL) with logarithm of the odds (LOD) scores. Significant QTL, Rav1 and Rav2, were identified by categorical trait multiple interval mapping (CT-MIM). Error bars indicate a 1-LOD confidence interval and scale is provided in centiMorgans (cM).


{kind=link}
{kind=link}
Table 1.
Response of cowpea lines to attempted parasitism by Alectra vogelii populations from Botswana, Nigeria, and Tanzania.
Table 1.
Response of cowpea lines to attempted parasitism by Alectra vogelii populations from Botswana, Nigeria, and Tanzania.
| Country | District/State | Village | Year | 524B | B301 | IT81D-994 |
|---|---|---|---|---|---|---|
| Botswana | Marajone | 2008 | S | S | R | |
| Nigeria | Borno | Tila | 2016 | S | R | R |
| Tanzania | Dodoma | Kikombo | 2011 | S | R | S |
| Tanzania | Dodoma | Bihawana | 2011 | S | R | R |
| Tanzania | Iringa | Mkungugu | 2011 | S | R | R |
524B: Blackeye type A. vogelii susceptible cowpea breeding line; B301: Cowpea landrace collected from Botswana with known A. vogellii Stiga gesnerioides resistance; IT81D-994: Cowpea line developed by the International Institute of Tropical Agriculture with known resistance to A. vogelii and S. gesnerioides; R = Resistant; S = Susceptible.
Table 2.
Alectra vogelii resistance of cowpea lines 524B, B301, and F2 progeny (524B × B301). Chi-squared (Χ2) analysis of F2 phenotypic segregation was performed based on an assumption of duplicate dominant unlinked genes.
Table 2.
Alectra vogelii resistance of cowpea lines 524B, B301, and F2 progeny (524B × B301). Chi-squared (Χ2) analysis of F2 phenotypic segregation was performed based on an assumption of duplicate dominant unlinked genes.
| Cultivar/ Population | Resistant Plants | Susceptible Plants | Χ2 (15:1) |
|---|---|---|---|
| 524B | 0 | 8 | NA |
| B301 | 8 | 0 | NA |
| Population 1 | 88 | 9 | 1.52 |
| Population 2 | 153 | 11 | 0.06 |
NA = Not applicable.
Table 3.
Characterization of Alectra vogelii resistance quantitative trait loci (QTL). Putative QTL were identified by categorical trait multiple interval mapping (CT-MIM). The QTL intervals are delineated by a 1-logarithm of the odds (LOD) confidence interval. Positive additive effects (a) indicate increased Alectra susceptibility associated with inheritance of the susceptible parental genotype (524B). Negative dominance effects (d) indicate resistance was dominant over susceptibility. The degree of dominance (d/a) was calculated where d/a < −1.0 indicates underdominance, d/a > 1.0 indicates overdominance, and −1.0 < d/a < 1.0 indicates partial dominance.
Table 3.
Characterization of Alectra vogelii resistance quantitative trait loci (QTL). Putative QTL were identified by categorical trait multiple interval mapping (CT-MIM). The QTL intervals are delineated by a 1-logarithm of the odds (LOD) confidence interval. Positive additive effects (a) indicate increased Alectra susceptibility associated with inheritance of the susceptible parental genotype (524B). Negative dominance effects (d) indicate resistance was dominant over susceptibility. The degree of dominance (d/a) was calculated where d/a < −1.0 indicates underdominance, d/a > 1.0 indicates overdominance, and −1.0 < d/a < 1.0 indicates partial dominance.
| QTL | Chromosome | Genetic Locus | Physical Locus | LOD | aA | dB | d/aC |
|---|---|---|---|---|---|---|---|
| cM | Mbp | ||||||
| Rav1 | Vu04 | 26.2–38.3 | 4.9–40.0 | 6.9 | 4.4 | −6.6 | −1.5 |
| Vu05 D | 71.5–73.3 | 42.5–43.0 | 3.1 | 10.7 | −12.2 | −1.1 | |
| Rav2 | Vu11 | 0–10.7 | 0–6.2 | 2.4 | 7.5 | −1.3 | −0.2 |
A Additive effect; B Dominance effect; C Degree of dominance; D Extreme marker segregation distortion of 0:51:46 (524B:Heterozygous:B301).
Table 4.
Segregation of SNP markers associated with Alectra vogelii resistance in susceptible class F2 cowpea.
Table 4.
Segregation of SNP markers associated with Alectra vogelii resistance in susceptible class F2 cowpea.
| SNP A | Chromosome | Genetic Locus | Physical Locus | pp | pq | qqB | X2 (1:2:1) | pS | pS–pNSC | σp D |
|---|---|---|---|---|---|---|---|---|---|---|
| cM | Mbp | |||||||||
| 2_21345 | 4 | 6.3 | 0.9 | 6 | 5 | 0 | 7.7 * | 0.77 | 0.27 ** | 0.07 |
| 2_07872 | 4 | 30.4 | 8.5 | 11 | 0 | 0 | 22.2 ** | 1.00 | 0.50 ** | 0.00 |
| 2_23898 | 4 | 37.7 | 33.3 | 11 | 0 | 0 | 22.2 ** | 1.00 | 0.50 ** | 0.00 |
| 2_05791 | 4 | 38.7 | 34.9 | 11 | 0 | 0 | 22.2 ** | 1.00 | 0.50 ** | 0.00 |
| 2_22012 | 4 | 38.7 | 36.4 | 11 | 0 | 0 | 22.2 ** | 1.00 | 0.50 ** | 0.00 |
| 2_04604 | 4 | 40.8 | 37.1 | 11 | 0 | 0 | 22.2 ** | 1.00 | 0.50 ** | 0.00 |
| 2_04705 | 4 | 46.0 | 38.8 | 11 | 0 | 0 | 22.2 ** | 1.00 | 0.50 ** | 0.00 |
| 2_16297 | 4 | 53.8 | 40.2 | 9 | 2 | 0 | 14.6 ** | 0.91 | 0.41 ** | 0.05 |
| 2_14455 | 11 | 4.6 | 0.8 | 5 | 6 | 0 | 6.6 * | 0.73 | 0.23 ** | 0.07 |
| 2_54689 | 11 | 6.7 | 3.0 | 5 | 6 | 0 | 6.6 * | 0.73 | 0.23 ** | 0.07 |
| 2_41050 | 11 | 7.2 | 3.4 | 5 | 6 | 0 | 6.6 * | 0.73 | 0.23 ** | 0.07 |
| 2_00054 | 11 | 28.4 | 30.4 | 1 | 10 | 0 | 8.2 * | 0.55 | 0.05 | 0.08 |
A SNP (single nucleotide polymorphism) identifier from Muñoz-Amatriaín et al. 2016. B pp, homozygous for 524B alleles, pq heterozygous, qq homozygous for B301 alleles; C Allele frequency differences between A. vogelii susceptible (pS) and not selected (pNS) F2 cowpea; D Standard error of the allele frequency difference; * p ≤ 0.05; ** p ≤ 0.01.
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Ohlson, E.W.; Timko, M.P. Mapping and Validation of Alectra vogelii Resistance in the Cowpea Landrace B301. Agronomy 2022, 12, 2654. https://doi.org/10.3390/agronomy12112654
AMA Style
Ohlson EW, Timko MP. Mapping and Validation of Alectra vogelii Resistance in the Cowpea Landrace B301. Agronomy. 2022; 12(11):2654. https://doi.org/10.3390/agronomy12112654
Chicago/Turabian StyleOhlson, Erik W., and Michael P. Timko. 2022. "Mapping and Validation of Alectra vogelii Resistance in the Cowpea Landrace B301" Agronomy 12, no. 11: 2654. https://doi.org/10.3390/agronomy12112654
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.