
Efficacy of Botanical Extract Formulations of Zanthroxylum usambarense and Warburgia ugandensis on Post-Harvest Management of Sitophilus zeamais in Maize
 ,
,  , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Identification of Sitophilus zeamais
2.2. Preparation of Maize Grains, Sitophilus zeamais Colony and Organic Plant Extracts
2.3. Determination of Plant Extracts Lethal Dose and Repellent Activities
2.3.1. Bioassays with Solvent Extracts
2.3.2. Contact Toxicity Bioassay of Plant Extracts
2.4. Phytochemicals Detection Using Gas Chromatography and Mass Spectrometry
2.5. Bioassays with Extract Formulation on Sitophilus zeamais
2.5.1. Plant Extract Formulation Repellent Activity
2.5.2. Plant Extract Formulation Mortality Assessment under Laboratory Conditions
2.5.3. Long-Term Evaluation of Extract Formulation Efficacy on Sitophilus zeamais in Stored Maize
2.6. Prediction of Active Compounds’ Molecular Targets via Molecular Docking
2.6.1. Sources of Data on Ligands and Targets
2.6.2. Prediction of Active Sites and Molecular Docking Simulation
2.7. Data Analysis
3. Results
3.1. Molecular Identification of Sitophilus zeamais
3.2. Qualitative Phytochemical Characterization Analysis Using GC-MS
3.3. Repellent Activity of Organic Extracts of Zanthroxylum usambarense and Warburgia ugandensis on Sitophilus zeamais
3.4. Mortality Activity of Hexane and Methanol Extracts of Zanthroxylum usambarense and Warburgia ugandensis on Sitophilus zeamais
3.5. Probit Analysis of Zanthroxylum usambarense and Warburgia ugandensis Extracts’ Mortality on Sitophilus zeamais
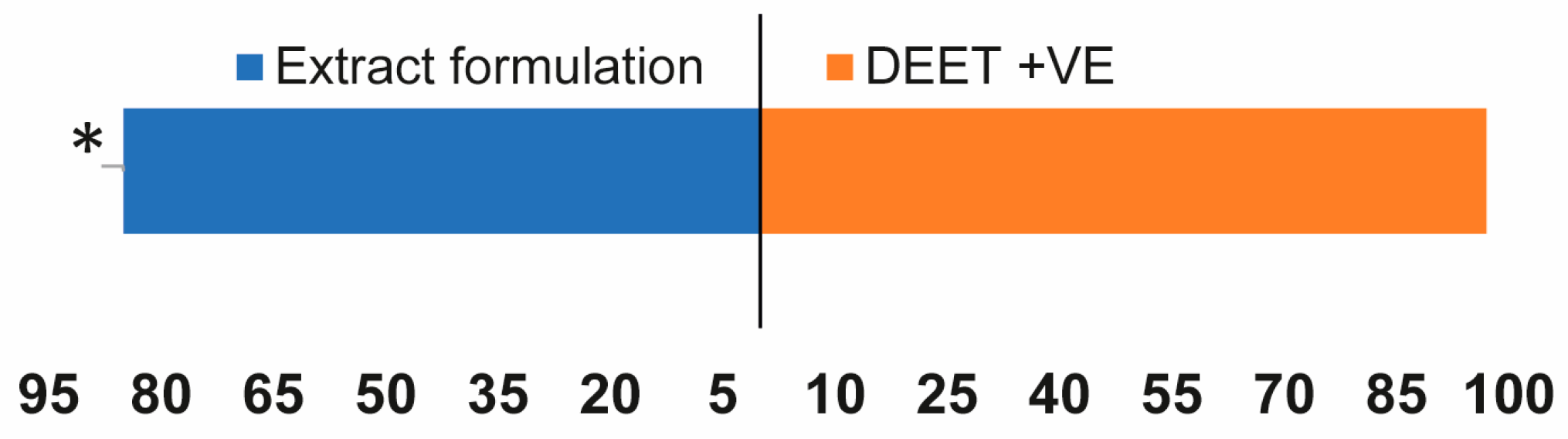
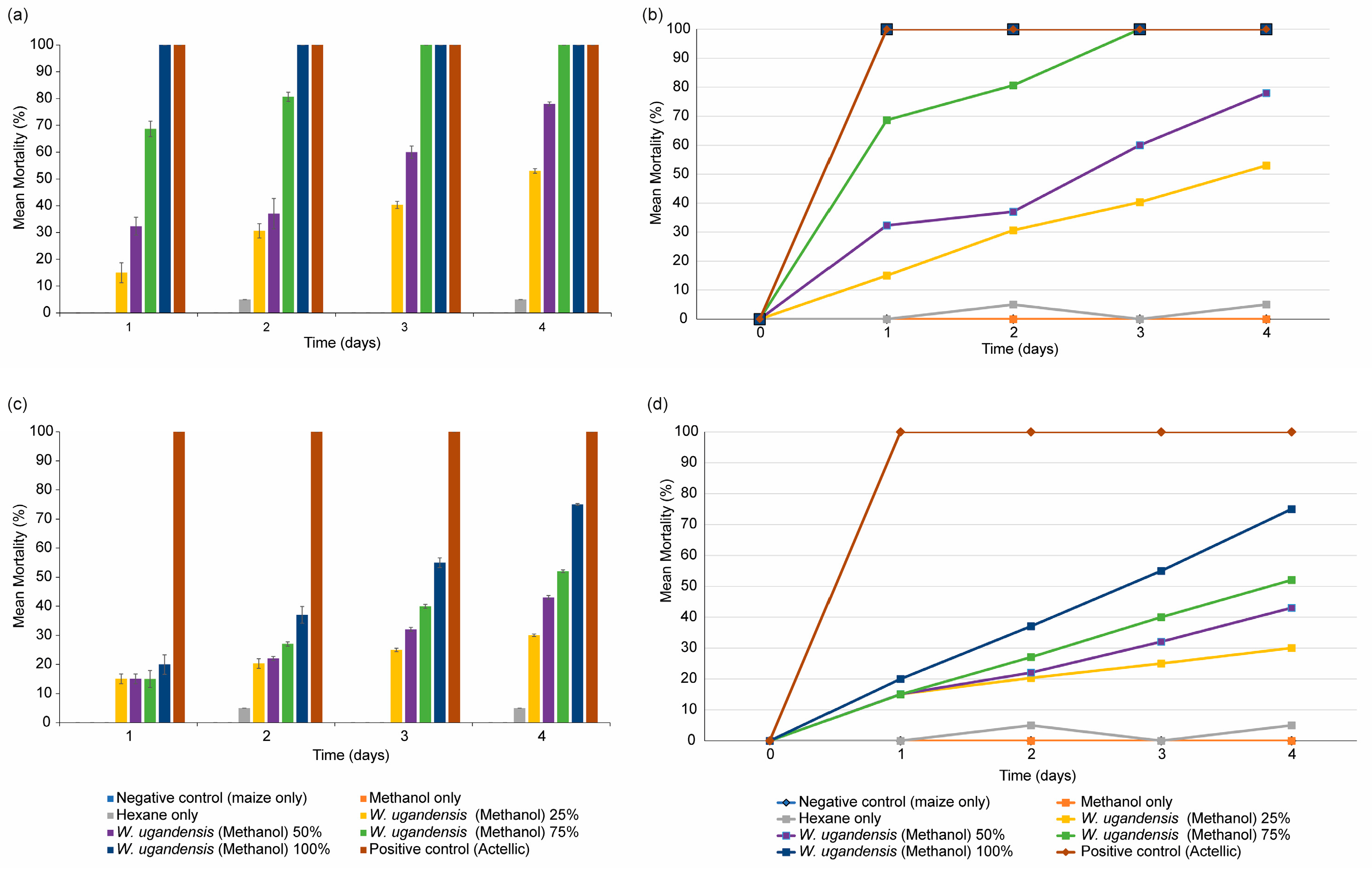
3.6. Insecticidal Activity of Extracts Formulation on Sitophilus zeamais in Maize Grains under Laboratory Conditions
3.7. Extract Formulation Insecticidal Activity on S. zeamais in Maize Grains Stored in a Granary for Six Months
3.8. Analysis of Molecular Targets for Repellent and Toxic Bioactive Compounds in Sitophilus zeamais
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Strategic-Plan-KALRO-2017-2021. Available online: https://www.kalro.org/download/kalro-strategic-plan-2017-2021/ (accessed on 11 May 2023).
- World Food and Agriculture—Statistical Yearbook 2021; FAO: Roman, Italy, 2021. [CrossRef]
- Sisay, B.; Tefera, T.; Wakgari, M.; Ayalew, G.; Mendesil, E. The efficacy of selected synthetic insecticides and botanicals against fall armyworm, Spodoptera frugiperda, in maize. Insects 2019, 10, 45. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, F.; Huang, J.; Qi, J.; Li, Q.; Wu, H.; Ju, J. Proteomic analysis of antifungal mechanism of star anise essential oil against Aspergillus niger and its application potential in prolonging bread shelf life. LWT 2022, 169, 114023. [Google Scholar] [CrossRef]
- Tudi, M. Agriculture development, pesticide application and its impact on the environment. Int. J. Environ. Res. Public Health 2021, 18, 1112. [Google Scholar] [CrossRef] [PubMed]
- Francikowski, J.; Baran, B.; Cup, M.; Janiec, J.; Krzyżowski, M. Commercially available essential oil formulas as repellents against the stored-product pest Alphitobius diaperinus. Insects 2019, 10, 96. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mladenović, M. The targeted pesticides as acetylcholinesterase inhibitors: Comprehensive cross-organism molecular modelling studies performed to anticipate the pharmacology of harmfulness to humans in vitro. Molecules 2018, 23, 2192. [Google Scholar] [CrossRef] [Green Version]
- Kita, T.; Mino, H.; Ozoe, F.; Ozoe, Y. Spatiotemporally different expression of alternatively spliced GABA receptor subunit transcripts in the housefly Musca domestica. Arch. Insect Biochem. Physiol. 2019, 101, e21541. [Google Scholar] [CrossRef]
- Shao, H.; Zhang, Y. Non-target effects on soil microbial parameters of the synthetic pesticide carbendazim with the biopesticides cantharidin and norcantharidin. Sci. Rep. 2017, 7, 5521. [Google Scholar] [CrossRef] [Green Version]
- Souto, A.L.; Sylvestre, M.; Tölke, E.D.; Tavares, J.F.; Barbosa-Filho, J.M.; Cebrián-Torrejón, G. Plant-derived pesticides as an alternative to pest management and sustainable agricultural production: Prospects, applications and challenges. Molecules 2021, 26, 4835. [Google Scholar] [CrossRef]
- Wink, M. Modes of Action of Herbal Medicines and Plant Secondary Metabolites. Medicines 2015, 2, 251–286. [Google Scholar] [CrossRef]
- Suleiman, M.; Ibrahim, N.D.; Majeed, Q. Control of Sitophilus zeamais (Motsch) [Coleoptera: Curculionidae] on Sorghum Using Some Plant Powders. Int. J. Agric. For. 2012, 2, 53–57. [Google Scholar] [CrossRef] [Green Version]
- He, W. Chemical constituents and biological activities of Zanthoxylum usambarense. Phytother. Res. 2002, 16, 66–70. [Google Scholar] [CrossRef] [PubMed]
- Rinkevich, F.D.; Bourgeois, L. In silico identification and assessment of insecticide target sites in the genome of the small hive beetle, Aethina tumida. BMC Genom. 2020, 21, 154. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Isman, M.B. Botanical insecticides, deterrents, and repellents in modern agriculture and an increasingly regulated world. Annu. Rev. Entomol. 2006, 51, 45–66. [Google Scholar] [CrossRef] [Green Version]
- Kinyanjui, G.; Khamis, F.M.; Ombura, F.L.O.; Kenya, E.U.; Ekesi, S.; Mohamed, S.A. Infestation Levels and Molecular Identification Based on Mitochondrial COI Barcode Region of Five Invasive Gelechiidae Pest Species in Kenya. J. Econ. Entomol. 2019, 112, 872–882. [Google Scholar] [CrossRef] [PubMed]
- Mutungi, C.; Gaspar, A. Maize Quality Standards and Specifications A Trainer’s Manual for Smallholder Farmers in Tanzania Maize Quality Standards and Specifications: A Trainer’s Manual for Smallholder Farm... Book · October 2020, Cassava Project View Project Hermetic Storage Technologies View Project”. [Online]. Available online: https://www.researchgate.net/publication/344645548 (accessed on 11 May 2023).
- Law-Ogbomo, K. Law-Ogbomo and Ogedegbe the Influence of Sett Size on Growth and Fresh Yield of Ipomeabatatas L. Journal of Applied Agricultural Research View Project Agronomic Response of Watermelon (Citrulus lunatus (thumb) Matsum and nakai) to Plant Population and Fertilizer Types in Low Fertile Soil Environment View Project. 2019. [Online]. Available online: https://www.researchgate.net/publication/334291027 (accessed on 11 May 2023).
- Zhang, Q.W.; Lin, L.G.; Ye, W.C. Techniques for extraction and isolation of natural products: A comprehensive review. Chin. Med. 2018, 13, 20. [Google Scholar] [CrossRef] [Green Version]
- World Health Organization. Standard Operating Procedure for Testing Insecticide Susceptibility of Adult Mosquitoes in WHO Bottle Bioassays; World Health Organization: Geneva, Switzerland, 2022. [Google Scholar]
- Obeng-Ofori, D.; Reichmuth, C.; Bekele, J.; Hassanali, A. Biological activity of 1,8 cineole, a major component of essential oil of Ocimum kenyense (Ayobangira) against stored product beetles. J. Appl. Entomol. 1997, 121, 237–243. [Google Scholar] [CrossRef]
- Takakura, K.I. Reconsiderations on evaluating methodology of repellent effects: Validation of indices and statistical analyses. J. Econ. Entomol. 2009, 102, 1977–1984. [Google Scholar] [CrossRef]
- Don-Pedro, K.N. Investigation of Single and Joint Fumigant Insecticidal Action of Citruspeel Oil Components. Pestic. Sci. 1996, 46, 79–84. [Google Scholar] [CrossRef]
- Yakubu, O.E.; Otitoju, O.; Onwuka, J. Gas Chromatography-Mass Spectrometry (GC-MS) Analysis of Aqueous Extract of Daniellia oliveri Stem Bark. Pharm. Anal. Acta 2017, 8, 1000568. [Google Scholar] [CrossRef]
- Fleming, R.; Retnakaran, A. Evaluating Single Treatment Data Using Abbott’s Formula with Reference to Insecticides. J. Econ. Entomol. 1985, 78, 1179–1181. [Google Scholar] [CrossRef]
- The State of Food and Agriculture 2021; FAO: Rome, Italy, 2021. [CrossRef]
- Ileke, K.D.; Idoko, J.E.; Ojo, D.O.; Adesina, B.C. Evaluation of botanical powders and extracts from Nigerian plants as protectants of maize grains against maize weevil, Sitophilus zeamais (Motschulsky) [Coleoptera: Curculionidae]. Biocatal. Agric. Biotechnol. 2020, 27, 101702. [Google Scholar] [CrossRef] [PubMed]
- Douglas, A.; Roos, D.; Mancini, F.; Couto, A.; Lusseau, D. An Introduction to R Software. Available online: https://intro2r.com/index.html (accessed on 11 May 2023).
- Otusanya, O.; Ilori, O. Phytochemical Screening and the Phytotoxic Effects of Aqueous Extracts of Tithonia diversifolia (Hemsl) A. Gray. Int. J. Biol. 2012, 4, 54. [Google Scholar] [CrossRef] [Green Version]
- Acero, L.H. Insecticidal Property of Jackfruit (Artocarpus heterophyllus) Peel Ethanol Extract against Rice Weevils (Sitophilus oryzae). Int. J. Biosci. Biochem. Bioinform. 2019, 9, 158–165. [Google Scholar] [CrossRef] [Green Version]
- Rajashekar, Y.; Bakthavatsalam, N.; Shivanandappa, T. Botanicals as grain protectants. Psyche 2012, 2012, 646740. [Google Scholar] [CrossRef]
- Liao, M. Transcriptomic analysis of Sitophilus zeamais in response to limonene fumigation. Pest Manag. Sci. 2022, 78, 4774–4782. [Google Scholar] [CrossRef]
- Ismail, M.S.M.; Tag, H.M.; Rizk, M.A. Acaricidal, ovicidal, and repellent effects of Tagetes patula leaf extract against Tetranychus urticae Koch (Acari: Tetranychidae). J. Plant Prot. Res. 2019, 59, 151–159. [Google Scholar] [CrossRef]
- Kosini, D.; Nukenine, E.N.; Agbor, G.A.; Tchinda, A.T.; Abdou, J.P.; Yaya, J.A.G.; Kowa, T.K. Fractionated Extracts From Gnidia kraussiana (Malvales: Thymeleaceae) as Bioactive Phytochemicals for Effective Management of Callosobruchus maculatus (Coleoptera: Chrysomelidae) in Stored Vigna unguiculata (Fabales: Fabaceae) Seeds. J. Insect. Sci. 2021, 21, 14. [Google Scholar] [CrossRef] [PubMed]
- Jerotich, A.; Michura, G. Synergistic Qualities of Selected Plant Extracts on Potency of Pyrethrins against the Maize Weevil, Sitophilus zeamais (Motsch.) (Coleoptera: Curculionidae). Ph.D. Thesis, Chuka University, Embu, Kenya, 2020. [Google Scholar]
- Zhang, H.; Chen, J.L.; Lin, J.H.; Lin, J.T.; Wu, Z.Z. Odorant-binding proteins and chemosensory proteins potentially involved in host plant recognition in the Asian citrus psyllid, Diaphorina citri. Pest Manag. Sci. 2020, 76, 2609–2618. [Google Scholar] [CrossRef]
- Ileke, K.D.; Ogungbite, O.C.; Olayinka-Olagunju, J.O. Powders and extracts of Syzygium aromaticum and Anacardium occidentale as entomocides against the infestation of Sitophilus oryzae (L.) [Coleoptera: Curculionidae] on stored sorghum grains. Afr. Crop Sci. J. 2014, 22, 267–273. [Google Scholar]
- Zhang, J.-W.; Li, B.-Y.; Lu, X.-X.; Zheng, Y.; Wang, D.; Zhang, Z.; Zeng, D.; Du, S.-S. Chemical Diversity and Anti-Insect Activity Evaluation of Essential Oils Extracted from Five Artemisia Species. Plants 2022, 11, 1627. [Google Scholar] [CrossRef]
- Ouko, R.O.; Koech, S.C.; Arika, W.M.; Osano, K.O.; Ogola, P.E.; Oduor, R.O.; Ngugi, N.P. Bioefficacy of organic extracts of A. sativum against S. zeamais (Coleoptera; Dryophthoridae). Biol. Syst. Open Access 2017, 6, 174. [Google Scholar] [CrossRef] [Green Version]
- Ikawati, S.; Himawan, T.; Abadi, A.L.; Tarno, H.; Fajarudin, A. In Silico Study of Eugenol and Trans-Caryophyllene also Clove Oil Fumigant Toxicity on Tribolium castaneum. J. Trop. Life Sci. 2022, 12, 339–349. [Google Scholar] [CrossRef]
- Ayalew, A.A. Insecticidal activity of Lantana camara extract oil on controlling maize grain weevils. Toxicol. Res. Appl. 2020, 4, 239784732090649. [Google Scholar] [CrossRef] [Green Version]
- Opiyo, S.A. Evaluation of Warburgia ugandensis Extracts and Compounds for Crop Protection against Prostephanus truncates. Adv. Anal. Chem. 2020, 10, 15–19. [Google Scholar] [CrossRef]
- De Bruyne, M.; Warr, C.G. Molecular and cellular organization of insect chemosensory neurons. BioEssays 2006, 28, 23–34. [Google Scholar] [CrossRef] [PubMed]
- Liao, M.; Yang, Q.; Xiao, J.; Huang, Y.; Zhou, L.; Hua, R.; Cao, H. Toxicity of Melaleuca alternifolia essential oil to the mitochondrion and NAD+/NADH dehydrogenase in Tribolium confusum. PeerJ 2018, 6, e5693. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, H.M.; Kwon, H.; Kim, K.; Lee, S.E. Antifungal and antiaflatoxigenic activities of 1,8-Cineole and t-Cinnamaldehyde on Aspergillus flavus. Appl. Sci. 2018, 8, 1655. [Google Scholar] [CrossRef] [Green Version]
- Silver, K.S.; Du, Y.; Nomura, Y.; Oliveira, E.E.; Salgado, V.L.; Zhorov, B.S.; Dong, K. Voltage-gated sodium channels as insecticide targets. Adv. Insect Physiol. 2014, 46, 389–433. [Google Scholar] [CrossRef]
- Niroumand, M.C. An evidence-based review on medicinal plants used as insecticide and insect repellent in traditional Iranian medicine. Iran. Red Crescent Med. J. 2016, 18, e22361. [Google Scholar] [CrossRef] [Green Version]
- Akwatulira, F. Vegetative propagation of Warburgia ugandensis Sprague: An important medicinal tree species in eastern Africa. J. Med. Plants Res. 2011, 5, 6615–6621. [Google Scholar] [CrossRef]
- Mahmood, I.; Imadi, S.R.; Shazadi, K.; Gul, A.; Hakeem, K.R. Effects of pesticides on environment. In Plant, Soil and Microbes: Volume 1: Implications in Crop Science; Springer International Publishing: Berlin/Heidelberg, Germany, 2016; pp. 253–269. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | Hexane | Methanol |
|---|---|---|
| W. ugandensis | 69.25 μg/mL ± 7.16 | 163.52 μg/mL ± 12.96 |
| Z. usambarense | 114.89 g/mL ± 9.06 | 197.19 g/mL ± 19.09 |
| Actellic superTM | 27.03 g/mL ± 4.83 | 42.13 g/mL ± 10.83 |
| Compound/Molecular Formula | Target Proteins | Binding Energy (ΔG) (kcal/mol) |
|---|---|---|
| D-Limonene | Glutathione-s-transferase epsilon 2 (GSTe2) | −13.49 |
| (C10H16) | ||
| Eugenol | Cytochrome c oxidase subunit 2 | −11.13 |
| (C10H12O2) | ||
| Terpinen-4-ol | Phenoloxidase-activating factor 2 | −9.97 |
| (C10H18O) | ||
| 1,8-cineole | Mitochondrial, Calcium uniporter protein | −7.15 |
| (C12H20O3) | ||
| Linalool | Cathepsin L-like proteinases | −7.00 |
| (C10H18O2) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mpoke, R.S.; Kinyua, J.; Kimani, J.W.; Ajene, I.J.; Likhayo, P.; Khamis, F.M. Efficacy of Botanical Extract Formulations of Zanthroxylum usambarense and Warburgia ugandensis on Post-Harvest Management of Sitophilus zeamais in Maize. Sustainability 2023, 15, 10833. https://doi.org/10.3390/su151410833
Mpoke RS, Kinyua J, Kimani JW, Ajene IJ, Likhayo P, Khamis FM. Efficacy of Botanical Extract Formulations of Zanthroxylum usambarense and Warburgia ugandensis on Post-Harvest Management of Sitophilus zeamais in Maize. Sustainability. 2023; 15(14):10833. https://doi.org/10.3390/su151410833
Chicago/Turabian StyleMpoke, Rhonest Siroya, Johnson Kinyua, Josephine Wambaire Kimani, Inusa Jacob Ajene, Paddy Likhayo, and Fathiya Mbarak Khamis. 2023. "Efficacy of Botanical Extract Formulations of Zanthroxylum usambarense and Warburgia ugandensis on Post-Harvest Management of Sitophilus zeamais in Maize" Sustainability 15, no. 14: 10833. https://doi.org/10.3390/su151410833