
DNA Barcodes for Accurate Identification of Selected Medicinal Plants (Caryophyllales): Toward Barcoding Flowering Plants of the United Arab Emirates

 ,
,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Specimen Collection and Data Acquisition
2.2. Tissue Sampling and DNA Extraction
2.3. PCR Amplification and Purification
2.4. Sequence Analysis
3. Results
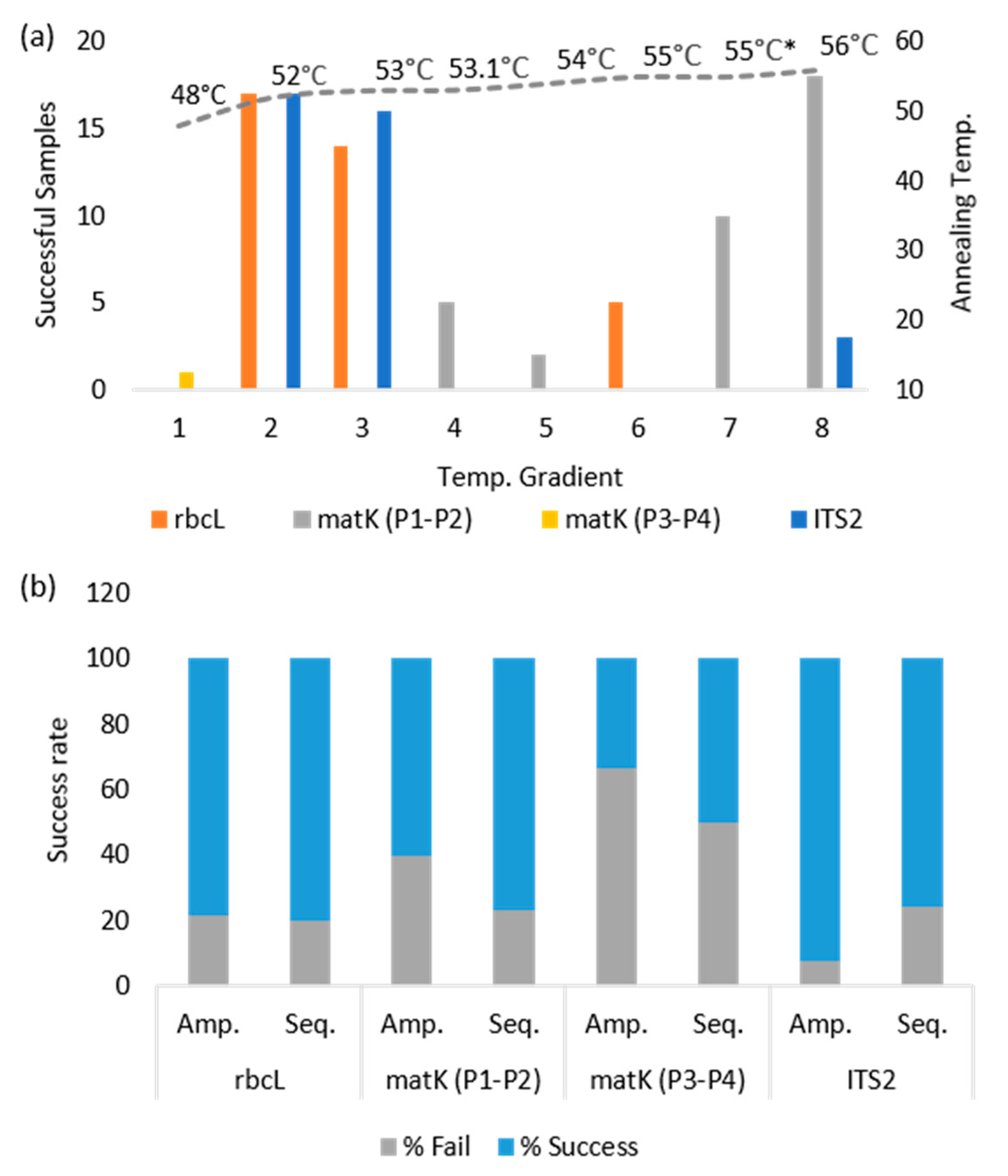
3.1. Barcode Amplification and Sequencing Success
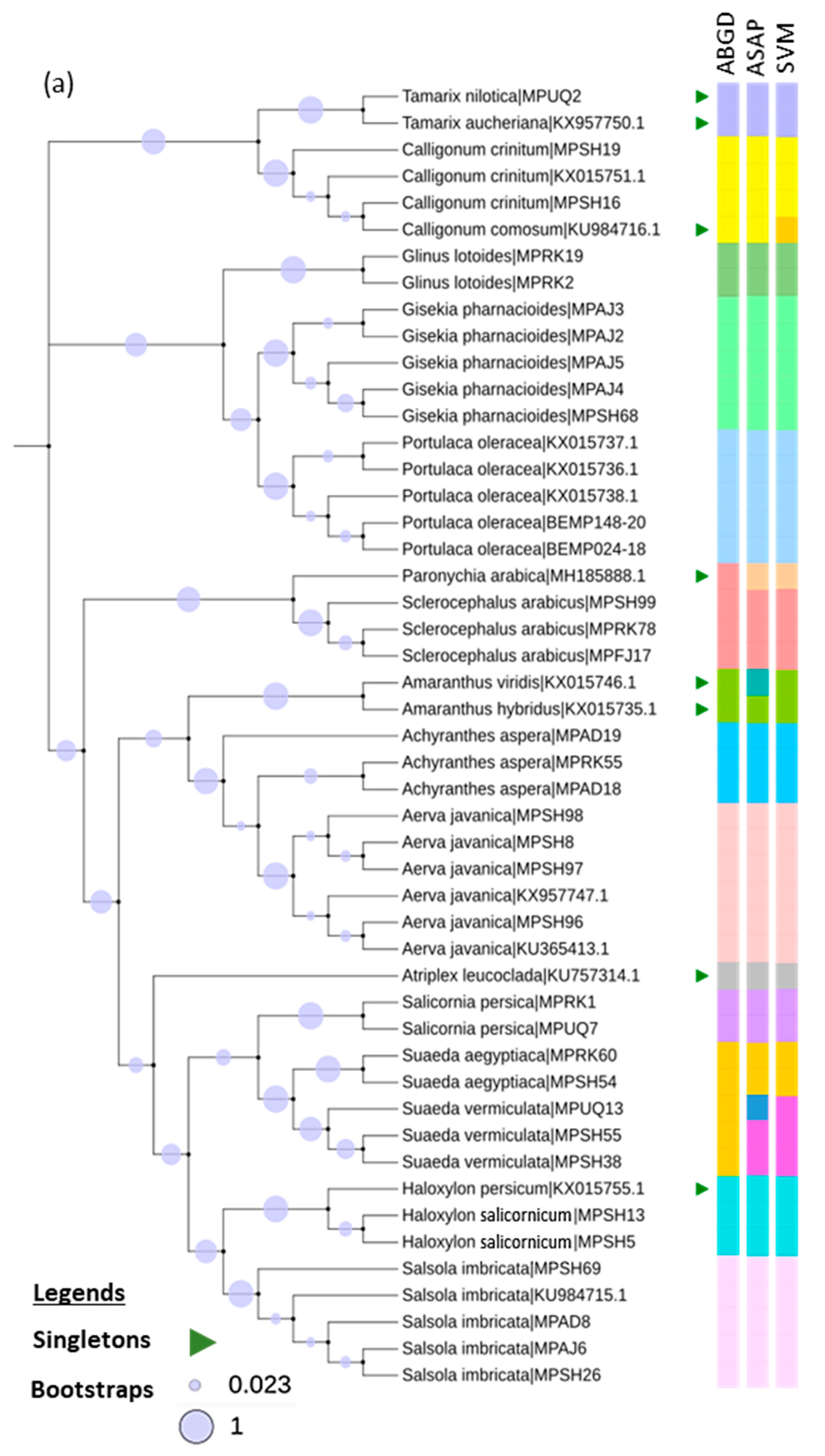
3.2. Taxonomic Assignment
3.3. Genetic Divergence
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Singh, A.; Singh, S.; Mohan Prasad, S. Role of Medicinal Plants for Health Perspective: Special Reference to Antioxidant Potential. J. Chem. Biol. Ther. 2016, 1, 106. [Google Scholar] [CrossRef]
- Hostettmann, K.; Marston, A.; Ndjoko, K.; Wolfender, J.-L. The Potential of African Plants as a Source of Drugs. Curr. Org. Chem. 2005, 4, 973–1010. [Google Scholar] [CrossRef]
- Ljubuncic, P.; Azaizeh, H.; Portnaya, I.; Cogan, U.; Said, O.; Saleh, K.A.; Bomzon, A. Antioxidant activity and cytotoxicity of eight plants used in traditional Arab medicine in Israel. J. Ethnopharmacol. 2005, 99, 43–47. [Google Scholar] [CrossRef] [PubMed]
- Cope, T.A.; Miller, A.G.; Morris, M. Plants of Dhofar: The Southern Region of Oman. Traditional, Economic and Medicinal Uses. Geogr. J. 1990, 156, 89. [Google Scholar] [CrossRef]
- Ghazanfar, S.A. Handbook of Arabian Medicinal Plants; CRC Press: Boca Raton, FL, USA, 1994. [Google Scholar]
- Bussmann, R.W.; Malca, G.; Glenn, A.; Sharon, D.; Nilsen, B.; Parris, B.; Dubose, D.; Ruiz, D.; Saleda, J.; Martinez, M.; et al. Toxicity of medicinal plants used in traditional medicine in Northern Peru. J. Ethnopharmacol. 2011, 137, 121–140. [Google Scholar] [CrossRef] [Green Version]
- Chandra, S.; Rawat, D.S. Medicinal plants of the family Caryophyllaceae: A review of ethno-medicinal uses and pharmacological properties. Integr. Med. Res. 2015, 4, 123–131. [Google Scholar] [CrossRef] [Green Version]
- Phondani, P.C.; Bhatt, A.; Elsarrag, E.; Horr, Y.A. Ethnobotanical magnitude towards sustainable utilization of wild foliage in Arabian Desert. J. Tradit. Complement. Med. 2016, 6, 209–218. [Google Scholar] [CrossRef] [Green Version]
- Ullah, R.; Alqahtani, A.S.; Noman, O.M.A.; Alqahtani, A.M.; Ibenmoussa, S.; Bourhia, M. A review on ethno-medicinal plants used in traditional medicine in the Kingdom of Saudi Arabia. Saudi J. Biol. Sci. 2020, 27, 2706–2718. [Google Scholar] [CrossRef]
- Aati, H.; El-Gamal, A.; Shaheen, H.; Kayser, O. Traditional use of ethnomedicinal native plants in the Kingdom of Saudi Arabia. J. Ethnobiol. Ethnomed. 2019, 15, 2. [Google Scholar] [CrossRef]
- Sakkir, S. Medicinal plants diversity and their conservation status in the United Arab Emirates (UAE). J. Med. Plants Res. 2012, 6, 1304–1322. [Google Scholar] [CrossRef]
- Cybulska, I.; Brudecki, G.; Alassali, A.; Thomsen, M.; Jed Brown, J. Phytochemical composition of some common coastal halophytes of the United Arab Emirates. Emirates J. Food Agric. 2014, 26, 1046–1056. [Google Scholar] [CrossRef] [Green Version]
- Sajjad, A.; Syed, A.; Hasnain, A.; Mohamed, E. Ethno Botanical Study of Traditional Native Plants in Al Ain UAE. Int. J. Adv. Res. Biol. Sci. 2017, 4, 1–10. [Google Scholar] [CrossRef]
- Parvathy, V.A.; Swetha, V.P.; Sheeja, T.E.; Sasikumar, B. Detection of plant-based adulterants in turmeric powder using DNA barcoding. Pharm. Biol. 2015, 53, 1774–1779. [Google Scholar] [CrossRef]
- Mosa, K.A.; Soliman, S.; El-Keblawy, A.; Ali, M.A.; Hassan, H.A.; Tamim, A.A.B.; Al-Ali, M.M. Using DNA Barcoding to Detect Adulteration in Different Herbal Plant-Based Products in the United Arab Emirates: Proof of Concept and Validation. Recent Pat. Food. Nutr. Agric. 2018, 9, 55–64. [Google Scholar] [CrossRef]
- Abdel-Aziz, S.M.; Aeron, A.; Kahil, T.A.; Abdel-Aziz, S.M.; Kahil, T.A.; Aeron, A. Health Benefits and Possible Risks of Herbal Medicine. In Microbes in Food and Health; Garg, N., Abdel-Aziz, S., Aeron, A., Eds.; Springer: Cham, Switzerland, 2016. [Google Scholar] [CrossRef]
- Pratiwi, R.; Dipadharma, R.H.F.; Prayugo, I.J.; Layandro, O.A. Recent Analytical Method for Detection of Chemical Adulterants in Herbal Medicine. Molecules 2021, 26, 6606. [Google Scholar] [CrossRef]
- Tungmunnithum, D.; Renouard, S.; Drouet, S.; Blondeau, J.P.; Hano, C. A Critical Cross-Species Comparison of Pollen from Nelumbo nucifera Gaertn. vs. Nymphaea lotus L. for Authentication of Thai Medicinal Herbal Tea. Plants 2020, 9, 921. [Google Scholar] [CrossRef]
- Wu, L.; Wu, M.; Cui, N.; Xiang, L.; Li, Y.; Li, X.; Chen, S. Plant super-barcode: A case study on genome-based identification for closely related species of Fritillaria. Chinese Med. 2021, 16, 1–11. [Google Scholar] [CrossRef]
- Mosa, K.A.; Gairola, S.; Jamdade, R.; El-Keblawy, A.; Al Shaer, K.I.; Al Harthi, E.K.; Shabana, H.A.; Mahmoud, T. The Promise of Molecular and Genomic Techniques for Biodiversity Research and DNA Barcoding of the Arabian Peninsula Flora. Front. Plant. Sci. 2019, 9, 1929. [Google Scholar] [CrossRef]
- Jamdade, R.; Upadhyay, M.; Al Shaer, K.; Al Harthi, E.; Al Sallani, M.; Al Jasmi, M.; Ketbi, A. Al Evaluation of Arabian Vascular Plant Barcodes (rbcL and matK): Precision of Unsupervised and Supervised Learning Methods towards Accurate Identification. Plants 2021, 10, 2741. [Google Scholar] [CrossRef]
- Jamdade, R.A.; Mahmoud, T.; Gairola, S. Prospects of genomic resources available at the global databases for the flora of United Arab Emirates. 3 Biotech. 2019, 9, 1–9. [Google Scholar] [CrossRef]
- Urumarudappa, S.K.J.; Tungphatthong, C.; Prombutara, P.; Sukrong, S. DNA metabarcoding to unravel plant species composition in selected herbal medicines on the National List of Essential Medicines (NLEM) of Thailand. Sci. Rep. 2020, 10, 18259. [Google Scholar] [CrossRef]
- Kress, W.J. Plant DNA barcodes: Applications today and in the future. J. Syst. Evol. 2017, 55, 291–307. [Google Scholar] [CrossRef] [Green Version]
- Aghayeva, P.; Cozzolino, S.; Cafasso, D.; Ali-zade, V.; Fineschi, S.; Aghayeva, D. DNA barcoding of native Caucasus herbal plants: Potentials and limitations in complex groups and implications for phylogeographic patterns. Biodivers. Data J. 2021, 9, 1–28. [Google Scholar] [CrossRef]
- Hebert, P.D.N.; Penton, E.H.; Burns, J.M.; Janzen, D.H.; Hallwachs, W. Ten species in one: DNA barcoding reveals cryptic species in the neotropical skipper butterfly Astraptes fulgerator. Proc. Natl. Acad. Sci. USA 2004, 101, 14812–14817. [Google Scholar] [CrossRef] [Green Version]
- Burns, J.M.; Janzen, D.H.; Hajibabaei, M.; Hallwachs, W.; Hebert, P.D.N. DNA barcodes and cryptic species of skipper butterflies in the genus Perichares in Area de Conservación Guanacaste, Costa Rica. Proc. Natl. Acad. Sci. USA 2008, 105, 6350–6355. [Google Scholar] [CrossRef] [Green Version]
- Dick, C.W.; Webb, C.O. Plant DNA Barcodes, Taxonomic Management, and Species Discovery in Tropical Forests. Methods Mol. Biol. 2012, 858, 379–393. [Google Scholar] [CrossRef] [Green Version]
- Celiński, K.; Kijak, H.; Wojnicka-Półtorak, A.; Buczkowska-Chmielewska, K.; Sokołowska, J.; Chudzińska, E. Effectiveness of the DNA barcoding approach for closely related conifers discrimination: A case study of the Pinus mugo complex. Comptes Rendus Biol. 2017, 340, 339–348. [Google Scholar] [CrossRef]
- Yu, J.; Wu, X.; Liu, C.; Newmaster, S.; Ragupathy, S.; Kress, W.J. Progress in the use of DNA barcodes in the identification and classification of medicinal plants. Ecotoxicol. Environ. Saf. 2021, 208, 111691. [Google Scholar] [CrossRef]
- Techen, N.; Parveen, I.; Pan, Z.; Khan, I.A. DNA barcoding of medicinal plant material for identification. Curr. Opin. Biotechnol. 2014, 25, 103–110. [Google Scholar] [CrossRef]
- Nazar, N.; Howard, C.; Slater, A.; Sgamma, T. Challenges in Medicinal and Aromatic Plants DNA Barcoding—Lessons from the Lamiaceae. Plants 2022, 11, 137. [Google Scholar] [CrossRef]
- CBOL Plant Working Group; Hollingsworth, P.M.; Forrest, L.L.; Spouge, J.L.; Hajibabaei, M.; Ratnasingham, S.; van der Bank, M.; Chase, M.W.; Cowan, R.S.; Erickson, D.L.; et al. A DNA barcode for land plants. Proc. Natl. Acad. Sci. USA 2009, 106, 12794–12797. [Google Scholar] [CrossRef] [Green Version]
- Kress, W.J.; Wurdack, K.J.; Zimmer, E.A.; Weigt, L.A.; Janzen, D.H. Use of DNA barcodes to identify flowering plants. Proc. Natl. Acad. Sci. USA 2005, 102, 8369–8374. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kress, W.J.; Erickson, D.L. A Two-Locus Global DNA Barcode for Land Plants: The Coding rbcL Gene Complements the Non-Coding trnH-psbA Spacer Region. PLoS ONE 2007, 2, e508. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chase, M.W.; Salamin, N.; Wilkinson, M.; Dunwell, J.M.; Kesanakurthi, R.P.; Haidar, N.; Savolainen, V. Land plants and DNA barcodes: Short-term and long-term goals. Philos. Trans. R. Soc. B Biol. Sci. 2005, 360, 1889–1895. [Google Scholar] [CrossRef] [Green Version]
- Taberlet, P.; Coissac, E.; Pompanon, F.; Gielly, L.; Miquel, C.; Valentini, A.; Vermat, T.; Corthier, G.; Brochmann, C.; Willerslev, E. Power and limitations of the chloroplast trnL (UAA) intron for plant DNA barcoding. Nucleic Acids Res. 2007, 35. [Google Scholar] [CrossRef] [Green Version]
- Mishra, P.; Kumar, A.; Nagireddy, A.; Mani, D.N.; Shukla, A.K.; Tiwari, R.; Sundaresan, V. DNA barcoding: An efficient tool to overcome authentication challenges in the herbal market. Plant. Biotechnol. J. 2016, 14, 8–21. [Google Scholar] [CrossRef]
- Hashim, A.M.; Alatawi, A.; Altaf, F.M.; Qari, S.H.; Elhady, M.E.; Osman, G.H.; Abouseadaa, H.H. Phylogenetic relationships and DNA barcoding of nine endangered medicinal plant species endemic to Saint Katherine protectorate. Saudi J. Biol. Sci. 2021, 28, 1919–1930. [Google Scholar] [CrossRef]
- Gao, T.; Yao, H.; Song, J.; Liu, C.; Zhu, Y.; Ma, X.; Pang, X.; Xu, H.; Chen, S. Identification of medicinal plants in the family Fabaceae using a potential DNA barcode ITS2. J. Ethnopharmacol. 2010, 130, 116–121. [Google Scholar] [CrossRef]
- Jiao, J.; Huang, W.; Bai, Z.; Liu, F.; Ma, C.; Liang, Z. DNA barcoding for the efficient and accurate identification of medicinal polygonati rhizoma in China. PLoS ONE 2018, 13, e0201015. [Google Scholar] [CrossRef]
- Pathak, M.R.; Mohamed, A.A.M.; Farooq, M. DNA Barcoding and Identification of Medicinal Plants in the Kingdom of Bahrain. Am. J. Plant. Sci. 2018, 9, 2757–2774. [Google Scholar] [CrossRef]
- Tahir, A.; Hussain, F.; Ahmed, N.; Ghorbani, A.; Jamil, A. Assessing universality of DNA barcoding in geographically isolated selected desert medicinal species of Fabaceae and Poaceae. Peer J. 2018, 6, e4499. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; Chen, M.; Dong, X.; Lin, R.; Fan, J.; Chen, Z. Evaluation of four commonly used DNA barcoding loci for Chinese medicinal plants of the family Schisandraceae. PLoS ONE 2015, 10, e0125574. [Google Scholar] [CrossRef]
- Moon, B.C.; Kim, W.J.; Ji, Y.; Lee, Y.M.; Kang, Y.M.; Choi, G. Molecular identification of the traditional herbal medicines, Arisaematis Rhizoma and Pinelliae Tuber, and common adulterants via universal DNA barcode sequences. Genet. Mol. Res. 2016, 15, gmr7064. [Google Scholar] [CrossRef]
- Kim, W.J.; Ji, Y.; Choi, G.; Kang, Y.M.; Yang, S.; Moon, B.C. Molecular identification and phylogenetic analysis of important medicinal plant species in genus Paeonia based on rDNA-ITS, matK, and rbcL DNA barcode sequences. Genet. Mol. Res. 2016, 15, gmr.15038472. [Google Scholar] [CrossRef]
- Theodoridis, S.; Stefanaki, A.; Tezcan, M.; Aki, C.; Kokkini, S.; Vlachonasios, K.E. DNA barcoding in native plants of the Labiatae (Lamiaceae) family from Chios Island (Greece) and the adjacent Çeşme-Karaburun Peninsula (Turkey). Mol. Ecol. Resour. 2012, 12, 620–633. [Google Scholar] [CrossRef]
- Schori, M.; Showalter, A.M. DNA barcoding as a means for identifying medicinal plants of Pakistan. Pakistan J. Bot. 2011, 43, 1–4. [Google Scholar]
- Ducasse, J.; Ung, V.; Lecointre, G.; Miralles, A. LIMES: A tool for comparing species partition. Bioinformatics 2020, 36, 2282–2283. [Google Scholar] [CrossRef]
- Puillandre, N.; Brouillet, S.; Achaz, G. ASAP: Assemble species by automatic partitioning. Mol. Ecol. Resour. 2021, 21, 609–620. [Google Scholar] [CrossRef]
- Puillandre, N.; Lambert, A.; Brouillet, S.; Achaz, G. ABGD, Automatic Barcode Gap Discovery for primary species delimitation. Mol. Ecol. 2012, 21, 1864–1877. [Google Scholar] [CrossRef]
- He, T.; Jiao, L.; Wiedenhoeft, A.C.; Yin, Y. Machine learning approaches outperform distance- and tree-based methods for DNA barcoding of Pterocarpus wood. Planta 2019, 249, 1617–1625. [Google Scholar] [CrossRef]
- Emu, M.; Sakib, S. Species Identification using DNA Barcode Sequences through Supervised Learning Methods. In Proceedings of the 2019 International Conference on Electrical, Computer and Communication Engineering (ECCE), Cox’sBazar, Bangladesh, 7–9 February 2019. [Google Scholar] [CrossRef]
- Weitschek, E.; Fiscon, G.; Felici, G. Supervised DNA Barcodes species classification: Analysis, comparisons and results. BioData Min. 2014, 7, 1–18. [Google Scholar] [CrossRef] [Green Version]
- Patil, T.S.; Jamdade, R.A.; Patil, S.M.; Govindwar, S.P.; Muley, D.V. DNA barcode based delineation of freshwater fishes from northern Western Ghats of India, one of the world’s biodiversity hotspots. Biodivers. Conserv. 2018, 27, 3349–3371. [Google Scholar] [CrossRef]
- Enan, M.R.; Palakkott, A.R.; Ksiksi, T.S. DNA barcoding of selected UAE medicinal plant species: A comparative assessment of herbarium and fresh samples. Physiol. Mol. Biol. Plants 2017, 23, 221–227. [Google Scholar] [CrossRef] [Green Version]
- Maloukh, L.; Kumarappan, A.; Jarrar, M.; Salehi, J.; El-wakil, H.; Rajya Lakshmi, T.V. Discriminatory power of rbcL barcode locus for authentication of some of United Arab Emirates (UAE) native plants. 3 Biotech. 2017, 7, 144. [Google Scholar] [CrossRef] [Green Version]
- Sukhorukov, A.P.; Mavrodiev, E.V.; Struwig, M.; Nilova, M.V.; Dzhalilova, K.K.; Balandin, S.A.; Erst, A.; Krinitsyna, A.A. One-seeded fruits in the core caryophyllales: Their origin and structural diversity. PLoS ONE 2015, 10, e0130783. [Google Scholar] [CrossRef] [Green Version]
- Kool, A.; de Boer, H.J.; Krüger, Å.; Rydberg, A.; Abbad, A.; Björk, L.; Martin, G. Molecular identification of commercialized medicinal plants in Southern Morocco. PLoS ONE 2012, 7, e39459. [Google Scholar] [CrossRef]
- Hernández-Ledesma, P.; Berendsohn, W.G.; Borsch, T.; Von Mering, S.; Akhani, H.; Arias, S.; Castañeda-Noa, I.; Eggli, U.; Eriksson, R.; Flores-Olvera, H.; et al. A taxonomic backbone for the global synthesis of species diversity in the angiosperm order caryophyllales. Willdenowia 2015, 45, 281–383. [Google Scholar] [CrossRef] [Green Version]
- Cuénoud, P.; Savolainen, V.; Chatrou, L.W.; Powell, M.; Grayer, R.J.; Chase, M.W. Molecular phylogenetics of Caryophyllales based on nuclear 18S rDNA and plastid rbcL, atpB, and matK DNA sequences. Am. J. Bot. 2002, 89, 132–144. [Google Scholar] [CrossRef]
- Jongbloed, M.V.D.; Feulner, G.R.; Böer, B.B.; Western, A.R. The Comprehensive Guide to the Wild Flowers of the United Arab Emirates; Environmental Research and Wildlife Development Agency: Abu Dhabi, United Arab Emirates, 2003; ISBN 978-9948408246. [Google Scholar]
- Karim, F.M.; Fawzi, N.M. Flora of the United Arab Emirates; Publications Department; United Arab Emirates University: Al Ain, United Arab Emirates, 2007; ISBN 9789948021407. [Google Scholar]
- Feulner, G.R. The Flora of the Ru’us al-Jibal—the Mountains of the Musandam Peninsula: An Annotated Checklist and Selected Observations. Available online: http://www.enhg.org/Portals/1/trib/V19/TribulusV19.pdf (accessed on 6 July 2021).
- Feulner, G.R. The Olive Highlands: A Unique “Island” of Biodiversity within the Hajar Mountains of the United Arab Emirates. Available online: http://www.enhg.org/Portals/1/trib/V22/TribulusV22.pdf (accessed on 6 July 2021).
- Levin, R.A.; Wagner, W.L.; Hoch, P.C.; Nepokroeff, M.; Pires, J.C.; Zimmer, E.A.; Sytsma, K.J. Family-level relationships of Onagraceae based on chloroplast rbc L and ndh F data. Am. J. Bot. 2003, 90, 107–115. [Google Scholar] [CrossRef] [Green Version]
- Lee, H.L.; Yi, D.K.; Kim, J.S. Development of plant DNA barcoding markers from the variable noncoding regions of chloroplast genome. In Proceedings of the Abstract Presented at the Second International Barcode of Life Conference, Academia Sinica, Taipei, Taiwan, 18–20 September 2007. [Google Scholar]
- Chen, S.; Yao, H.; Han, J.; Liu, C.; Song, J.; Shi, L.; Zhu, Y.; Ma, X.; Gao, T.; Pang, X.; et al. Validation of the ITS2 Region as a Novel DNA Barcode for Identifying Medicinal Plant Species. PLoS ONE 2010, 5, e8613. [Google Scholar] [CrossRef]
- White, T.; Bruns, T.; Lee, S.; Taylor, J. No Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols: A Guide to Methods and Applications; MA, W.T., Gelfand, D., Sninsky, J., Eds.; Academic Press: New York, NY, USA, 1990; pp. 315–322. [Google Scholar]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef] [PubMed]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Hall, M.; Frank, E.; Holmes, G.; Pfahringer, B.; Reutemann, P.; Witten, I.H. The WEKA data mining software. ACM SIGKDD Explor. Newsl. 2009, 11, 10–18. [Google Scholar] [CrossRef]
- Balunas, M.J.; Kinghorn, A.D. Drug discovery from medicinal plants. Life Sci. 2005, 78, 431–441. [Google Scholar] [CrossRef] [PubMed]
- Bafeel, S.O.; Arif, I.A.; Bakir, M.A.; Khan, H.A.; Al Farhan, A.H.; Al Homaidan, A.A.; Ahamed, A.; Thomas, J. Comparative Evaluation of PCR Success with Universal Primers of Maturase K (matK) and Ribulose-1, 5-Bisphosphate Carboxylase Oxygenase Large Subunit (rbcL) for Barcoding of Some Arid Plants. Plant. Omics 2011, 4, 195–198. [Google Scholar]
- Costion, C.; Ford, A.; Cross, H.; Crayn, D.; Harrington, M.; Lowe, A. Plant DNA Barcodes Can Accurately Estimate Species Richness in Poorly Known Floras. PLoS ONE 2011, 6, e26841. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- YU, J.; XUE, J.-H.; ZHOU, S.-L. New universal matK primers for DNA barcoding angiosperms. J. Syst. Evol. 2011, 49, 176–181. [Google Scholar] [CrossRef]
- Fazekas, A.J.; Kuzmina, M.L.; Newmaster, S.G.; Hollingsworth, P.M. DNA barcoding methods for land plants. Methods Mol. Biol. 2012, 858, 223–252. [Google Scholar] [CrossRef]
- Von Cräutlein, M.; Korpelainen, H.; Pietiläinen, M.; Rikkinen, J. DNA barcoding: A tool for improved taxon identification and detection of species diversity. Biodivers. Conserv. 2011, 20, 373–389. [Google Scholar] [CrossRef]
- Casiraghi, M.; Labra, M.; Ferri, E.; Galimberti, A.; de Mattia, F. DNA barcoding: A six-question tour to improve users’ awareness about the method. Brief. Bioinform. 2010, 11, 440–453. [Google Scholar] [CrossRef]
- Kapralov, M.V.; Akhani, H.; Voznesenskaya, E.V.; Edwards, G.; Franceschi, V.; Roalson, E.H. Phylogenetic Relationships in the Salicornioideae / Suaedoideae / Salsoloideae s.l. (Chenopodiaceae) Clade and a Clarification of the Phylogenetic Position of Bienertia and Alexandra Using Multiple DNA Sequence Datasets. Syst. Bot. 2006, 31, 571–585. [Google Scholar] [CrossRef]
- Wyler, S.C.; Naciri, Y. Evolutionary histories determine DNA barcoding success in vascular plants: Seven case studies using intraspecific broad sampling of closely related species. BMC Evol. Biol. 2016, 16, 103. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.F.; Yang, C.H.; Han, H.L.; Ward, R.D.; Zhang, A. Identifying species of moths (Lepidoptera) from Baihua Mountain, Beijing, China, using DNA barcodes. Ecol. Evol. 2014, 4, 2472–2487. [Google Scholar] [CrossRef]
- Yang, Z.; Landry, J.-F.; Hebert, P.D.N. A DNA Barcode Library for North American Pyraustinae (Lepidoptera: Pyraloidea: Crambidae). PLoS ONE 2016, 11, e0161449. [Google Scholar] [CrossRef] [Green Version]
- Zou, S.; Fei, C.; Song, J.; Bao, Y.; He, M.; Wang, C. Combining and Comparing Coalescent, Distance and Character-Based Approaches for Barcoding Microalgaes: A Test with Chlorella-Like Species (Chlorophyta). PLoS ONE 2016, 11, e0153833. [Google Scholar] [CrossRef] [Green Version]
- Jaén-Molina, R.; Marrero-Rodríguez, Á.; Reyes-Betancort, J.A.; Santos-Guerra, A.; Naranjo-Suárez, J.; Caujapé-Castells, J. Molecular taxonomic identification in the absence of a ‘barcoding gap’: A test with the endemic flora of the Canarian oceanic hotspot. Mol. Ecol. Resour. 2015, 15, 42–56. [Google Scholar] [CrossRef]
- Jiang, K.W.; Zhang, R.; Zhang, Z.F.; Pan, B.; Tian, B. DNA barcoding and molecular phylogeny of Dumasia (Fabaceae: Phaseoleae) reveals a cryptic lineage. Plant. Divers. 2020, 42, 376–385. [Google Scholar] [CrossRef]
- Barley, A.J.; Thomson, R.C. Assessing the performance of DNA barcoding using posterior predictive simulations. Mol. Ecol. 2016, 25, 1944–1957. [Google Scholar] [CrossRef]
- Morello, L.; Braglia, L.; Gavazzi, F.; Gianì, S.; Breviario, D. Tubulin-Based DNA Barcode: Principle and Applications to Complex Food Matrices. Genes 2019, 10, 229. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Barcode Marker | Method | Correct (%) | Incorrect (%) | Ambiguous (%) | Singleton (%) | |
|---|---|---|---|---|---|---|
| rbcL (Spm. = 49, Sp. = 20) | ABGD | SPECIMENS | 57.14 | 40.82 | 0.00 | 2.04 |
| SPECIES | 35.00 | 60.00 | 0.00 | 5.00 | ||
| ASAP | SPECIMENS | 67.35 | 18.37 | 6.12 | 8.16 | |
| SPECIES | 45.00 | 30.00 | 5.00 | 20.00 | ||
| SVM | SPECIMENS | 79.59 | 4.08 | 0.00 | 16.33 | |
| SPECIES | 55.00 | 5.00 | 0.00 | 40.00 | ||
| matK (Spm. = 38, Sp. = 15) | ABGD/ASAP | SPECIMENS | 76.32 | 18.42 | 0.00 | 5.26 |
| SPECIES | 60.00 | 26.67 | 0.00 | 13.33 | ||
| SVM | SPECIMENS | 89.47 | 0.00 | 0.00 | 10.53 | |
| SPECIES | 73.33 | 0.00 | 0.00 | 26.67 | ||
| ITS2 (Spm. = 29, Sp. = 12) | ABGD/ASAP/SVM | SPECIMENS | 96.55 | 0.00 | 0.00 | 3.45 |
| SPECIES | 90.91 | 0.00 | 0.00 | 9.09 |
| Barcode Marker | Max Intra-Sp. Dist. | Avg. Intra-Sp. Dist. | Min. Intra-Sp. Dist. | Max. Inter-Sp. Dist. | Avg. Inter-Sp. Dist. | Min. Inter-Sp. Dist. |
|---|---|---|---|---|---|---|
| rbcL | 0.77 | 0.06 | 0 | 1.55 | 0.45 | 0 |
| matK | 0.69 | 0.18 | 0 | 1.21 | 0.74 | 0 |
| ITS2 | 2.45 | 0.06 | 0 | 2.58 | 2.32 | 2.19 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jamdade, R.; Mosa, K.A.; El-Keblawy, A.; Al Shaer, K.; Al Harthi, E.; Al Sallani, M.; Al Jasmi, M.; Gairola, S.; Shabana, H.; Mahmoud, T. DNA Barcodes for Accurate Identification of Selected Medicinal Plants (Caryophyllales): Toward Barcoding Flowering Plants of the United Arab Emirates. Diversity 2022, 14, 262. https://doi.org/10.3390/d14040262
Jamdade R, Mosa KA, El-Keblawy A, Al Shaer K, Al Harthi E, Al Sallani M, Al Jasmi M, Gairola S, Shabana H, Mahmoud T. DNA Barcodes for Accurate Identification of Selected Medicinal Plants (Caryophyllales): Toward Barcoding Flowering Plants of the United Arab Emirates. Diversity. 2022; 14(4):262. https://doi.org/10.3390/d14040262
Chicago/Turabian StyleJamdade, Rahul, Kareem A. Mosa, Ali El-Keblawy, Khawla Al Shaer, Eman Al Harthi, Mariam Al Sallani, Mariam Al Jasmi, Sanjay Gairola, Hatem Shabana, and Tamer Mahmoud. 2022. "DNA Barcodes for Accurate Identification of Selected Medicinal Plants (Caryophyllales): Toward Barcoding Flowering Plants of the United Arab Emirates" Diversity 14, no. 4: 262. https://doi.org/10.3390/d14040262