
Large-Scale In Vitro Multiplication and Phytochemical Analysis of Himantoglossum affine (Boiss.) Schltr.: An Endangered Euro-Mediterranean Terrestrial Orchid



Abstract
:1. Introduction
2. Materials and Methods
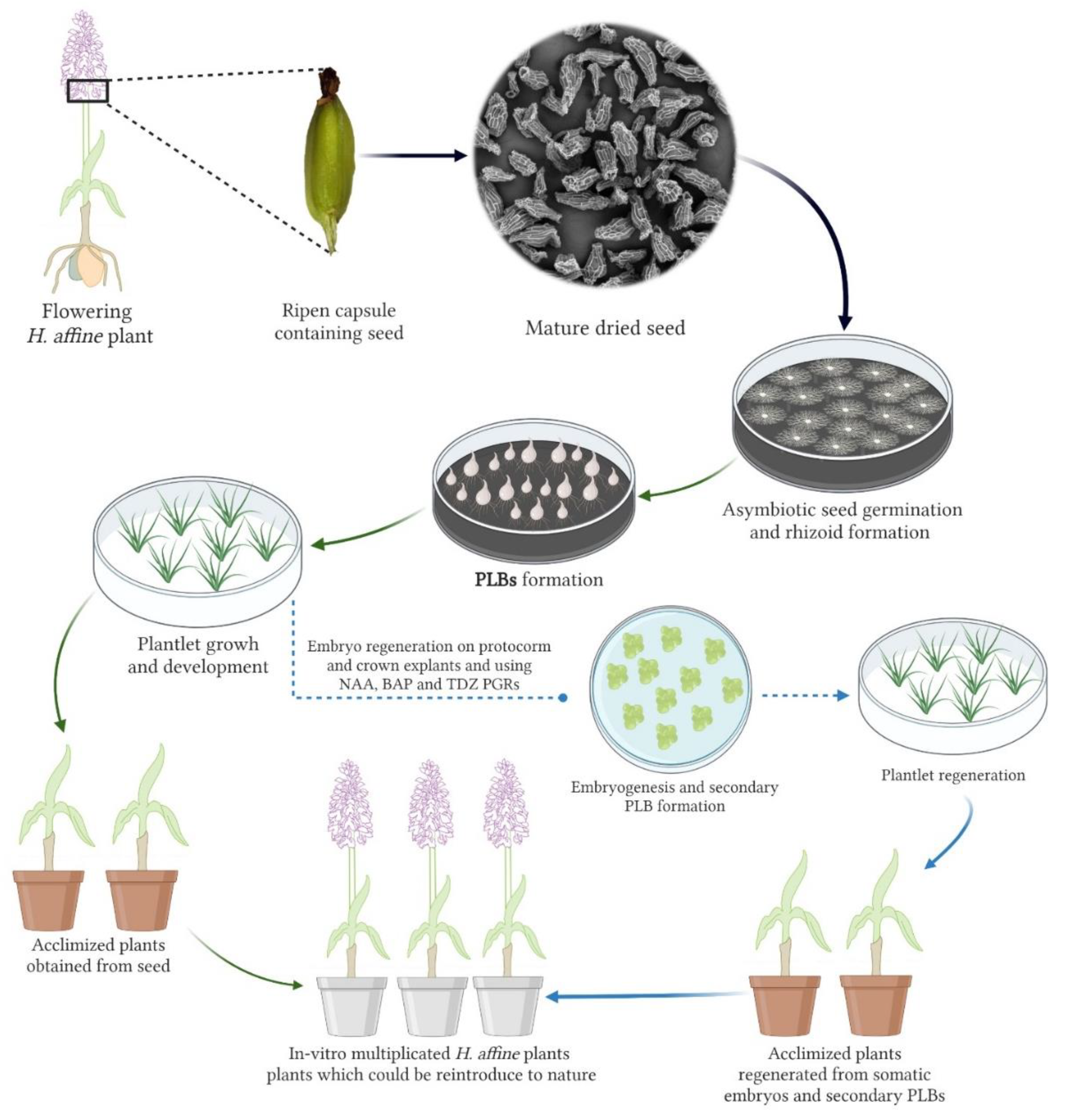
2.1. Plant Material and Explant Preparation for In Vitro Study
2.2. Media Preparation and Treatments
2.3. Extraction and Gas Chromatography–Mass Spectrometry (GC–MS) Analysis
2.4. Determination of Total Phenolic Content (TP), Total Flavonoids Content (TF), and Total Antioxidant Activity (TAA)
2.5. Tuber Bio-Physiochemical Analyses
2.6. SEM and Seed Viability Analyses
2.7. Histochemical Investigation
2.8. Data Analyses
3. Results
3.1. Asymbiotic Seed Germination
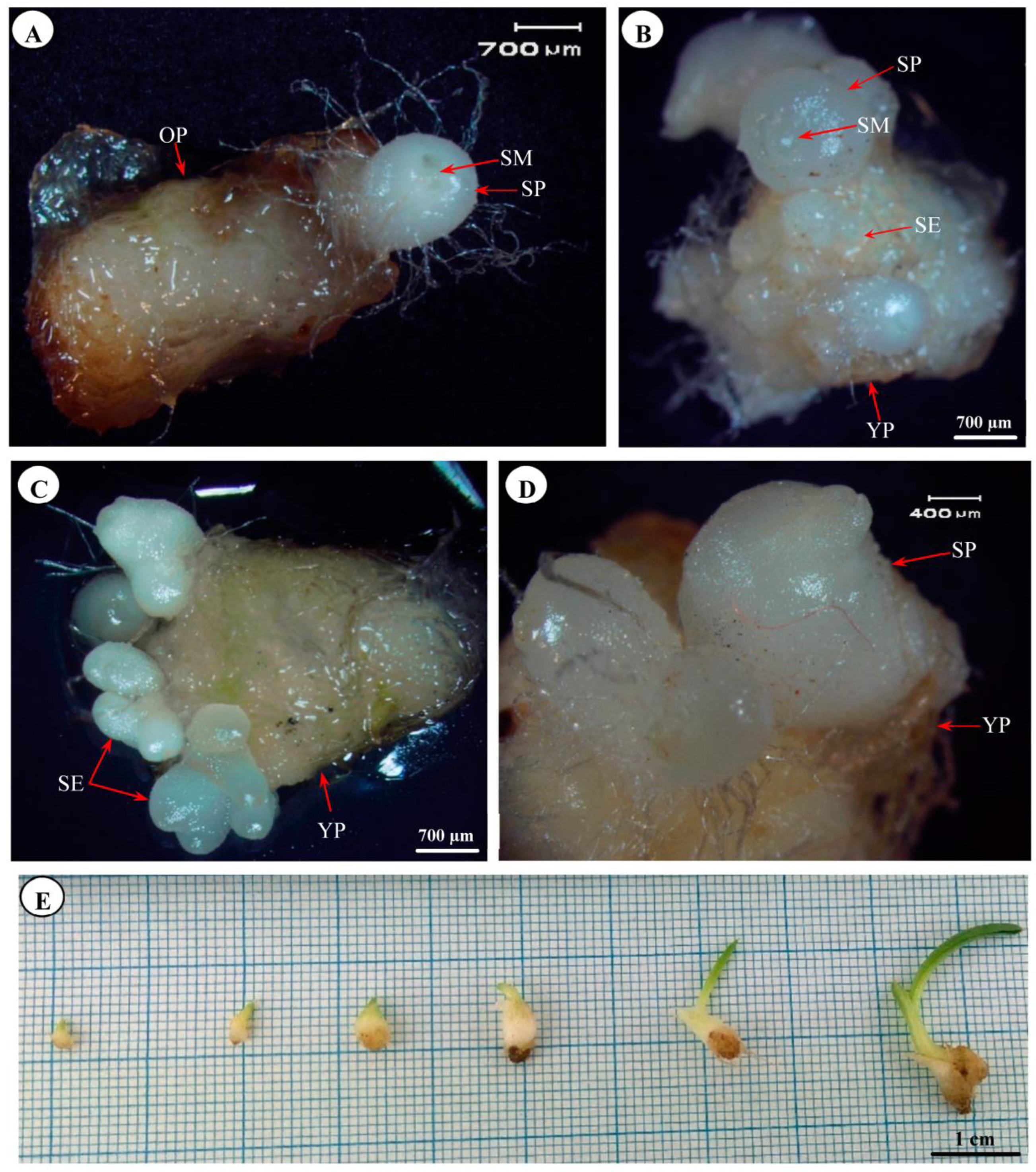
3.2. Somatic Embryogenesis and Secondary Protocorm Formation
3.3. Histology of In Vitro-Raised and Acclimatized Plantlets
3.4. Phytochemical Analysis
3.5. Physico-Biochemical Evaluation
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Shrestha, M.; Dyer, A.; Dorin, A.; Ren, Z.X.; Burd, M. Rewardlessness in orchids: How frequent and how rewardless? Plant Biol. 2020, 22, 555–561. [Google Scholar] [CrossRef] [PubMed]
- Shefferson, R.P.; Jacquemyn, H.; Kull, T.; Hutchings, M.J. The demography of terrestrial orchids: Life history, population dynamics and conservation. Bot. J. Linn. Soc. 2020, 192, 315–332. [Google Scholar] [CrossRef]
- Gaskett, A.C.; Gallagher, R.V. Orchid diversity: Spatial and climatic patterns from herbarium records. Ecol. Evol. 2018, 8, 11235–11245. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de Boer, H.J.; Ghorbani, A.; Manzanilla, V.; Raclariu, A.C.; Kreziou, A.; Ounjai, S.; Osathanunkul, M.; Gravendeel, B. DNA metabarcoding of orchid-derived products reveals widespread illegal orchid trade. Proc. Biol. Sci. 2017, 284, 20171182. [Google Scholar] [CrossRef] [PubMed]
- Renz, J. Flora Iranica. Part 126: Orchidaceae; Naturhistorisches Museum: Vienna, Austria, 1978. [Google Scholar]
- Teoh, E.S. Introduction. In Medicinal Orchids of Asia; Springer International Publishing: Cham, Switzerland, 2016; pp. 3–17. [Google Scholar]
- Farhoosh, R.; Riazi, A. A compositional study on two current types of salep in Iran and their rheological properties as a function of concentration and temperature. Food Hydrocoll. 2007, 21, 660–666. [Google Scholar] [CrossRef]
- Ghorbani, A.; Gravendeel, B.; Zare, S.; De Boer, H.J. Illegal wild collection and international trade of CITES-listed terrestrial orchid tubers in Iran. Traffic Bull. 2014, 26, 52–58. [Google Scholar]
- Ghorbani, A.; Gravendeel, B.; Naghibi, F.; de Boer, H. Wild orchid tuber collection in Iran: A wake-up call for conservation. Biodivers. Conserv. 2014, 23, 2749–2760. [Google Scholar] [CrossRef]
- Ghorbani, A.; Gravendeel, B.; Selliah, S.; Zarre, S.; de Boer, H. DNA barcoding of tuberous Orchidoideae: A resource for identification of orchids used in Salep. Mol. Ecol. Resour. 2017, 17, 342–352. [Google Scholar] [CrossRef] [Green Version]
- Vafaee, Y.; Mohammadi, G.; Nazari, F.; Fatahi, M.; Kaki, A.; Gholami, S.; Ghorbani, A.; Khadivi, A. Phenotypic characterization and seed-micromorphology diversity of the threatened terrestrial orchids: Implications for conservation. S. Afr. J. Bot. 2021, 137, 386–398. [Google Scholar] [CrossRef]
- Gholami, S.; Vafaee, Y.; Nazari, F.; Ghorbani, A. Exploring genetic variations in threatened medicinal orchids using start codon targeted (SCoT) polymorphism and marker-association with seed morphometric traits. Physiol. Mol. Biol. Plants 2021, 27, 769–785. [Google Scholar] [CrossRef]
- Kaki, A.; Vafaee, Y.; Khadivi, A. Genetic variation of Anacamptis coriophora, Dactylorhiza umbrosa, Himantoglossum affine, Orchis mascula, and Ophrys schulzei in the western parts of Iran. Ind. Crops Prod. 2020, 156, 112854. [Google Scholar] [CrossRef]
- Gholami, S.; Vafaee, Y.; Nazari, F.; Ghorbani, A. Molecular characterization of endangered Iranian terrestrial orchids using ISSR markers and association with floral and tuber-related phenotypic traits. Physiol. Mol. Biol. Plants 2021, 27, 53–68. [Google Scholar] [CrossRef] [PubMed]
- Bateman, R.M.; Molnar, V.A.; Sramko, G. In situ morphometric survey elucidates the evolutionary systematics of the Eurasian Himantoglossum clade (Orchidaceae: Orchidinae). PeerJ 2017, 5, e2893. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sramko, G.; Attila, M.V.; Hawkins, J.A.; Bateman, R.M. Molecular phylogeny and evolutionary history of the Eurasiatic orchid genus Himantoglossum s.l. (Orchidaceae). Ann. Bot. 2014, 114, 1609–1626. [Google Scholar] [CrossRef]
- Phillips, R.D.; Reiter, N.; Peakall, R. Orchid conservation: From theory to practice. Ann. Bot. 2020, 126, 345–362. [Google Scholar] [CrossRef]
- Zhao, D.K.; Selosse, M.A.; Wu, L.; Luo, Y.; Shao, S.C.; Ruan, Y.L. Orchid reintroduction based on seed germination-promoting mycorrhizal fungi derived from protocorms or seedlings. Front. Plant Sci. 2021, 12, 701152. [Google Scholar] [CrossRef] [PubMed]
- Barthlott, W.; Große-Veldmann, B.; Korotkova, N. Orchid seed diversity: A scanning electron microscopy survey. Englera 2014, 32, 1–245. [Google Scholar]
- Batty, A.L.; Dixon, K.W.; Brundrett, M.; Sivasithamparam, K. Constraints to symbiotic germination of terrestrial orchid seed in a mediterranean bushland. New Phytol. 2001, 152, 511–520. [Google Scholar] [CrossRef]
- Moradi, S.; Dianati Daylami, S.; Arab, M.; Vahdati, K. Direct somatic embryogenesis in Epipactis veratrifolia, a temperate terrestrial orchid. J. Hortic. Sci. Biotechnol. 2016, 92, 88–97. [Google Scholar] [CrossRef]
- Fatahi, M.; Vafaee, Y.; Nazari, F.; Tahir, N.A.-r. In vitro asymbiotic seed germination, protocorm formation, and plantlet development of Orchis simia Lam.: A threatened terrestrial orchid species. S. Afr. J. Bot. 2022, 151, 156–165. [Google Scholar] [CrossRef]
- Rasmussen, H.N.; Dixon, K.W.; Jersakova, J.; Tesitelova, T. Germination and seedling establishment in orchids: A complex of requirements. Ann. Bot. 2015, 116, 391–402. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Malmgren, S. Orchid propagation: Theory and practice. In Proceedings of the North American Native Orchids: Propagation and Production, Germantown, MD, USA, 16–17 March 1996; pp. 63–71. [Google Scholar]
- Gutiérrez, R.M.P. Orchids: A review of uses in traditional medicine, its phytochemistry and pharmacology. J. Med. Plants. Res. 2010, 4, 592–638. [Google Scholar]
- Wang, Z.; Zhao, M.; Cui, H.; Li, J.; Wang, M. Transcriptomic landscape of medicinal Dendrobium reveals genes associated with the biosynthesis of bioactive components. Front. Plant Sci. 2020, 11, 391. [Google Scholar] [CrossRef] [PubMed]
- Ece Tamer, C.; Karaman, B.; Utku Copur, O. A Traditional Turkish Beverage: Salep. Food Rev. Int. 2006, 22, 43–50. [Google Scholar] [CrossRef]
- Acemi, A.; Cobanoglu, O.; Turker-Kaya, S. FTIR-based comparative analysis of glucomannan contents in some tuberous orchids, and effects of pre-processing on glucomannan measurement. J. Sci. Food Agric. 2019, 99, 3681–3686. [Google Scholar] [CrossRef]
- Murashige, T.; Skoog, F. A revised medium for rapid growth and bio assays with tobacco tissue cultures. Physiol. Plant. 1962, 15, 473–497. [Google Scholar] [CrossRef]
- Mozafari, A.A.; Vafaee, Y.; Shahyad, M. Phytochemical composition and in vitro antioxidant potential of Cynodon dactylon leaf and rhizome extracts as affected by drying methods and temperatures. J. Food Sci. Technol. 2018, 55, 2220–2229. [Google Scholar] [CrossRef]
- Singleton, V.L.; Orthofer, R.; Lamuela-Raventós, R.M. [14] Analysis of total phenols and other oxidation substrates and antioxidants by means of folin-ciocalteu reagent. Methods Enzymol. 1999, 299, 152–178. [Google Scholar]
- Toor, R.K.; Savage, G.P. Antioxidant activity in different fractions of tomatoes. Food Res. Int. 2005, 38, 487–494. [Google Scholar] [CrossRef]
- Brand-Williams, W.; Cuvelier, M.-E.; Berset, C. Use of a free radical method to evaluate antioxidant activity. LWT-Food Sci. Technol. 1995, 28, 25–30. [Google Scholar] [CrossRef]
- AOAC. Official Methods of Analysis of the Association of Official Analytical Chemists, 14th ed.; Association of Official Analytical Chemists: Washington, DC, USA, 2019. [Google Scholar]
- Bradford, M.M. A rapid and sensitive method for the quantification of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef] [PubMed]
- Liu, P.; Zhang, S.; Zhu, G.; Chen, Y.; Ouyang, H.; Han, M. Professional Standard of the People’ Republic of China for Konjac Flour; Ministry of Agriculture of the People’s Republic of China: Beijing, China, 2002. [Google Scholar]
- McCready, R.; Guggolz, J.; Silviera, V.; Owens, H. Determination of starch and amylose in vegetables. Anal. Chem. 1950, 22, 1156–1158. [Google Scholar] [CrossRef]
- Pritchard, H. Determination of orchid seed viability using fluorescein diacetate. Plant Cell Environ. 1985, 8, 727–730. [Google Scholar]
- Van Waes, J.; Debergh, P. Adaptation of the tetrazolium method for testing the seed viability, and scanning electron microscopy study of some Western European orchids. Physiol. Plant. 1986, 66, 435–442. [Google Scholar] [CrossRef]
- Aybeke, M. Morphological and histochemical investigations on Himantoglossum robertianum (Loisel.) P. Delforge (Orchidaceae) seeds. Plant Syst. Evol. 2013, 300, 91–97. [Google Scholar] [CrossRef]
- Aybeke, M. Comparative anatomy of selected rhizomatous and tuberous taxa of subfamilies Orchidoideae and Epidendroideae (Orchidaceae) as an aid to identification. Plant Syst. Evol. 2012, 298, 1643–1658. [Google Scholar] [CrossRef]
- Şenel, G.; Şeker, Ş.S.; Akbulut, M.K.; Akçin, Ö.E. An integrative anatomical, morphological, micromorphological and molecular approach to Turkish epidendroid and orchidoid species (Orchidaceae). Nord. J. Bot. 2018, 36, e01700. [Google Scholar] [CrossRef]
- Salazar Mercado, S.A.; Caleno, J.D.Q.; Suarez, J.P.R. Optimization of the tetrazolium test in three species of orchids of the Andean forest. Aust. J. Crop Sci. 2020, 14, 822–829. [Google Scholar] [CrossRef]
- Utami, E.S.W.; Hariyanto, S.; Manuhara, Y.S.W. In vitro propagation of the endangered medicinal orchid, Dendrobium lasianthera J.J.Sm through mature seed culture. Asian Pac. J. Trop. Biomed. 2017, 7, 406–410. [Google Scholar] [CrossRef]
- Vujanovic, V.; St-Arnaud, M.; Barabé, D.; Thibeault, G. Viability testing of orchid seed and the promotion of colouration and germination. Ann. Bot. 2000, 86, 79–86. [Google Scholar] [CrossRef] [Green Version]
- Yamazaki, J.; Miyoshi, K. In vitro asymbiotic germination of immature seed and formation of protocorm by Cephalanthera falcata (Orchidaceae). Ann. Bot. 2006, 98, 1197–1206. [Google Scholar] [CrossRef] [PubMed]
- Seaton, P.T.; Hosomi, S.T.; Custódio, C.C.; Marks, T.R.; Machado-Neto, N.B.; Pritchard, H.W. Orchid seed and pollen: A toolkit for long-term storage, viability assessment and conservation. In Orchid Propagation—From Laboratories to Greenhouses; Yeung, L.Y., Chee-Tak, E., Eds.; Humana Press: New York, NY, USA, 2018; pp. 71–98. [Google Scholar]
- Gao, Y.; Zhao, Z.; Li, J.; Liu, N.; Jacquemyn, H.; Guo, S.; Xing, X. Do fungal associates of co-occurring orchids promote seed germination of the widespread orchid species Gymnadenia conopsea? Mycorrhiza 2020, 30, 221–228. [Google Scholar] [CrossRef] [PubMed]
- Perotto, S.; Rodda, M.; Benetti, A.; Sillo, F.; Ercole, E.; Rodda, M.; Girlanda, M.; Murat, C.; Balestrini, R. Gene expression in mycorrhizal orchid protocorms suggests a friendly plant-fungus relationship. Planta 2014, 239, 1337–1349. [Google Scholar] [CrossRef] [PubMed]
- Quiroz, K.; Saavedra, J.; Vogel, H.; Verdugo, G.; Caligari, P.D.S.; Garcia-Gonzales, R. In vitro asymbiotic germination for micropropagation of the recalcitrant terrestrial orchid Chloraea crispa (Orchidaceae). Appl. Plant Sci 2017, 5, apps1600142. [Google Scholar] [CrossRef] [PubMed]
- Nabieva, A.Y. Asymbiotic seed germination and in vitro seedling development of Orchis militaris, an endangered orchid in Siberia. J. Genet. Eng. Biotechnol. 2021, 19, 122. [Google Scholar] [CrossRef]
- Swarts, N.D.; Dixon, K.W. Conservation Methods for Terrestrial Orchids; Ross Publishing: Richmond, VA, USA, 2017; p. 240. [Google Scholar]
- Figura, T.; Tylova, E.; Jersakova, J.; Vohnik, M.; Ponert, J. Fungal symbionts may modulate nitrate inhibitory effect on orchid seed germination. Mycorrhiza 2021, 31, 231–241. [Google Scholar] [CrossRef]
- Figura, T.; Weiser, M.; Ponert, J. Orchid seed sensitivity to nitrate reflects habitat preferences and soil nitrate content. Plant Biol. 2020, 22, 21–29. [Google Scholar] [CrossRef]
- Stewart, S.L.; Kane, M.E. Asymbiotic seed germination and in vitro seedling development of Habenaria macroceratitis (Orchidaceae), a rare Florida terrestrial orchid. Plant Cell Tissue Organ Cult. 2006, 86, 147–158. [Google Scholar] [CrossRef]
- Del Vecchio, S.; Pierce, S.; Fantinato, E.; Buffa, G. Increasing the germination percentage of a declining native orchid (Himantoglossum adriaticum) by pollen transfer and outbreeding between populations. Plant Biol. 2019, 21, 935–941. [Google Scholar] [CrossRef]
- Calevo, J.; Copetta, A.; Marchioni, I.; Bazzicalupo, M.; Pianta, M.; Shirmohammadi, N.; Cornara, L.; Giovannini, A. The use of a new culture medium and organic supplement to improve in vitro early stage development of five orchid species. Plant Biosyst. Int. J. Deal. All Asp. Plant Biol. 2022, 156, 143–151. [Google Scholar] [CrossRef]
- Dulić, J.; Ljubojević, M.; Ognjanov, V.; Barać, G.; Dulić, T. In vitro germination and seedling development of two European orchid species, Himantoglossum jankae Somlyay, Kreutz & Óvári and Spiranthes spiralis (L.) Chevall. In Vitro Cell. Dev. Biol. Plant 2019, 55, 380–391. [Google Scholar] [CrossRef]
- Kitsaki, C.K.; Zygouraki, S.; Ziobora, M.; Kintzios, S. In vitro germination, protocorm formation and plantlet development of mature versus immature seeds from several Ophrys species (Orchidaceae). Plant Cell Rep. 2004, 23, 284–290. [Google Scholar] [CrossRef] [PubMed]
- Yong, J.W.; Ge, L.; Ng, Y.F.; Tan, S.N. The chemical composition and biological properties of coconut (Cocos nucifera L.) water. Molecules 2009, 14, 5144–5164. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Utami, E.S.W.; Hariyanto, S. Organic compounds: Contents and their role in improving seed germination and protocorm development in orchids. Int. J. Agron. 2020, 2020, 2795108. [Google Scholar] [CrossRef]
- Kunakhonnuruk, B.; Inthima, P.; Kongbangkerd, A. In vitro propagation of Epipactis flava Seidenf., an endangered rheophytic orchid: A first study on factors affecting asymbiotic seed germination, seedling development and greenhouse acclimatization. Plant Cell Tissue Organ Cult. 2018, 135, 419–432. [Google Scholar] [CrossRef]
- Jørgensen, B.I. Hardy orchids: Symbiotic in vitro propagation and cultivation. Environ. Eff. Control Plant Tissue Cult. 1994, 393, 165–172. [Google Scholar] [CrossRef]
- Manokari, M.; Priyadharshini, S.; Shekhawat, M.S. Direct somatic embryogenesis using leaf explants and short term storage of synseeds in Spathoglottis plicata Blume. Plant Cell Tissue Organ Cult. 2021, 145, 321–331. [Google Scholar] [CrossRef]
- Soonthornkalump, S.; Nakkanong, K.; Meesawat, U. In vitro cloning via direct somatic embryogenesis and genetic stability assessment of Paphiopedilum niveum (Rchb.f.) Stein: The endangered Venus’s slipper orchid. In Vitro Cell. Dev. Biol. Plant 2019, 55, 265–276. [Google Scholar] [CrossRef]
- Shen, H.J.; Chen, J.T.; Chung, H.H.; Chang, W.C. Plant regeneration via direct somatic embryogenesis from leaf explants of Tolumnia Louise Elmore ‘Elsa’. Bot. Stud. 2018, 59, 4. [Google Scholar] [CrossRef] [Green Version]
- Sherif, N.A.; Franklin Benjamin, J.H.; Senthil Kumar, T.; Rao, M.V. Somatic embryogenesis, acclimatization and genetic homogeneity assessment of regenerated plantlets of Anoectochilus elatus Lindl., an endangered terrestrial jewel orchid. Plant Cell Tissue Organ Cult. 2017, 132, 303–316. [Google Scholar] [CrossRef]
- Parthibhan, S.; Rao, M.V.; Teixeira da Silva, J.; Senthil Kumar, T. Somatic embryogenesis from stem thin cell layers of Dendrobium aqueum. Biol. Plant. 2018, 62, 439–450. [Google Scholar] [CrossRef]
- Mahendran, G.; Narmatha Bai, V. Direct somatic embryogenesis of Malaxis densiflora (A. Rich.) Kuntze. J. Genet. Eng. Biotechnol. 2016, 14, 77–81. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chugh, S.; Guha, S.; Rao, I.U. Micropropagation of orchids: A review on the potential of different explants. Sci. Hortic. 2009, 122, 507–520. [Google Scholar] [CrossRef]
- Cardoso, J.C.; Zanello, C.A.; Chen, J.T. An overview of orchid protocorm-like bodies: Mass propagation, biotechnology, molecular aspects, and breeding. Int. J. Mol. Sci. 2020, 21, 985. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Draper, J.T.; Conran, J.G.; Crouch, N.; Weinstein, P.; Simpson, B.S. Sexual dimorphism in the dioecious monocot Lomandra leucocephala ssp. robusta and its potential ecosystem and conservation significance. Aust. J. Bot. 2020, 68, 275–287. [Google Scholar] [CrossRef]
- McCormick, D. Characterisation of Vanilla Extracts Based on Sensory Properties and Chemical Composition: A Thesis Presented in Partial Fulfilment of the Requirements for the Degree of Doctor of Philosophy in Food Technology at Massey University, New Zealand. Ph.D. Thesis, Massey University, Palmerston North, New Zealand, 2018. [Google Scholar]
- Casuga, F.P.; Castillo, A.L.; Corpuz, M.J.-A.T. GC–MS analysis of bioactive compounds present in different extracts of an endemic plant Broussonetia luzonica (Blanco)(Moraceae) leaves. Asian Pac. J. Trop. Biomed. 2016, 6, 957–961. [Google Scholar] [CrossRef] [Green Version]
- Salih, T.F.M.; Mohammed, L.M.; Qader, K.O. Chemical Analysis and Growth Inhibitory Effect of Rosemary Plant on Aspergillus niger. Kurd. J. Appl. Res. 2017, 2, 151–154. [Google Scholar] [CrossRef] [Green Version]
- Chowdhury, J.U.; Bhuiyan, M.N.I.; Begum, J. Constituents of leaf and fruits essential oil of Carum roxburghianum Benth. J. Sci. Res. 2009, 1, 160–163. [Google Scholar] [CrossRef]
- Wang, Q.; Ou, Z.; Lei, H.; Zeng, X.; Ying, Y.; Bai, W. Antimicrobial activities of a new formula of spice water extracts against foodborne bacteria. J. Food Process. Preserv. 2012, 36, 374–381. [Google Scholar] [CrossRef]
- Jungermann, E.; Sonntag, N.O. Glycerine: A Key Cosmetic Ingredient; CRC Press: Boca Raton, FL, USA, 1991; Volume 11. [Google Scholar]
- Olaniyan, O.T.; Kunle-Alabi, O.T.; Raji, Y. Protective effects of methanol extract of Plukenetia conophora seeds and 4H-Pyran-4-One 2,3-Dihydro-3,5-Dihydroxy-6-Methyl on the reproductive function of male Wistar rats treated with cadmium chloride. JBRA Assist. Reprod. 2018, 22, 289–300. [Google Scholar] [CrossRef]
- Ghosh, G.; Panda, P.; Rath, M.; Pal, A.; Sharma, T.; Das, D. GC-MS analysis of bioactive compounds in the methanol extract of Clerodendrum viscosum leaves. Pharmacogn. Res. 2015, 7, 110–113. [Google Scholar] [CrossRef] [Green Version]
- Peng, W.-X.; Wu, S.-B.; Yang, L.; Li, K.-F.; Peng, W.-L.; Hong, S.-Y. Determination of Chemical Components of Benzene/Ethanol Extractives of Castanopsis Fissa Wood by GC/MS. In Proceedings of the 2008 2nd International Conference on Bioinformatics and Biomedical Engineering, Shanghai, China, 16–18 May 2008; pp. 3165–3167. [Google Scholar]
- Sympli, H.D. Estimation of drug-likeness properties of GC-MS separated bioactive compounds in rare medicinal Pleione maculata using molecular docking technique and SwissADME in silico tools. Netw. Model. Anal. Health Inf. Bioinform. 2021, 10, 14. [Google Scholar] [CrossRef] [PubMed]
- Khadartsev, A.; Sukhikh, G.T.; Volochaeva, M.V.; Platonov, V.V.; Dunaeva, I.V. Chromato-mass spectrometry of Orchis maculata ethanol extract (Orchidaceae), e-edition. J. New Med. Technol. 2019, 13, 102–121. [Google Scholar]
- Shah, S.; Chand, K.; Rekadwad, B.; Shouche, Y.S.; Sharma, J.; Pant, B. A prospectus of plant growth promoting endophytic bacterium from orchid (Vanda cristata). BMC Biotechnol. 2021, 21, 16. [Google Scholar] [CrossRef] [PubMed]
- Bazzicalupo, M.; Burlando, B.; Denaro, M.; Barreca, D.; Trombetta, D.; Smeriglio, A.; Cornara, L. Polyphenol Characterization and Skin-Preserving Properties of Hydroalcoholic Flower Extract from Himantoglossum robertianum (Orchidaceae). Plants 2019, 8, 502. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hurkan, K.; Yuksel, M.B.; Hurkan, Y.K.; Demir, N. Determination of total phenolic and flavonoid contents, antioxidant and antimicrobial activities of some important salep orchids. Eskişehir Teknik Üniversitesi Bilim ve Teknoloji Dergisi-C Yaşam Bilimleri ve Biyoteknoloji 2019, 8, 191–202. [Google Scholar] [CrossRef]
- Dalar, A.; Guo, Y.; Esim, N.; Bengu, A.S.; Konczak, I. Health attributes of an endemic orchid from Eastern Anatolia, Dactylorhiza chuhensis Renz&Taub.—In vitro investigations. J. Herb. Med. 2015, 5, 77–85. [Google Scholar] [CrossRef] [Green Version]
- Bhattacharyya, P.; Kumaria, S.; Job, N.; Tandon, P. En-masse production of elite clones of Dendrobium crepidatum: A threatened, medicinal orchid used in Traditional Chinese Medicine (TCM). J. Appl. Res. Med. Aromat. Plants 2016, 3, 168–176. [Google Scholar] [CrossRef]
- Şen, M.A.; Palabiyik, I.; Kurultay, Ş. Composition, viscosity and solubility of saleps from twenty different orchid (Orchidaceae) species. J. Food Meas. Charact. 2018, 12, 1334–1339. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Trait | PJ | CW | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| AV | Pep | CH | AV + Pep | AV + CH | Pep + CH | AV | Pep | CH | AV + Pep | AV + CH | Pep + CH | |
| Germination percent (%) | 99.46 ± 0.38 a | 98.77 ± 0.37 a | 99.00 ± 0.47 a | 95.38 ± 1.51 b | 64.85 ± 2.01 e | 75.99 ± 1.43 cd | 61.28 ± 1.17 f | 58.2 ± 0.76 g | 50.06 ± 1.44 h | 77.64 ± 0.73 c | 75.46 ± 0.53 cd | 66.62 ± 0.49 e |
| Time to germination (day) | 51.20 ± 1.46 i | 56.00 ± 1.64 h | 34.20 ± 0.20 g | 32.80 ± 0.91 f | 31.00 ± 0.77 f | 27.60 ± 0.40 e | 19.60 ± 0.50 c | 22.60 ± 0.50 d | 16.60 ± 0.40 a | 18.00 ± 0.00 b | 17.00 ± 0.31 a | 38.00 ± 0.83 i |
| Protocorm length (cm) | 0.56 ± 0.04 b | 0.30 ± 0.02 b | 0.26 ± 0.02 c | 0.58 ± 0.06 b | 0.25 ± 0.03 d | 0.38 ± 0.03 d | 0.64 ± 0.04 a | 0.64 ± 0.05 a | 0.48 ± 0.06 c | 0.32 ± 0.03 e | 0.34 ± 0.02 e | 0.42 ± 0.04 d |
| Protocorm width (cm) | 0.28 ± 0.040 b | 0.20 ± 0.05 cd | 0.20 ± 0.03 c | 0.26 ± 0.05 b | 0.17 ± 0.02 d | 0.20 ± 0.00 c | 0.32 ± 0.05 a | 0.27 ± 0.03 b | 0.24 ± 0.02 c | 0.20 ± 0.04 cd | 0.19 ± 0.02 d | 0.26 ± 0.08 bc |
| Protocorm weight (g) | 0.07 ± 0.00 b | 0.04 ± 0.00 d | 0.03 ± 0.00 ef | 0.08 ± 0.00 ab | 0.03 ± 0.00 ef | 0.04 ± 0.00 de | 0.09 ± 0.01 a | 0.08 ± 0.00 ab | 0.66 ± 0.00 c | 0.03 ± 0.00 e | 0.04 ± 0.00 de | 0.06 ± 0.00 c |
| Plantlet length (cm) | 3.58 ± 0.03 d | 4.20 ± 0.04 a | 2.58 ± 0.03 d | 3.10 ± 0.05 b | 3.06 ± 0.09 b | 2.44 ± 0.01 e | 2.52 ± 0.03 d | 2.54 ± 0.02 d | 2.72 ± 0.03 c | 2.50 ± 0.04 d | 2.16 ± 0.02 g | 2.24 ± 0.02 f |
| Plantlet weight (g) | 0.38 ± 0.00 f | 0.58 ± 0.00 a | 0.25 ± 0.00 e | 0.34 ± 0.00 cd | 0.30 ± 0.003 c | 0.22 ± 0.00 g | 0.26 ± 0.00 ef | 0.27 ± 0.00 d | 0.26 ± 0.00 b | 0.22 ± 0.00 g | 0.02 ± 0.00 i | 0.23 ± 0.00 h |
| Shoot length (cm) | 1.76 ± 0.02 d | 1.96 ± 0.04 a | 1.22 ± 0.03 e | 1.86 ± 0.03 b | 1.62 ± 0.02 b | 1.09 ± 0.02 e | 1.20 ± 0.03 c | 1.36 ± 0.02 f | 1.64 ± 0.02 f | 1.33 ± 0.01 cd | 0.87 ± 0.01 g | 1.04 ± 0.05 g |
| Shoot diameter (cm) | 0.19 ± 0.10 b | 0.20 ± 0.00 b | 0.19 ± 0.00 b | 0.19 ± 0.00 b | 0.09 ± 0.00 e | 0.10 ± 0.00 e | 0.23 ± 0.02 a | 0.20 ± 0.00 b | 0.20 ± 0.01 b | 0.10 ± 0.00 e | 0.12 ± 0.00 d | 0.15 ± 0.00 c |
| Root number (cm) | 1.60 ± 0.024 d | 1.60 ± 0.024 d | 1.00 ± 0.01 g | 1.40 ± 0.02 e | 1.20 ± 0.02 f | 1.20 ± 0.02 f | 5.20 ± 0.20 a | 3.20 ± 0.02 b | 3.00 ± 0.03 c | 0.40 ± 0.02 i | 0.80 ± 0.03 h | 0.40 ± 0.02 i |
| PGRs (mg/L) | Regeneration (%) | The Number of Embryos per Explants | |||||
|---|---|---|---|---|---|---|---|
| Crown | Young PLB | Old PLB | Crown | Young PLB | Old PLB | ||
| NAA | TDZ | ||||||
| 0 | 0 | 0.00 ± 0.00 e | 0.00 ± 0.00 e | 0.00 ± 0.00 e | 0.00 ± 0.00 e | 0.00 ± 0.00 e | 0.00 ± 0.00 e |
| 0 | 0.5 | 0.00 ± 0.00 e | 0.00 ± 0.00 e | 0.00 ± 0.00 e | 0.00 ± 0.00 e | 0.00 ± 0.00 e | 0.00 ± 0.00 e |
| 0 | 1 | 0.00 ± 0.00 e | 4.00 ± 8.00 d | 4.00 ± 8.00 d | 0.00 ± 0.00 e | 5.00 ± 0.00 c | 0.00 ± 0.00 e |
| 0 | 2 | 0.00 ± 0.00 e | 4.00 ± 8.00 d | 8.00 ± 9.79 cd | 0.00 ± 0.00 e | 3.50 ± 0.50 ef | 3.25 ± 0.47 f |
| 0.5 | 0 | 0.00 ± 0.00 e | 0.00 ± 0.00 e | 0.00 ± 0.00 e | 0.00 ± 0.00 e | 0.00 ± 0.00 e | 0.00 ± 0.00 e |
| 0.5 | 0.5 | 8.00 ± 9.79 cd | 12.00 ± 9.79 cd | 8.00 ± 9.79 cd | 4.50 ± 0.50 de | 8.50 ± 0.50 a | 3.50 ± 0.50 ef |
| 0.5 | 1 | 32.00 ± 8.00 b | 44.00 ± 0.00 a | 32.00 ± 8.00 b | 8.33 ± 1.20 ab | 10.12 ± 2.08 a | 4.87 ± 1.05 cf |
| 0.5 | 2 | 12.00 ± 9.79 cd | 20.00 ± 8.00 bc | 16.00 ± 0.00 c | 4.66 ± 1.24 cd | 7.20 ± 0.74 b | 5.75 ± 0.43 c |
| 1 | 0 | 8.00 ± 9.79 cd | 4.00 ± 8.00 d | 0.00 ± 0.00 e | 3.85 ± 0.50 ef | 4.00 ± 0.00 e | 0.00 ± 0.00 e |
| 1 | 0.5 | 12.00 ± 9.79 cd | 24.00 ± 9.79 b | 4.00 ± 8.00 d | 6.33 ± 0.47 bc | 6.20 ± 0.89 bc | 4.00 ± 0.00 e |
| 1 | 1 | 12.00 ± 9.79 cd | 28.00 ± 9.79 b | 24.00 ± 9.79 b | 5.60 ± 0.94 b–d | 8.42 ± 0.72 ab | 8.33 ± 0.94 ab |
| 1 | 2 | 0.00 ± 0.00 e | 12.00 ± 9.79 cd | 20.00 ± 8.00 bc | 0.00 ± 0.00 e | 4.33 ± 0.47 de | 2.50 ± 0.50 h |
| NAA | BAP | ||||||
| 0 | 0.5 | 0.00 ± 0.00 e | 0.00 ± 0.00 e | 0.00 ± 0.00 e | 0.00 ± 0.00 e | 0.00 ± 0.00 e | 0.00 ± 0.00 e |
| 0 | 1 | 0.00 ± 0.00 e | 0.00 ± 0.00 e | 0.00 ± 0.00 e | 0.00 ± 0.00 e | 0.00 ± 0.00 e | 0.00 ± 0.00 e |
| 0 | 2 | 4.00 ± 8.00 d | 4.00 ± 8.00 d | 8.00 ± 9.79 cd | 2.00 ± 0.00 i | 2.00 ± 0.00 i | 2.00 ± 0.00 i |
| 0.5 | 0.5 | 12.00 ± 9.79 cd | 8.00 ± 9.79 cd | 4.00 ± 8.00 d | 3.00 ± 0.00 h | 4.66 ± 1.24 c-f | 2.50 ± 0.50 f-i |
| 0.5 | 1 | 8.00 ± 9.79 cd | 24.00 ± 0.00 b | 4.00 ± 8.00 d | 2.50 ± 0.50 h | 6.50 ± 0.50 bc | 4.00 ± 0.00 e |
| 0.5 | 2 | 12.00 ± 9.79 cd | 16.00 ± 0.00 c | 8.00 ± 9.79 cd | 2.33 ± 0.47 hi | 8.00 ± 0.00 ab | 4.25 ± 0.43 de |
| 1 | 0.5 | 24.00 ± 8.00 b | 8.00 ± 9.79 cd | 4.00 ± 8.00 c | 4.6 ± 1.01 cd | 4.80 ± 0.74 cd | 3.00 ± 0.00 h |
| 1 | 1 | 8.00 ± 9.79 cd | 12.00 ± 9.79 cd | 16.00 ± 8.00 cd | 2.00 ± 1.00 i | 3.25 ± 0.43 f | 1.75 ± 0.43 ij |
| 1 | 2 | 4.00 ± 8.00 d | 12.00 ± 9.79 cd | 20.00 ± 8.00 bc | 2.00 ± 0.00 i | 2.00 ± 0.00 i | 1.60 ± 0.48 ij |
| Methanolic Extract | |||||||||
|---|---|---|---|---|---|---|---|---|---|
| Tuber | Leaf | ||||||||
| Compound | Formula | RT | % | MW | Compound | Formula | RT | % | MW |
| 1,2,3-propanetriol, monoacetate | C10H12O | 17.420 | 47.63 ± 0.82 | 134 | 1,2,3-propanetriol, monoacetate | C10H12O | 17.128 | 42.65 ± 1.84 | 134 |
| Benzene, 1-methoxy-4-(1-propenyl)- | C5H10O4 | 17.964 | 26.75 ± 1.23 | 148 | 4H-Pyran-4-one, 2,3-dihydro-3,5-dihydroxy-6-methyl | C6H8O4 | 14.675 | 11.32 ± 1.86 | 136 |
| Glycerin | C3H8O3 | 28.819 | 16.42 ± 0.66 | 92 | 6-Acetyl-.beta.-d-mannose | C8H14O7 | 20.116 | 10.45 ± 1.80 | 222 |
| (2-Methyl-1-cyclohexenyl) formaldehyde semicarbazone | C9H15N3O | 20.166 | 4.55 ± 0.34 | 137 | alpha-D-glucopyranosyl-(1->3) | C18H32O16 | 20.914 | 10.16 ± 1.02 | 504 |
| Bicyclo[2.2.1]heptan-2-one, 1,3,3-trimethyl- | C10H16O | 12.234 | 1.10 ± 0.50 | 152 | Phenol, 4-(ethoxymethyl)- | C9H12O2 | 20.265 | 6.96 ± 0.91 | 152 |
| 2,5-Methylene-d,l-rhamnitol | C7H14O5 | 14.351 | 0.73 ± 0.28 | 178 | Ribitol | C5H12O5 | 15.720 | 2.59 ± 0.32 | 152 |
| Ethyl(dimethyl)allyloxysilane | C7H16OSi | 9.512 | 0.71 ± 0.12 | 144 | 2-Propanone, 1,1-diethoxy- | C7H14O3 | 15.399 | 2.43 ± 0.49 | 146 |
| Sorbitol | C6H14O6 | 20.958 | 0.46 ± 0.18 | 182 | Hexadecanoic acid, 3-hydroxy-, methyl ester | C17H34O3 | 15.809 | 2.34 ± 0.27 | 286 |
| Trimethylsilylmethanol | C4H12OSi | 9.765 | 0.43 ± 0.20 | 104 | Hexadecanoic acid, methyl ester | C17H34O2 | 38.249 | 1.89 ± 0.36 | 270 |
| Phenol, 3-methyl- | C7H8O16 | 12.832 | 0.33 ± 0.11 | 108 | Trimethylsilylmethanol | C4H12OSi | 10.199 | 1.41 ± 0.27 | 104 |
| Isosorbide Dinitrate | C6H8N2O8 | 11.626 | 0.31 ± 0.16 | 236 | L-Lyxose | C5H10O5 | 9.798 | 1.37 ± 0.44 | 150 |
| Propionic acid, 2-mercapto-, allyl ester | C6H10O2S | 10.176 | 0.26 ± 0.92 | 146 | D-Glucose, 6-O-.alpha.-D-galactopyranosyl- | C12H22O11 | 9.351 | 1.09 ± 0.90 | 342 |
| d-Mannitol, 1-decylsulfonyl- | Undecanoic acid, 3-hydroxy-, methyl ester | C12H24O3 | 18.928 | 0.95 ± 0.28 | 216 | ||||
| N-Methyl-N-(toluene-4-sulfonylmethyl)-acetamide | C11H15NO3S | 13.994 | 0.81 ± 0.55 | 241 | |||||
| beta.-D-Glucopyranose, 4-O-.beta.-D-galactopyranosyl- | C12H22O11 | 24.626 | 0.56 ± 0.21 | 170 | |||||
| 1,2,3,4,5-Cyclopentanepentol | C5H10O5 | 8.255 | 0.54 ± 0.16 | 155 | |||||
| Decanoic acid, 3-hydroxy-, methyl ester | C11H22O3 | 19.110 | 0.45 ± 0.17 | 202 | |||||
| Ethanolic Extract | |||||||||
| Tuber | Leaf | ||||||||
| Compound | Compound | Formula | RT | % | MW | ||||
| 4H-Pyran-4-one, 2,3-dihydro-3,5-dihydroxy-6-methyl | C6H8O4 | 18.028 | 37.53 ± 1.12 | 144 | 2-Butenedioic acid, 2-methyl-, (E)- | C5H6O4 | 12.818 | 19.40 ± 2.95 | 130 |
| alpha.-Pinene | C10H16 | 9.672 | 14.30 ± 1.08 | 136 | Phenol, 4-(ethoxymethyl)- | C9H12O2 | 19.815 | 19.01 ± 1.55 | 152 |
| 2,4-Octadienoic acid, 7-hydroxy-6-methyl- | C9H14O3 | 22.731 | 13.93 ± 1.10 | 170 | 1,3-Diazacyclooctane-2-thione | C6H12N2S | 13.073 | 13.08 ± 2.11 | 144 |
| 2-Furancarboxaldehyde, 5-(hydroxymethyl)- | C6H6O3 | 22.134 | 11.70 ± 0.88 | 121 | Nonanediamide, N,N′-di-benzoyloxy- | C23H26N2O6 | 14.634 | 6.60 ± 0.49 | 426 |
| Phenol, 4-(ethoxymethyl)- | C9H12O2 | 24.558 | 8.26 ± 0.92 | 152 | 5-Methyl-1-phenylhex-5-en-1-one | C13H16O | 15.233 | 6.48 ± 1.41 | 188 |
| 2,4-Dihydroxy-2,5-dimethyl-3(2H)-furan-3-one | C6H8O4 | 11.652 | 3.06 ± 0.28 | 144 | 1,3-Dioxolane | C3H6O2 | 8.790 | 6.09 ± 1.50 | 74 |
| 2-n-Propylthiane | C8H16S | 18.498 | 1.50 ± 0.41 | 144 | 1,3-Dioxolane, 2-(dichloromethyl)- | C4H6Cl2O2 | 8.395 | 5.37 ± 1.17 | 156 |
| Isopinocarveol | C10H16O | 16.817 | 1.20 ± 0.55 | 152 | 6-Acetyl-.beta.-d-mannose | C8H14O7 | 17.292 | 4.72 ± 0.67 | 222 |
| d-Gala-l-ido-octonic amide | C8H17NO8 | 13.044 | 0.92 ± 0.37 | 255 | Benzenemethanol, 2-methoxy-.alpha.-2-propenyl- | C11H14O2 | 17.954 | 2.91 ± 0.89 | 178 |
| D-Glucose, 6-O-.alpha.-D-galactopyranosyl | C12H22O11 | 11.871 | 0.91 ± 0.47 | 342 | 1,2,3,4-Cyclohexanetetrol | C6H12O4 | 9.407 | 2.53 ± 1.01 | 148 |
| 2,2-Dimethyl-3-vinyl-bicyclo[2.2.1]heptane | C11H18 | 15.746 | 0.86 ± 0.10 | 150 | 2H-Indeno[1,2-b]furan-2-one, 3,3a,4,5,6,7,8,8b-octahydro-8,8-dimethyl | C13H18O2 | 22.569 | 2.52 ± 0.52 | 206 |
| Bicyclo[3.1.1]hept-3-en-2-ol, 4,6,6-trimethyl- | C10H16O | 17.104 | 0.83 ± 0.16 | 152 | Bicyclo[3.2.1]oct-6-ene-6,8-dimethanol, 1,7-dimethyl-4-isopropyl-, bis(3,5-dinitrobenzoate) | C29H30N4O12 | 15.606 | 1.99 ± 0.38 | 626 |
| N-Methyl-N-(toluene-4-sulfonylmethyl)-acetamide | C11H15NO3S | 13.994 | 0.81 ± 0.16 | 241 | Geranyl isovalerate | C15H26O2 | 22.862 | 1.97 ± 0.44 | 238 |
| l-Gala-l-ido-octonic lactone | C8H14O8 | 13.158 | 0.70 ± 0.09 | 238 | Dithiocarbamate, S-methyl-,N-(2-methyl-3-oxobutyl)- | C7H13NOS2 | 16.040 | 1.66 ± 0.32 | 191 |
| Ocimene | C10H16 | 12.502 | 0.53 ± 0.11 | 136 | D-Melezitose | C18H32O16 | 11.200 | 1.58 ± 0.47 | 504 |
| Cyclohexane, 1,4-dimethoxy-2-methyl-, stereoisomer | C9H18O2 | 17.183 | 0.44 ± 0.08 | 158 | Cyclohexanone, 5-(1-hydroxy-2-propenyl)-2,2-dimethyl-, | C11H18O2 | 17.452 | 1.50 ± 0.28 | 182 |
| Isobornyl acetate | C12H20O2 | 21.144 | 0.32 ± 0.09 | 196 | 2-Nitrohept-2-en-1-ol | C7H13NO3 | 9.739 | 1.03 ± 0.27 | 159 |
| 1,5,5-Trimethyl-6-methylene-cyclohexene | C10H16 | 15.168 | 0.30 ± 0.04 | 136 | E-9-Methyl-8-tridecen-2-ol, acetate | C16H30O2 | 17.686 | 0.67 ± 0.30 | 254 |
| Organ | Solvent | TP (mg/100 g FW) | TF (mg/100 g FW) | TAA (%) |
|---|---|---|---|---|
| Leaf | Methanol | 0.40 ± 0.01 c | 0.31 ± 0.01 c | 0.63 ± 0.02 bcd |
| Ethanol | 0.38 ± 0.01 d | 0.28 ± 0.01 d | 0.61 ± 0.02 d | |
| Tuber | Methanol | 0.49 ± 0.01 a | 0.39 ± 0.01 a | 0.76 ± 0.01 a |
| Ethanol | 0.44 ± 0.01 b | 0.33 ± 0.01 b | 0.64 ± 0.01 bc |
| Trait | Value |
|---|---|
| Moisture (%) | 11.55 ± 0.70 |
| Ash (g/100g) | 1.76 ± 0.05 |
| Protein (g/100g) | 3.95 ± 0.04 |
| Starch (%) | 31.31 ± 0.60 |
| Glucomannan (%) | 36.96 ± 1.22 |
| Viscosity (cP) | 17.96 ± 0.85 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fatahi, M.; Vafaee, Y.; Tahir, N.A.-r.; Khorshidi, J. Large-Scale In Vitro Multiplication and Phytochemical Analysis of Himantoglossum affine (Boiss.) Schltr.: An Endangered Euro-Mediterranean Terrestrial Orchid. Diversity 2022, 14, 1137. https://doi.org/10.3390/d14121137
Fatahi M, Vafaee Y, Tahir NA-r, Khorshidi J. Large-Scale In Vitro Multiplication and Phytochemical Analysis of Himantoglossum affine (Boiss.) Schltr.: An Endangered Euro-Mediterranean Terrestrial Orchid. Diversity. 2022; 14(12):1137. https://doi.org/10.3390/d14121137
Chicago/Turabian StyleFatahi, Mozhgan, Yavar Vafaee, Nawroz Abdul-razzak Tahir, and Jalal Khorshidi. 2022. "Large-Scale In Vitro Multiplication and Phytochemical Analysis of Himantoglossum affine (Boiss.) Schltr.: An Endangered Euro-Mediterranean Terrestrial Orchid" Diversity 14, no. 12: 1137. https://doi.org/10.3390/d14121137