Simple and Divided Leaves in Ferns: Exploring the Genetic Basis for Leaf Morphology Differences in the Genus Elaphoglossum (Dryopteridaceae)


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Evolutionary History of Class I KNOX Transcription Factors in Ferns
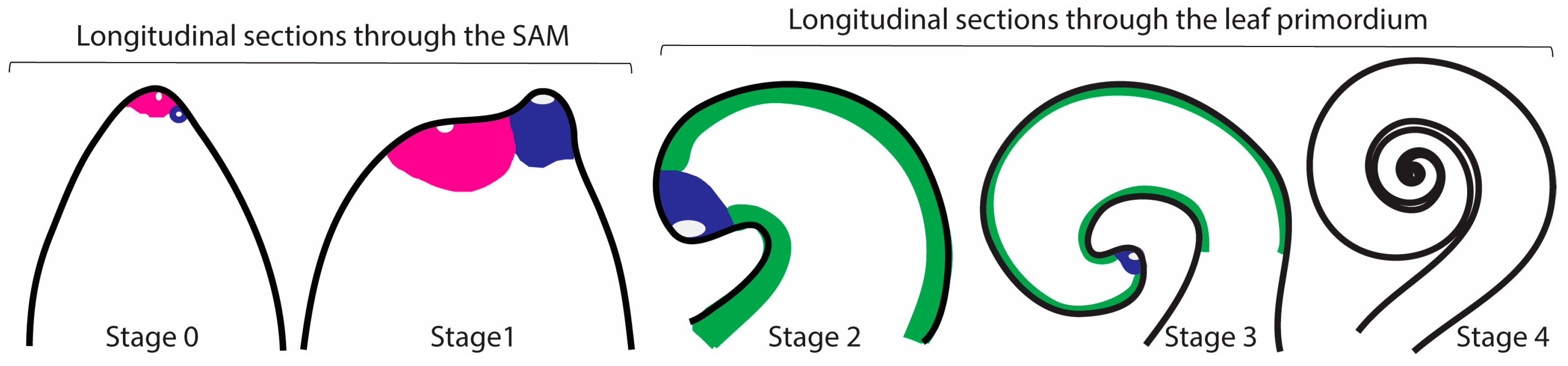
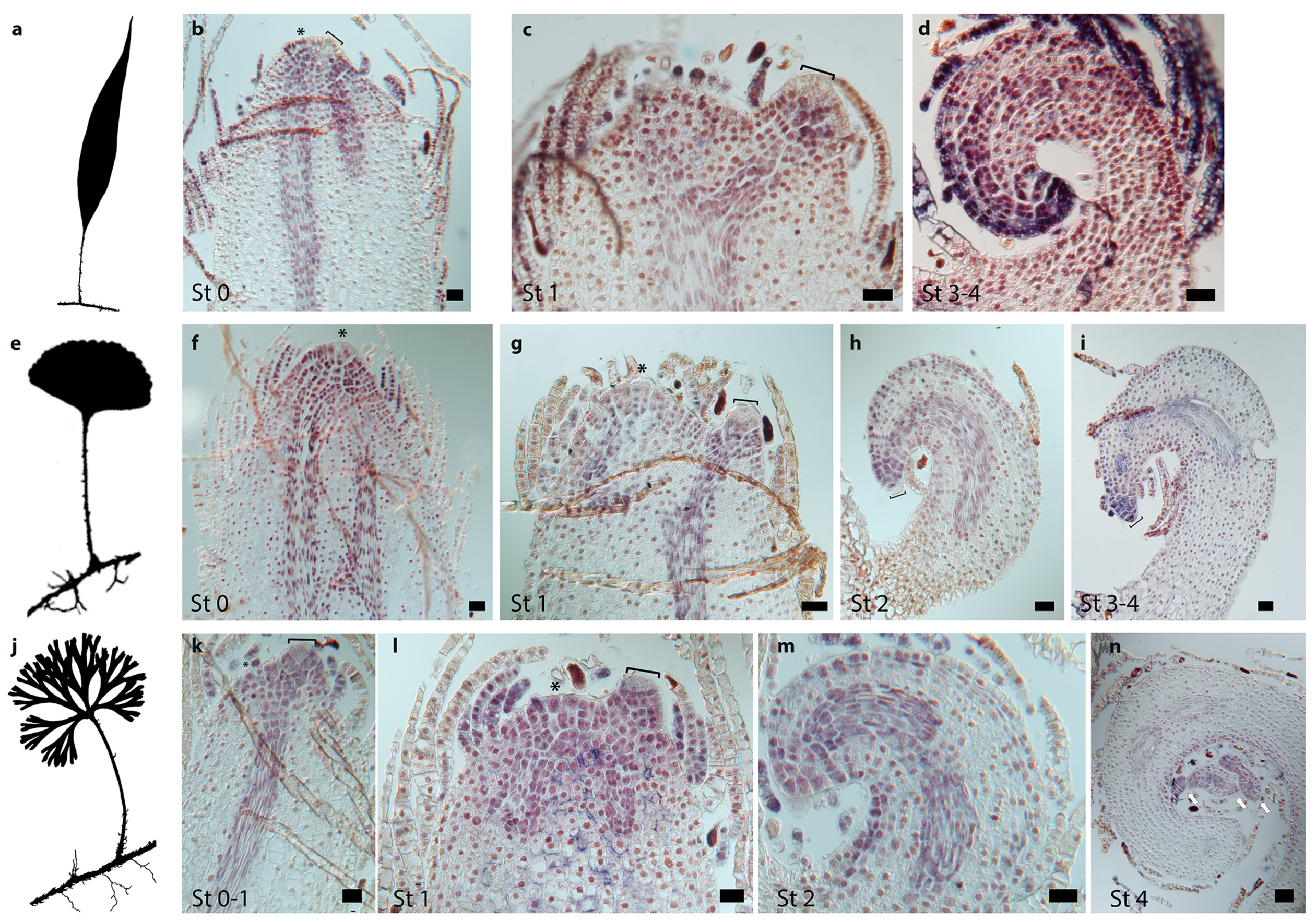
2.2. Development of Simple and Divided Leaves of Elaphoglossum
2.3. Patterns of Cell Division in Simple and Divided Leaves of Elaphoglossum
2.4. Class I KNOX Gene Expression Patterns in Simple and Divided Leaves of the Fern Genus Elaphoglossum
3. Discussion
3.1. Evolutionary History of Class I KNOX Transcription Factors in Ferns
3.2. Class I KNOX Genes Are Expressed in Shoot and Leaf Fern Meristems
3.3. Class I KNOX Gene Expression in Fern Leaves Recapitulates Shoot Expression
3.4. Development of Simple and Divided Leaves in Ferns
3.5. A Conserved Mechanism of Leaf Dissection in Megaphylls
3.6. Class I KNOX Genes and Megaphyll Evolution
4. Materials and Methods
4.1. Sampling for the Phylogenetic Analyses of Class I KNOX Genes
4.2. Sequence Analysis, Alignment, and Phylogenetic Analysis
4.3. Taxonomic Sampling for Gene Expression Studies
4.4. RNA and DNA Extraction and cDNA Synthesis
4.5. Primer Design and PCR
4.6. In Situ Hybridization
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| C1KNOX | Class I KNOTTED1-like HOMEOBOX |
| LAI | Leaf Apical Initial |
| LAM | Leaf Apical Meristem |
| MM | Marginal Meristem |
| RAM | Root Apical Meristem |
| SAM | Shoot Apical Meristem |
Appendix A
Appendix B
References
- Gifford, E.M.; Foster, A.S. Morphology and Evolution of Vascular Plants, 3rd ed.; W.H. Freeman and Co.: New York, NY, USA, 1989; ISBN 0716719460. [Google Scholar]
- Beerling, D.J. Leaf Evolution: Gases, Genes and Geochemistry. Ann. Bot. 2005, 96, 345–352. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boyce, C.K.; Knoll, A.H. Evolution of developmental potential and the multiple independent origins of leaves in Paleozoic vascular plants. Paleobiology 2002, 28, 70–100. [Google Scholar] [CrossRef]
- Boyce, C.K. The evolution of plant development in a paleontological context. Curr. Opin. Plant Biol. 2010, 13, 102–107. [Google Scholar] [CrossRef] [PubMed]
- Sanders, H.; Rothwell, G.W.; Wyatt, S.E. Key morphological alterations in the evolution of leaves. Int. J. Plant Sci. 2009, 170, 860–868. [Google Scholar] [CrossRef]
- Tomescu, A.M.F. Megaphylls, microphylls and the evolution of leaf development. Trends Plant Sci. 2009, 14, 5–12. [Google Scholar] [CrossRef]
- Vasco, A.; Moran, R.C.; Ambrose, B.A. The evolution, morphology, and development of fern leaves. Front. Plant Sci. 2013, 4, 345. [Google Scholar] [CrossRef] [Green Version]
- Harrison, C.J.; Morris, J.L. The origin and early evolution of vascular plant shoots and leaves. Philos. Trans. R. Soc. B Biol. Sci. 2017, 373, 20160496. [Google Scholar] [CrossRef]
- Bharathan, G.; Goliber, T.E.; Moore, C.; Kessler, S.; Pham, T.; Sinha, N.R. Homologies in leaf form inferred from KNOXI gene expression during development. Science 2002, 296, 1858–1860. [Google Scholar] [CrossRef]
- Harrison, C.J.; Corley, S.B.; Moylan, E.C.; Alexander, D.L.; Scotland, R.W.; Langdale, J.A. Independent recruitment of a conserved developmental mechanism during leaf evolution. Nature 2005, 434, 509–514. [Google Scholar] [CrossRef]
- Floyd, S.K.; Bowman, J.L. Gene expression patterns in seed plant shoot meristems and leaves: Homoplasy or homology? J. Plant Res. 2010, 123, 43–55. [Google Scholar] [CrossRef]
- Bar, M.; Ori, N. Compound leaf development in model plant species. Curr. Opin. Plant Biol. 2015, 23, 61–69. [Google Scholar] [CrossRef]
- Du, F.; Guan, C.; Jiao, Y. Molecular Mechanisms of Leaf Morphogenesis. Mol. Plant 2018, 11, 1117–1134. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vollbrecht, E.; Veit, B.; Sinha, N.R.; Hake, S. The developmental gene Knotted-1 is a member of a maize homeobox gene family. Nature 1991, 350, 241–243. [Google Scholar] [CrossRef] [PubMed]
- Jackson, D.; Veit, B.; Hake, S. Expression of maize KNOTTED1 related homeobox genes in the shoot apical meristem predicts patterns of morphogenesis in the vegetative shoot. Development 1994, 120, 405–413. [Google Scholar]
- Kerstetter, R.; Vollbrecht, E.; Lowe, B.; Veit, B.; Yamaguchi, J.; Hake, S. Sequence analysis and expression patterns divide the maize knotted1-like homeobox genes into two classes. Plant Cell 1994, 6, 1877–1887. [Google Scholar]
- Lincoln, C.; Long, J.; Yamaguchi, J.; Serikawa, K.; Hake, S. A knotted1-like homeobox gene in Arabidopsis is expressed in the vegetative meristem and dramatically alters leaf morphology when overexpressed in transgenic plants. Plant Cell 1994, 6, 1859–1876. [Google Scholar]
- Endrizzi, K.; Moussian, B.; Haecker, A.; Levin, J.Z.; Laux, T. The SHOOT MERISTEMLESS gene is required for maintenance of undifferentiated cells in Arabidopsis shoot and floral meristems and acts at a different regulatory level than the meristem genes WUSCHEL and ZWILLE. Plant J. 1996, 10, 967–979. [Google Scholar] [CrossRef]
- Long, J.A.; Moan, E.I.; Medford, J.I.; Barton, M.K. A member of the KNOTTED class of homeodomain proteins encoded by the STM gene of Arabidopsis. Nature 1996, 379, 66–69. [Google Scholar] [CrossRef]
- Nishimura, A.; Tamaoki, M.; Sato, Y.; Matsuoka, M. The expression of tobacco knotted1-type class 1 homeobox genes correspond to regions predicted by the cytohistological zonation model. Plant J. 1999, 18, 337–347. [Google Scholar] [CrossRef] [Green Version]
- Reiser, L.; Sanchez-Baracaldo, P.; Hake, S.; Sánchez-Baracaldo, P. Knots in the family tree: Evolutionary relationships and functions of knox homeobox genes. Plant Mol. Biol. 2000, 42, 151–166. [Google Scholar] [CrossRef]
- Hay, A.; Tsiantis, M. The genetic basis for differences in leaf form between Arabidopsis thaliana and its wild relative Cardamine hirsuta. Nat. Genet. 2006, 38, 942–947. [Google Scholar] [CrossRef]
- Kimura, S.; Koenig, D.; Kang, J.; Yoong, F.Y.; Sinha, N.R. Natural variation in leaf morphology results from mutation of a novel KNOX gene. Curr. Biol. 2008, 672–677. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hareven, D.; Gutfinger, T.; Parnis, A.; Eshed, Y.; Lifschitz, E. The making of a compound leaf: Genetic manipulation of leaf architecture in tomato. Cell 1996, 84, 735–744. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.J.; Janssen, B.J.; Williams, A.; Sinha, N.R. A gene fusion at a homeobox locus: Alterations in leaf shape and implications for morphological evolution. Plant Cell 1997, 9, 1289–1304. [Google Scholar] [PubMed] [Green Version]
- Janssen, B.J.; Williams, A.; Chen, J.J.; Mathern, J.; Hake, S.; Sinha, N.R. Isolation and characterization of two knotted-like homeobox genes from tomato. Plant Mol. Biol. 1998, 36, 417–425. [Google Scholar] [CrossRef] [PubMed]
- Sinha, N.R.; Williams, R.E.; Hake, S. Overexpression of the maize homeo box gene, KNOTTED-1, causes a switch from determinate to indeterminate cell fates. Genes Dev. 1993, 7, 787–795. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ori, N.; Eshed, Y.; Chuck, G.; Bowman, J.L.; Hake, S. Mechanisms that control knox gene expression in the Arabidopsis shoot. Development 2000, 127, 5523–5532. [Google Scholar]
- Koenig, D.; Bayer, E.; Kang, J.; Kuhlemeier, C.; Sinha, N.R. Auxin patterns Solanum lycopersicum leaf morphogenesis. Development 2009, 136, 2997–3006. [Google Scholar] [CrossRef] [Green Version]
- Sundås-Larsson, A.; Svenson, M.; Liao, H.; Engström, P. A homeobox gene with potential developmental control function in the meristem of the conifer Picea abies. Proc. Natl. Acad. Sci. USA 1998, 95, 15118–15122. [Google Scholar] [CrossRef] [Green Version]
- Pham, T.; Sinha, N.R. Role of Knox Genes in Shoot Development of Welwitschia mirabilis. Int. J. Plant Sci. 2003, 164, 333–343. [Google Scholar] [CrossRef]
- Sano, R.; Juárez, C.M.; Hass, B.; Sakakibara, K.; Ito, M.; Banks, J.A.; Hasebe, M.; Juarez, C.M. KNOX homeobox genes potentially have similar function in both diploid unicellular and multicellular meristems, but not in haploid meristems. Evol. Dev. 2005, 7, 69–78. [Google Scholar] [CrossRef] [PubMed]
- Ambrose, B.A.; Vasco, A. Bringing the multicellular fern meristem into focus. New Phytol. 2016, 210, 790–793. [Google Scholar] [CrossRef]
- Mickel, J.T.; Smith, A.R. The pteridophytes of Mexico. Mem. N. Y. Bot. Gard. 2004, 88, 1–1055. [Google Scholar]
- Vasco, A.; Mickel, J.T.; Moran, R.C. Taxonomic revision of the neotropical species of Elaphoglossum Sect. Squamipedia (Dryopteridaceae). Ann. Missouri Bot. Gard. 2013, 99, 244–286. [Google Scholar] [CrossRef]
- Vasco, A.; Lóriga, J.; Rouhan, G.; Ambrose, B.A.; Moran, R.C. Divided leaves in the genus Elaphoglossum (Dryopteridaceae): A phylogeny of Elaphoglossum section Squamipedia. Syst. Bot. 2015, 40, 46–55. [Google Scholar] [CrossRef]
- Matos, F.B.; Vasco, A.; Moran, R.C. Elaphoglossum doanense and Elaphoglossum tonduzii: New members of Elaphoglossum Sect. Squamipedia (Dryopteridaceae) and their significance for inferring the evolution of rhizome habit and nest-forming leaves within the genus. Int. J. Plant Sci. 2018, 179, 296–313. [Google Scholar] [CrossRef]
- Moran, R.C.; Labiak, P.H.; Sundue, M. Phylogeny and character evolution of the bolbitidoid ferns (Dryopteridaceae). Int. J. Plant Sci. 2010, 171, 547–559. [Google Scholar] [CrossRef]
- Dinneny, J.R.; Yadegari, R.; Fischer, R.L.; Yanofsky, M.F.; Weigel, D. The role of JAGGED in shaping lateral organs. Development 2004, 131, 1101–1110. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- von Willdenow, C.L. Species Plantarum. Editio quarta 5; G.C. Nauk: Berlin, Germany, 1810. [Google Scholar]
- Christ, H. Monographie des Genus Elaphoglossum. Neue Denkschriften der Allg. Schweizerischen Gesellschaft für die gesammten Naturwissenschaften 1899, 36, 1–159. [Google Scholar]
- Morton, C.V. Notes on Elaphoglossum, III. The publication of Elaphoglossum and Rhipidopteris. Am. Fern J. 1955, 45, 11–14. [Google Scholar] [CrossRef]
- Gómez, L.D. Contribuciones a la pteridología costarricense. VI. El género Peltapteris Link en Costa Rica. Brenesia 1975, 6, 25–31. [Google Scholar]
- Mickel, J.T. Relationships of the dissected elaphoglossoid ferns. Brittonia 1980, 32, 109–117. [Google Scholar] [CrossRef]
- Steeves, T.A.; Sussex, I.M. Patterns in Plant Development; Cambridge University Press: New York, NY, USA, 1989. [Google Scholar]
- Wardlaw, C.W. Experimental and analytical studies of pteridophytes: XIV. Leaf formation and phyllotaxis in Dryopteris aristata Druce. Ann. Bot. 1949, 13, 163–198. [Google Scholar] [CrossRef]
- Saha, B. Morphogenetic studies on the distribution and activities of leaf meristems in ferns. Ann. Bot. 1963, 27, 269–279. [Google Scholar] [CrossRef]
- Zurakowski, K.A.; Gifford, E.M. Quantitative studies of pinnule development in the ferns Adaiantum raddiantum and Cheilanthes viridis. Am. J. Bot. 1988, 75, 1559–1570. [Google Scholar] [CrossRef]
- Bierhorst, D.W. On the stem apex, leaf initiation and early leaf ontogeny in filicalean ferns. Am. J. Bot. 1977, 64, 125–152. [Google Scholar] [CrossRef]
- Bower, F.O. The Ferns (Filicales) Treated Comparatively with a View to Their Natural Classification; Cambridge University Press: Cambridge, UK, 1923. [Google Scholar]
- Prigge, M.J.; Otsuga, D.; Alonso, J.M.; Ecker, J.R.; Drews, G.N.; Clark, S.E. Class III homeodomain-leucine zipper gene family members have overlapping, antagonistic, and distinct roles in Arabidopsis development. Plant Cell 2005, 17, 61–76. [Google Scholar] [CrossRef] [Green Version]
- Vasco, A.; Smalls, T.L.; Graham, S.W.; Cooper, E.D.; Wong, G.K.-S.; Stevenson, D.W.; Moran, R.C.; Ambrose, B.A. Challenging the paradigms of leaf evolution: Class III HD-Zips in ferns and lycophytes. New Phytol. 2016, 212, 745–758. [Google Scholar] [CrossRef]
- PPG I A community-derived classification for extant lycophytes and ferns. J. Syst. Evol. 2016, 54, 560–603.
- Li, F.-W.; Brouwer, P.; Carretero-Paulet, L.; Cheng, S.; de Vries, J.; Delaux, P.-M.; Eily, A.; Koppers, N.; Kuo, L.-Y.; Li, Z.; et al. Fern genomes elucidate land plant evolution and cyanobacterial symbioses. Nat. Plants 2018, 4, 460–472. [Google Scholar] [CrossRef] [Green Version]
- Rothfels, C.J.; Li, F.-W.; Sigel, E.M.; Huiet, L.; Larsson, A.; Burge, D.O.; Ruhsam, M.; Deyholos, M.; Soltis, D.E.; Stewart, C.N.; et al. The evolutionary history of ferns inferred from 25 low-copy nuclear genes. Am. J. Bot. 2015, 102, 1089–1107. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leebens-Mack, J.H.; Barker, M.S.; Carpenter, E.J.; Deyholos, M.K.; Gitzendanner, M.A.; Graham, S.W.; Grosse, I.; Li, Z.; Melkonian, M.; Mirarab, S.; et al. One thousand plant transcriptomes and the phylogenomics of green plants. Nature 2019, 574, 679–685. [Google Scholar]
- Wardlaw, C.W. Experimental studies of the sporophytes of ferns. J. Linn. Soc. London, Bot. 1963, 8, 385–400. [Google Scholar] [CrossRef]
- White, R.; Turner, M. Anatomy and development of the fern sporophyte. Bot. Rev. 1995, 61, 281–305. [Google Scholar] [CrossRef]
- Imaichi, R. Meristem organization and organ diversity. In Biology and Evolution of Ferns and Lycophytes; Ranker, T.A., Haufler, C.H., Eds.; Cambridge University Press: Cambridge, UK, 2008; pp. 75–103. [Google Scholar]
- Bower, F.O. The comparative examination of the meristems of ferns, as a phylogenetic study. Ann. Bot. 1889, 3, 305–392. [Google Scholar] [CrossRef]
- Pray, T.R. Ontogeny of the open dichotomous venation in the pinna of the fern Nephrolepis. Am. J. Bot. 1960, 47, 319–328. [Google Scholar] [CrossRef]
- Pray, T.R. Ontogeny of the Closed Dichotomous Venation of Regnellidium. Am. J. Bot. 1962, 49, 464–472. [Google Scholar] [CrossRef]
- Wardlaw, C.W. Reflections on the unity of the embryonic tissues in ferns. Phytomorphology 1958, 8, 323–327. [Google Scholar]
- Gaudet, J.J. Morphology of Marsilea vestita. II. Morphology of the Adult Land and Submerged Leaves. Am. J. Bot. 1964, 51, 591–597. [Google Scholar] [CrossRef]
- Steeves, T.A. A study of the developmental potentialities of excised leaf primordia in sterile culture. Phytomorphology 1961, 11, 346–359. [Google Scholar]
- Steeves, T.A. Morphogenetic studies of fern leaves. J. Linn. Soc. London, Bot. 1963, 58, 401–415. [Google Scholar] [CrossRef]
- Wardlaw, C.W. Experiments on organogenesis in ferns. Growth 1949, 13, 93–131. [Google Scholar]
- Wardlaw, C.W. Phyllotaxis and organogenesis in ferns. Nature 1949, 164, 167–169. [Google Scholar] [CrossRef]
- Cutter, E.G. Experimental induction of buds from fern leaf primordia. Nature 1954, 173, 440–441. [Google Scholar] [CrossRef]
- Cutter, E.G. Experimental and analytical studies of pteridophytes: XXXIII. The experimental induction of buds from leaf primordia in Dryopteris aristata Druce. Ann. Bot. 1956, 20, 143–165. [Google Scholar] [CrossRef]
- White, R. Experimental and developmental studies of the fern sporophyte. Bot. Rev. 1971, 37, 509–540. [Google Scholar] [CrossRef]
- Steeves, T.A.; Hicks, G.; Steeves, M.; Retallack, B. Leaf determination in the Fern Osmunda cinnamomea—A reinvestigation. Ann. Bot. 1993, 71, 511–517. [Google Scholar] [CrossRef]
- Sanders, H.L.; Darrah, P.R.; Langdale, J. a Sector analysis and predictive modelling reveal iterative shoot-like development in fern fronds. Dev. Cambridge Engl. 2011, 138, 2925–2934. [Google Scholar] [CrossRef] [Green Version]
- Plackett, A.R.G.; Di Stilio, V.S.; Langdale, J.A. Ferns: The missing link in shoot evolution and development. Front. Plant Sci. 2015, 6, 972. [Google Scholar] [CrossRef] [Green Version]
- Arber, A. The interpretation of leaf and root in the angiosperms. Biol. Rev. 1941, 16, 81–105. [Google Scholar] [CrossRef]
- Arber, A. The Natural Philosophy of Plant Form; Cambridge University Press: Cambridge, UK, 1950. [Google Scholar]
- Hay, A.; Tsiantis, M. KNOX genes: Versatile regulators of plant development and diversity. Development 2010, 137, 3153–3165. [Google Scholar] [CrossRef] [Green Version]
- Barkoulas, M.; Hay, A.; Kougioumoutzi, E.; Tsiantis, M. A developmental framework for dissected leaf formation in the Arabidopsis relative Cardamine hirsuta. Nat. Genet. 2008, 40, 1136–1141. [Google Scholar] [CrossRef] [PubMed]
- Blein, T.; Pulido, A.; Vialette-Guiraud, A.; Nikovics, K.; Morin, H.; Hay, A.; Johansen, I.E.; Tsiantis, M.; Laufs, P. A Conserved Molecular Framework for Compound Leaf Development. Science 2008, 322, 1835–1839. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peng, J.; Chen, R. Auxin efflux transporter MtPIN10 regulates compound leaf and flower development in Medicago truncatula. Plant Signal. Behav. 2011, 6, 1537–1544. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Byrne, M.E. Making leaves. Curr. Opin. Plant Biol. 2012, 15, 24–30. [Google Scholar] [CrossRef] [PubMed]
- Reinhardt, D.; Mandel, T.; Kuhlemeier, C. Auxin Regulates the Initiation and Radial Position of Plant Lateral Organs. Plant Cell 2000, 12, 507–518. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Benková, E.; Michniewicz, M.; Sauer, M.; Teichmann, T.; Seifertová, D.; Jürgens, G.; Friml, J. Local, Efflux-Dependent Auxin Gradients as a Common Module for Plant Organ Formation. Cell 2003, 115, 591–602. [Google Scholar] [CrossRef] [Green Version]
- Berger, Y.; Harpaz-Saad, S.; Brand, A.; Melnik, H.; Sirding, N.; Alvarez, J.P.; Zinder, M.; Samach, A.; Eshed, Y.; Ori, N. The NAC-domain transcription factor GOBLET specifies leaflet boundaries in compound tomato leaves. Development 2009, 136, 823–832. [Google Scholar] [CrossRef] [Green Version]
- Floyd, S.K.; Bowman, J.L. Distinct developmental mechanisms reflect the independent origins of leaves in vascular plants. Curr. Biol. 2006, 16, 1911–1917. [Google Scholar] [CrossRef] [Green Version]
- Prigge, M.J.; Clark, S.E. Evolution of the class III HD-Zip gene family in land plants. Evol. Dev. 2006, 8, 350–361. [Google Scholar] [CrossRef]
- Evkaikina, A.I.; Berke, L.; Romanova, M.A.; Proux-Wéra, E.; Ivanova, A.N.; Rydin, C.; Pawlowski, K.; Voitsekhovskaja, O.V. The Huperzia selago shoot tip transcriptome sheds new light on the evolution of leaves. Genome Biol. Evol. 2017, 9, 2444–2460. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frangedakis, E.; Saint-Marcoux, D.; Moody, L.A.; Rabbinowitsch, E.; Langdale, J.A. Nonreciprocal complementation of KNOX gene function in land plants. New Phytol. 2017, 216, 591–604. [Google Scholar] [CrossRef]
- Katoh, K.; Standley, D.M. MAFFT multiple sequence alignment software version 7: Improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maddison, W.P.; Maddison, D.R. Mesquite: A modular System for Evolutionary Analysis. Version 3.61. Available online: http://mesquiteproject.org (accessed on 21 July 2020).
- Miller, M.A.; Pfeiffer, W.; Schwartz, T. Creating the CIPRES Science Gateway for inference of large phylogenetic trees. In Proceedings of the Gateway Computing Environments Workshop (GCE), New Orleans, LA, USA, 14 November 2010; pp. 1–8. [Google Scholar]
- Lanfear, R.; Frandsen, P.B.; Wright, A.M.; Senfeld, T.; Calcott, B. PartitionFinder 2: New methods for selecting partitioned models of evolution for molecular and morphological phylogenetic analyses. Mol. Biol. Evol. 2017, 34, 772–773. [Google Scholar] [CrossRef] [Green Version]
- Ronquist, F.R.; Huelsenbeck, J.P. MrBayes: Bayesian inference of phylogeny. Bioinformatics 2003, 19, 1572–1574. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cone, K.C.; Burr, F.A.; Burr, B. Molecular analysis of the maize anthocyanin locus C1. Proc. Natl. Acad. Sci. USA 1986, 83, 9631–9635. [Google Scholar] [CrossRef] [Green Version]
- Ambrose, B.A.; Lerner, D.R.; Ciceri, P.; Padilla, C.M.; Yanofsky, M.F.; Schmidt, R.J. Molecular and genetic analyses of the Silky1 gene reveal conservation in floral organ specification between eudicots and monocots. Mol. Cell 2000, 5, 569–579. [Google Scholar] [CrossRef]
- Torres, M.-A.; Rigau, J.; Puigdomenech, P.; Stiefel, V. Specific distribution of mRNAs in maize growing pollen tubes observed by whole-mount in situ hybridization with non-radioactive probes. Plant J. 1995, 8, 317–321. [Google Scholar] [CrossRef]





© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vasco, A.; Ambrose, B.A. Simple and Divided Leaves in Ferns: Exploring the Genetic Basis for Leaf Morphology Differences in the Genus Elaphoglossum (Dryopteridaceae). Int. J. Mol. Sci. 2020, 21, 5180. https://doi.org/10.3390/ijms21155180
Vasco A, Ambrose BA. Simple and Divided Leaves in Ferns: Exploring the Genetic Basis for Leaf Morphology Differences in the Genus Elaphoglossum (Dryopteridaceae). International Journal of Molecular Sciences. 2020; 21(15):5180. https://doi.org/10.3390/ijms21155180
Chicago/Turabian StyleVasco, Alejandra, and Barbara A. Ambrose. 2020. "Simple and Divided Leaves in Ferns: Exploring the Genetic Basis for Leaf Morphology Differences in the Genus Elaphoglossum (Dryopteridaceae)" International Journal of Molecular Sciences 21, no. 15: 5180. https://doi.org/10.3390/ijms21155180