|
|
In: Van Driesche, R., et al., 2002, Biological Control of Invasive Plants in the Eastern United States, USDA Forest Service Publication FHTET-2002-04, 413 p. Pest Status of Weed Waterhyacinth, Eichhornia crassipes (Mart.) Solms.-Laubach (Fig. 1), is considered one of the world’s worst weeds (Holm et al., 1977), invading lakes, ponds, canals, and rivers. It was introduced into many countries during the late 19th and early 20th centuries, where it spread and degraded aquatic ecosystems. It is still rapidly spreading throughout Africa, where new infestations are creating life-threatening situations as well as environmental and cultural upheaval (Cock et al., 2000). Control with herbicides, particularly 2,4-D, is feasible, but is costly and temporary.
Nature of Damage Economic damage. Waterhyacinth grows rapidly (Penfound and Earle, 1948) forming expansive colonies of tall, interwoven floating plants. It blankets large waterbodies (Fig. 2), creating impenetrable barriers and obstructing navigation (Gowanloch and Bajkov, 1948; Zeiger, 1962). Floating mats block drainage, causing flooding or preventing subsidence of floodwaters. Large rafts accumulate where water channels narrow, sometimes causing bridges to Collapse. Waterhyacinth hinders irrigation by impeding water flow, by clogging irrigation pumps, and by interfering with weirs (Penfound and Earle, 1948). Multimillion-dollar flood control and water supply projects can be rendered useless by waterhyacinth infestations (Gowanloch and Bajkov, 1948). Infestations block access to recreational areas and decrease waterfront property values, oftentimes harming the economies of communities that depend upon fishing and water sports for revenue. Shifting waterhyacinth mats sometimes prevent boats from reaching shore, trapping the occupants and exposing them to environmental hazards (Gowanloch and Bajkov, 1948; Harley, 1990). Waterhyacinth infestations intensify mosquito problems by hindering insecticide application, interfering with predators, increasing habitat for species that attach to plants, and impeding runoff and water circulation (Seabrook, 1962). Ecological damage. Dense mats reduce light to submerged plants, thus depleting oxygen in aquatic communities (Ultsch, 1973). The resultant lack of phytoplankton (McVea and Boyd, 1975) alters the composition of invertebrate communities (O’Hara, 1967; Hansen et al., 1971), ultimately affecting fisheries. Drifting mats scour vegetation, destroying native plants and wildlife habitat. Waterhyacinth also Competes with other plants, often displacing wildlife forage and habitat (Gowanloch, 1944). Higher sediment loading occurs under waterhyacinth mats due to increased detrital production and siltation. Herbicidal treatment or mechanical harvesting of waterhyacinth often damages nearby desirable vegetation. Extent of losses. Waterhyacinth caused annual losses (all causes) of $65 to 75 million in Louisiana during the 1940s (Gowanloch and Bajkov, 1948). Fish and wildlife losses alone in the six southeastern states exceeded $4 million per year in 1947 and waterhyacinth control provided a benefit to Cost ratio of 15.3:1 (Tabita and Woods, 1962). Holm et al. (1969) ascribed losses of $43 million in 1956 to waterhyacinth infestations in Florida, Mississippi, Alabama, and Louisiana. The U.S. Army Corps of Engineers estimated benefits from waterhyacinth control programs at nearly $14 million in 1965 (Gordon and Coulson, 1974). Florida spent more than $43 million during 1980 to 1991 to suppress waterhyacinth and waterlettuce (Schmitz et al., 1993). Currently, annual costs for waterhyacinth management range from $500,000 in California to $3 million in Florida (Mullin et al., 2000). The largest infestations occur in Louisiana, where the Department of Fisheries herbicidally treats about 25,000 acres of waterhyacinth per year, mostly at boat ramps, at an annual cost of $2 million (R. Brassette, pers. comm.).
Taxonomy The English common names of the plant are waterhyacinth, water hyacinth, and water-hyacinth. Waterhyacinth is the standardized spelling adopted by the Weed Science Society of America (WSSA, 1984) to denote that it is not an aquatic relative of true “hyacinth” (Hyacinthus spp.), as the two-word spelling suggests. The taxonomic placement of waterhyacinth, based on Cronquist (1988), Thorne (1992), and Takhtajan (1997), is as follows: division Magnoliophyta; class Liliopsida; subclass Commelinidae (Liliidae [Cronquist, 1988; Thorne, 1992]); superorder Commelinanae (Thorne, 1992); order Pontederiales (Liliales [Cronquist, 1988]; Philydrales [Thorne, 1992]); family Pontederiaceae, genus Eichhornia; specific epithet crassipes (Martius) Solms-Laubach.
sediments for 15 to 20 years (Matthews, 1967; Gopal, 1987). Seeds germinate on moist sediments or in warm shallow water (Haigh, 1936; Hitchcock et al., 1950) and flowering can occur 10 to 15 weeks thereafter (Barrett, 1980). Lack of germination sites limits seedling recruitment except during drought, on decaying mats after herbicide applications (Matthews, 1967), or at the margins of waterbodies. Populations increase mainly by vegetative means.
Weber (1950), Richards (1982), Watson (1984), and Watson and Cook (1982, 1987) describe waterhyacinth growth and population expansion as the result of differentiation of apical or axillary meristems. The single apical meristem on each stem tip can be vegetative, producing leaves with axillary buds, or reproductive, producing flowers. If an inflorescence develops, termination of the apical meristem halts leaf production. In this event, the axillary bud immediately below the inflorescence differentiates into a continuation shoot. This produces a new apical meristem that allows leaf production to proceed. If the axillary bud doesn’t form a continuation shoot, then it produces a stolon. Elongation of the stolon internode moves the axillary bud apex away from the parent rosette. It then produces short internodes that grow vertically into a new rosette. Waterhyacinth grows best in neutral pH, water high in macronutrients, warm temperatures (28° to 30°C), and high light intensities. It tolerates pH levels from 4.0 to 10.0 (Haller and Sutton, 1973), but not more than 20 to 25% sea water (Muramoto et al., 1991). The plants survive frost if the rhizomes don’t freeze, even though emergent portions may succumb (Webber, 1897). Prolonged cold kills the plants (Penfound and Earle, 1948), but reinfestation from seed follows during later warmer periods. Ueki (1978) matched the northern limit of waterhyacinth to the 1o C average January isotherm in Japan. Growth is inhibited at water temperatures above 33°C (Knipling et al., 1970). Plants stranded on moist sediments can survive several months (Parija, 1934). Analysis of Related Native Plants in the Eastern United States Waterhyacinth is a member of the pickerelweed family (Pontederiaceae). Families most closely allied with the Pontederiaceae are Commelinaceae, Haemodoraceae (including Conostylidaceae [Takhtajan, 1997]), Philydraceae, and Hanguanaceae (Hahn, 1997; APG, 1998). The subclass Commelinidae includes the Arecales, Poales, Commelinales, and Zingiberales (APG, 1998). The Pontederiaceae is a small family of herbaceous mono Cotyledons that includes six genera and 30 to 35 species (Eckenwalder and Barrett, 1986). All are palustrine or aquatic and most are confined to the Americas. All seven members of the genus Eichhornia originated in tropical America, except for Eichhornia natans (P. Beauv.), which is from tropical Africa. Fourteen species of Pontederiaceae occur in the U.S./Canadian flora (Table 1), six of which are adventive; none are considered threatened or endangered (USDA, NRCS, 1999). Table 1. Species of Pontederiaceae in the United States.
History of Biological Control Efforts in the Eastern United States Area of Origin of Weed The diversity of other species of Eichhornia, particularly the more primitive Eichhornia paniculata (Spreng.) Solms. and Eichhornia paradoxa (Mart.) Solms., and the overlapping range of the closely related Eichhornia azurea (Sw.) Kunth suggest that E. crassipes arose in tropical South America. Areas Surveyed for Natural Enemies Although several expeditions have been made to South America to survey for natural enemies of waterhyacinth (Center, 1994), most were limited in scope and failed to encompass the upper Amazon basin where waterhyacinth may have originated. Bennett and Zwölfer (1968) explored the northernmost range of the plant. Other authors have explored the eastern parts of the range but the western portion has seldom been visited. The discovery of new organisms associated with waterhyacinth was thought to be unlikely because of the long history of exploration in South America. Recent findings of new, potentially useful natural enemies suggest otherwise (Cordo, 1999). Natural Enemies Found Beginning in the early 1970s, the USDA and CIBC (now CABI-Bioscience) released the weevils Neochetina eichhorniae Warner, Neochetina bruchi Hustache, and, later, the pyralid moth Niphograpta (=Sameodes) albiguttalis (Warren). These three agents, plus the mite Orthogalumna terebrantis Wallwork, are now widely used (Table 2). Many countries that have initiated biological control programs against waterhyacinth have reported successes (Julien and Griffiths, 1998). All four agents are important, although the two Neochetina weevils seem most successful. Nonetheless, the control achieved has not always been sufficient. The relatively slow action of the biological control agents is sometimes incompatible with other management practices (Center et al., 1999a). In other cases, the explosive growth of waterhyacinth stimulated by high nutrient levels precludes effective control (Heard and Winteron, 2000). Clearly, needs exist to develop and use compatible management practices and to seek new agents that are capable of rapid population growth. About 19 of 43 species (Table 2) have been indentified as potential control agents because of the damage they cause or because of their narrow host range (Perkins, 1974). This list suggests that there are additional safe and effective agents among those already known, while others remain to be discovered. Host Range Tests and Results The two weevil species (N. eichhorniae and N. bruchi) have been released on waterhyacinth in 30 and 27 countries, respectively. Both have been subjected to extensive screening. They have been tested against 274 plant species in 77 families worldwide (Julien et al., 1999). Some use of a few non-target species, mainly other Pontederiaceae, was observed that was insignificant when compared to waterhyacinth. The other agents released on waterhyacinth, the fungus Cercospora piaropi Tharp, the mirid Eccritotarsus catarinensis (Carvalho), the moths N. albiguttalis and Xubida infusellus (Walker), and the mite O. terebrantis, have been introduced to fewer countries and have therefore been subjected to fewer host specificity trials. However, no host range extensions by these species have been recorded except for the predicted feeding by the weevils on pickerelweed Pontederia cordata L. (Center, 1982; Hill et al., 2000; Stanley and Julien, unpub). Post-release evaluations of natural enemies in countries of introduction can provide additional biosafety data and render further quarantine-based trials unnecessary. For example, field cage studies in Australia showed that the moth X. infusellus would harm pickerelweed (Julien, pers. comm.). It is therefore no longer being considered for release in the United States. On the other hand, the mirid E. catarinensis fed and developed on pickerelweed during quarantine trials in South Africa, but subsequent field trials in that country showed that it inflicted little damage to pickerelweed and didn’t readily colonize isolated pickerelweed stands (Hill et al., 2000). Many of the plant-feeding insects associated with waterhyacinth in South America utilize other species of Pontederiaceae (Table 2). Therefore, decisions for their release must rely on a risk-benefit analysis between the importance of native Pontederiaceae and the potential benefits offered by the natural enemy. Table 2. Characterization of Major Arthropods Associated with Waterhyacinth
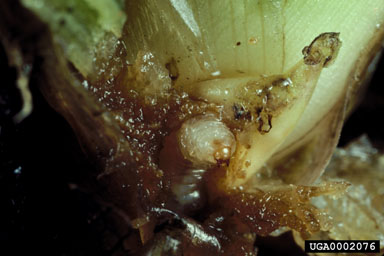
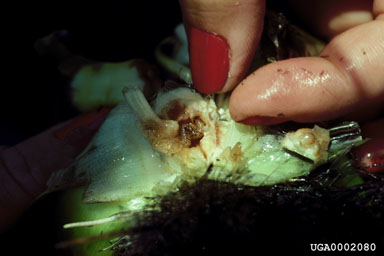
Releases Made Three insects, all originally from Argentina, have been released in the United States. The weevils N. eichhorniae and N. bruchi were released in Florida in 1972 and 1974, respectively, followed by the pyralid moth N. albiguttalis in 1977. Other Agents That Have Been, or Now Are, Under Consideration Three native North American species sometimes severely affect waterhyacinth populations, as well. These are the noctuid moth B. densa, the oribatid mite O. terebrantis, and the spider mite Tetranychus tumidus Banks. The moth X. infusellus has been rejected for release in the United States because it is clearly a threat to pickerelweed (DeLoach et al., 1980; Julien and Stanley, 1999). Cordo’s (unpublished report) conclusion that the arctiid Paracles tenuis Berg was polyphagous led to its rejection as well. Silveira Guido and Perkins (1975) and, later, Hill (unpub.) tested the grasshopper Cornops aquaticum (Bruner). Although Silveira Guido and Perkins (1975) considered it to be specific, concerns for pickerelweed precluded further consideration for release in the United States. The mirid E. catarinensis is still under consideration as the risk to pickerelweed seems minimal under field conditions (Hill et al., 2000), but information on its efficacy is needed for a proper risk-benefit analysis. Dolichopodid flies in the genus Thrypticus and planthoppers in the genera Megamelus and Taosa are now under consideration. Biology and Ecology of Key Natural Enemies Neochetina eichhorniae and N. bruchi (Coleoptera: Curculionidae) Members of the genus Neochetina are semiaquatic weevils that feed only on species of Pontederiaceae. Center (1994) reviewed the biologies of N. eichhorniae and N. bruchi. Adults of the two species (Fig. 5) are distinguished by the color and pattern of the scales on the elytra (Warner, 1970; DeLoach, 1975; O’Brien, 1976). Neochetina bruchi is typically brown with a tan band across the elytra. Neochetina eichhorniae is usually mottled gray and brown. Both species have two parallel tubercles on the elytra on either side of the mid-line, which are short and situated near mid-length on N. bruchi, but are longer and further forward on N. eichhorniae.
The whitish, ovoid eggs (0.75 mm in length) are embedded in plant tissue. Larvae are whitish with a yellow-orange head (Fig. 6). They have no legs or prolegs, only enlarged pedal lobes bearing apical setae. Larvae can be distinguished by the presence (N. bruchi) or absence (N. eichhorniae) of setal-bearing protuberences on these pedal lobes (Habeck and Lott, unpub. report). Neonate larvae are about 2 mm and fully-grown third instar larvae are 8 to 9 mm in length. Pupae are white and enclosed in a cocoon that is attached to a root below the water surface. Neochetina eichhorniae deposits eggs singly, whereas N. bruchi often deposits several in the same site. Neochetina bruchi prefers leaves with inflated petioles, especially those at the periphery of the plant (DeLoach and Cordo, 1976a), whereas eggs of N. eichhorniae are found in intermediate-aged leaves (Center, 1987a). Eggs hatch in seven to 10 days at 24°C. The first instar larva excavates a sub-epidermal burrow and tunnels downwards. There are three instars and late-instar larvae are generally found near the crown where they often damage axillary buds. The entire larval period requires 30 to 45 days with N. bruchi developing somewhat faster than N. eichhorniae (Center, 1994). The fully developed larva exits the plant and crawls to the upper root zone to pupate. The pupal stage requires about seven days, but teneral adults may remain in cocoons for extended periods. Emerging adults climb onto emergent plant parts to feed and mate, often aggregating within a furled expanding leaf or beneath membranous ligules. Females lay their first eggs soon after emergence (DeLoach and Cordo, 1976a, b). As many as 300 to 400 eggs are produced cyclically over a life span of up to 300 days (Center, 1994). Both species of Neochetina undergo flight muscle generation and degeneration (Buckingham and Passoa, 1985), possibly reflecting alternating dispersive and reproductive phases. Center and Dray (1992) theorized that plant quality and phenostage influenced the weevil’s propensity to switch between phases, with N. bruchi being more sensitive to plant quality (see also Heard and Winterton, 2000) and more likely to disperse. Adult feeding creates characteristic rectangular scars on the leaves, about 2 to 3 mm in width and of variable length, sometimes girdling the leaf petioles at the distal end and causing the blade to dessicate (see DeLoach and Cordo, 1983; Wright and Center, 1984; Center et al., 1999a). Moderate to severe weevil infestations cause plants to be shorter with smaller leaves, fewer offsets and flowers, lower tissue nutrient content, and reduced overall vigor (Fig. 7) than uninfested or lightly infested plants (Center and Van, 1989).
Eccritotarsus catarinensis (Heteroptera: Miridae) Eccritotarsus catarinensis (Fig. 8) is a leaf-sucking bug (2 to 3 mm long). Eggs are inserted into the leaf tissue parallel to the surface and the four nymphal instars feed gregariously with the adults on the underside of the leaves, causing severe chlorosis. Development of the eggs and nymphs requires 23 days and adults live 50 days (Hill et al., 1999). Bennett and Zwölfer (1968) observed a mirid on waterhyacinth in Belém, Brazil, but the insect was never collected or named. A mirid later collected in Rio de Janeiro during 1989 was identified as E. catarinensis. It was imported to quarantine in South Africa in 1992 from Canavieras, Brazil (Hill et al., 1999). More recently, it was found on the Kumaceba River in the upper reaches of the Amazon River, near Iquitos, Peru in 1999 (Cordo et al., unpub.). Host specificity of this mirid was determined in South Africa from tests using 67 species in 36 families. Some feeding and development occurred on three native African Pontederiaceae, (i.e., Eichhornia natans [P. Beauv.], Monochoria africana [Solms-Laubach], and Heteranthera callifolia Kunth.), but the risk to these plants was deemed minimal and the insect was released in 1997 (Hill et al., 1999, 2000). This insect was later imported to Australia, where additional host specificity testing was done. However, the potential for damage to native Australian Monochoria species precluded its use (Stanley and Julien, 1999). Some Monochoria species are serious weeds of rice paddies and not considered to have conservation value in Asia. As a result, E. catarinensis has been released in China (Ding et al., 2001) and imported into Thailand for pre-release evaluation (A. Winotai, pers. comm.). This mirid is being considered for release in the United States. However, host specificity trials in both South Africa and Australia demonstrated feeding and development on pickerelweed (Hill et al., 1999; Stanley and Julien, 1999). Pickerelweed, being an introduced plant in both of these countries, played no role in the decision to release this insect. But pickerelweed is native to North America, so any threat to it would be unacceptable in the United States. Several studies are therefore being undertaken in South Africa to quantify the impact of E. catarinensis on pickerelweed under field conditions. Eccritotarsus catarinensis is now established in South Africa (Hill and Cilliers, 1999) and its effects are being monitored. Although the impact of this insect on waterhyacinth performance has not yet been quantified, it does reach very high densities in tropical areas of the country where it is capable of causing severe die back of the plants (Fig. 9). It also has been released in Benin, Zambia, and Malawi, and cultures have been sent to Zimbabwe, Thailand, and China.
Niphograpta albiguttalis (Lepidoptera: Pyralidae) The small (ca. 0.3 mm), spherical, and creamy-white eggs of N. albiguttalis take three to four days to hatch at 25°C. The newly emerged larva (1.5 mm in length) is brown with darker spots and has a dark brown head (Fig. 10). There are five larval instars, the last of which grows to about 2 cm long, with a dark orange head and a cream-colored body covered with conspicuous dark brown spots. Larval development requires about two weeks. The fully-grown larva excavates a cavity in a healthy leaf petiole, in which it forms its cocoon. Pupation occurs in the cocoon and the pupal stage lasts seven to 10 days. The emerging adult moth exits the petiole through a silken tunnel prepared by the larvae before pupation. Adults (Fig. 11) live about seven to 10 days. Mating occurs shortly after emergence from the pupa and the female lays the majority of her eggs the following night. An average female will deposit 450 to 600 eggs. The entire life cycle requires three to four weeks. Center et al. (1982a) provide further information on the biology and identification of this species.
Orthogalumna terebrantis (Acarina: Galumnidae) The waterhyacinth mite, O. terebrantis (Fig. 12), like other mites, has piercing mouthparts with which it sucks plant juices. Its host plants include pickerelweed and waterhyacinth (Gordon and Coulson, 1969). Cordo and DeLoach (1975, 1976) described the biology and life history of O. terebrantis. Adults are shiny black, about 0.5 mm long and narrowed anteriorly. Females lay their eggs in small round holes chewed in the leaves. Eggs hatch in seven to eight days (at 25°C) and produce small (less than 0.24 mm), whitish, slow-moving larvae. Complete development requires about 15 days (at 25°C). Feeding damage is restricted to the leaf blades. Larval feeding causes small reddish spots on the leaf surface and the nymphs produce galleries that extend about 6 mm towards the apex. The adults emerge through round exit holes at the end of the gallery. Large mite populations produce up to 2,500 galleries on a single leaf, which desiccate the blade (Gordon and Coulson, 1969). Severe damage is usually localized or confined to a few plants but, when combined with other stresses, it can contribute to declines (Delfosse, 1978). Xubida infusellus (Lepidoptera: Pyralidae) Silveira Guido (1965, 1971) considered the pyralid X. infusellus (Fig. 13) to be one of the most important phytophagous species on waterhyacinth in South America. Larvae (Fig. 14) severely damage leaf petioles and can destroy shoots by feeding on apical meristems and burrowing into rhizomes. Although damage is similar to that of N. albiguttalis or B. densa, it was thought that the introduction of X. infusellus might complement the effects of N. albiguttalis (Bennett and Zwölfer, 1968; DeLoach et al., 1980). Xubia infusellus prefers advanced phenostage plants with elongate leaf petioles (see Center et al., 1999a), whereas N. albiguttalis prefers younger plants with inflated leaf petioles. Sands and Kassulke (1983) describe the adults in detail.
Silveira Guido (1965, 1971), DeLoach et al. (1980), and Sands and Kassulke (1983) provide notes on the life history of X. infusellus. The no Cturnal females lay egg masses in crevices such as the folds of leaves or the overlapping edges of furled leaves. Females lay indiscriminantly, sometimes on plants not used as larval hosts or, in the laboratory, on cage materials. Numbers of eggs per egg mass vary from a few to several hundred. Eggs hatch in six to seven days at 26°C. First instar larvae briefly feed externally, sometimes girdling a petiole before entering it, but then feed internally. They burrow downward, sometimes transferring to adjacent leaves, until they eventually encounter the rhizome. The number of larval instars varies from seven to ten, and development requires about 48 days (Sands and Kassulke, 1983). Larvae become about 25 mm long when fully grown (DeLoach et al., 1980). Late instar larvae form large burrows, causing extensive damage. Larvae cut emergence holes in the petiole prior to pupation that they close with silk, and then pupate just below the covered opening. The pupal stage lasts about nine days and total developmental requires 64 days at 26°C (Sands and Kassulke, 1983). The adult lives four to eight days (Silveira Guido, 1965, 1971; Sands and Kassulke, 1983). This insect has established in Australia (Julien and Griffiths, 1998). It also was released in Papua New Guinea (Julien and Stanley, 1999). A decision was made not to release it in the United States due to the threat to pickerelweed.
presence in Belize, Jamaica, Barbados, or St. Vincent. Mitchell and Thomas (1972) found members of the genus in Argentina, Uruguay, Brazil, Guyana, and Trinidad. The species found by Bennett and Zwölfer (1968) in northern South America was later identified as Thrypticus insularis Van Duzee (Bennett, 1976) and still later synonomized with Thrypticus minutus Parent (Dyte, 1993). However, this specific epithet was rarely referred to in later literature and the insect continued to be known as Thrypticus sp. Dr. Christian Thompson of the U.S. National Museum concluded that several Thrypticus species collected in Argentina probably represented undescribed species.
Cruttwell (1973) described the life history of a Thrypticus sp. from waterhyacinth in Trinidad. The adults are 1.5 to 2 mm long and light brown in color. Females lay eggs singly in young petioles of E. crassipes, inserting eggs into the tissues, usually just above the water line. Eggs are yellow, 0.5 mm long and 0.17 mm in diameter, curved, with one end narrower than the other. Petioles are suitable for oviposition only when recently separated from the sheath; thus all galleries in an individual petiole are of similar age. Eggs hatch in a few days and the larvae tunnel horizontally, making a second exit hole at the other end of the gallery. Larvae continue to feed in galleries, which they enlarge and lengthen. There are three instars and the larval stage lasts 35 to 42 days. Mature larvae are about 4 mm long. They prepare an emergence window in the petiole before pupating in an enlarged chamber below the exit hole. Adults emerge in seven to 12 days and lay up to 50 eggs.
Thrypticus were found attacking E. crassipes, E. azurea, P. cordata, and Pontederia rotundifolia L. in northern Argentina (H. Cordo, unpub.). Comparisons of genitalia and larval mining patterns of insects from various Pontederiaceae suggested that several distinct species were represented, some of which seemed restricted to waterhyacinth. The effects of the mining damage caused by Thrypticus species on waterhyacinth performance have not been measured. However, the strict monophagy, ubiquity, and abundance of these species make them promising as biological control agents. The tiny, but often abundant, tunnels produced by the larvae of these species have been judged trivial by some authors, but the damage may enhance the stress produced by other agents. The apparent high degree of specialization of Thrypticus species among species of Pontederiaceae suggests that they are host specific and augurs well for their potential use in biological control.
Females lay groups of 30 to 70 eggs enclosed in egg cases that are inserted into the youngest leaf petiole on a plant (Silveira Guido and Perkins, 1975). Eggs hatch in 25 to 30 days, producing green-and-red-striped nymphs (Fig. 18). There are six or seven nymphal instars and development requires about 50 days. Nymphs are highly mobile and very damaging. The dark green adults copulate soon after emergence, and produce up to eight egg cases 25 to 30 days later. Adults live up to 110 days, are mobile, strong fliers, and are extremely damaging to the plant (Fig. 19).
Cornops aquaticum feeds and develops on waterhyacinth, E. azurea, P. cordata, and Commelina sp. under laboratory conditions (Silveira Guido and Perkins, 1975). We observed C. aquaticum on E. azurea, P. cordata, P. rotundifolia and Pontederia subovata (Seub. in Markt.) Lowden, in addition to waterhyacinth (H. Cordo et al., unpub.) during field surveys in northern Argentina (1997) and the upper Amazon River in Peru (1999). This oligophagous species is clearly not suitable for release in the United States. Despite these results, C. aquaticum is under study in South Africa where its oligophagy, including development on P. cordata and Canna indica L. (Cannaceae), has been confirmed. Further large-scale, multi-choice trails will quantify the threat of C. aquaticum to African Pontederiaceae. Cercospora spp. (Hyphomycetes) Cercospora piaropi and Cercospora rodmanii Conway cause dark brown leaf spots on waterhyacinth that can lead to necrosis of older leaves and petioles. Characters used to separate these two species are variable, so these fungi may represent a single species (Morris, 1990). Cercospora piaropi, described in 1917 from Texas, was apparently introduced into the United States with the plant (Tharp, 1917). Extensive research has been conducted on the use of this species as a natural enemy of waterhyacinth (Freeman and Charudattan, 1984). Charudattan et al. (1985) investigated application techniques for C. rodmanii and concluded that this pathogen was unlikely to Control the plant with a single application. In 1986, Cercospora piaropi was found in South Africa associated with the decline of a waterhyacinth mat at a reservoir in the eastern province of Mpumulanga (Morris, 1990; Morris et al., 1999). It is now established throughout South Africa as the result of transplanting infected plants. Cercospora rodmanii was introduced to South Africa from Florida in 1988. Although these pathogens now occur widely in the western Cape province, there has been no resultant decline in weed populations. Other Species There are a number of other species about which little is known but which may have potential as control agents. They include the following:
Evaluation of Project Outcomes Establishment and Spread of Agents Neochetina eichhorniae was released in southern Florida in 1972, using eggs from 2,479 adults sent from Argentina during August 1972 to March 1973. Adults removed from founder colonies were then redistributed by numerous agencies. As a result, N. eichhorniae was released at 199 sites in Florida, 492 sites in Louisiana, one site in Texas, and four sites in California (Manning, 1979; Cofrancesco, 1984, 1985). This intensive effort seemed necessary because of the belief that this species didn’t fly. However, N. eichhorniae was already present when initial releases were made in Texas, having apparently dispersed from southern Louisiana, and by 1984 it was at several waterhyacinth infestations between Port Arthur and Corpus Christi (Cofrancesco, 1984; Stewart, 1987). Large numbers of weevils, many actively flying, were observed at lights in southern Louisiana during 1980 (Center, 1982), clearly indicating a capacity to disperse. When N. bruchi became available, there was no similar dissemination campaign. As a result, it was released at only 40 sites: 21 in Florida, 10 in Louisiana, five in Texas, and four in California (Manning, 1979; Stewart, 1985, 1987). Despite this disparity in release efforts, both species are now ubiquitous in Florida (Center and Dray, 1992; Center et al., 1999a), but the status of N. bruchi elsewhere is unclear. Niphograpta albiguttalis was initially released only in southern Florida, but populations dispersed more than 500 km within 18 months (Center, 1984). This moth was released at two sites in Louisiana during May 1979 and collected 27 km from the nearest release site a year later (Brou, undated). Niphograpta albiguttalis appeared to be absent from Texas in 1985, and so was released at a few sites during May 1986. It was widely dispersed by July 1986 (Stewart, 1987), probably originating from Louisiana, rather than the more recent Texas releases. DeLoach and Center (unpub.) found N. albiguttalis in Mexico near Veracruz and near Tapachula, the latter being on the Pacific coast near the border with Guatamala. This insect was never released in Mexico (Julien and Griffiths, 1998). So it is likely that these populations derived from ones in the United States, with the nearest release site being about 1,600 km away. Likewise, although there are no recorded releases of N. albiguttalis in Puerto Rico (Julien and Griffiths, 1998), larvae were collected near San Juan in 1995 (specimen records, Malaria Canal, 18 April, 1995, collector T. D. Center; Lago Loiza, 19 April, 1995, collector T. D. Center). Labrada (1996) reported its presence in Cuba, too, so perhaps N. albiguttalis “island hopped” from Florida to the West Indies. Suppression of Target Weed
carbohydrate reserves prevents clonal expansion. In recent experiments, for example, plots with weevils doubled or tripled in coverage, whereas unino Culated controls expanded nearly six-fold during the growing season (Center et al., 1999b). Hence, control stems from growth suppression, reduction of the seed bank, and destruction of existing plants.
The most recent and most spectacular effects of the waterhyacinth weevils have occurred at Lake Victoria in East Africa (Fig. 27). Waterhyacinth was first recorded on the lake in 1980 and by the mid-1990s some 12,000 ha of the weed were clogging bays and inlets. Uganda made the first introductions of N. eichhorniae and N. bruchi in 1995, followed by Kenya and Tanzania in 1997 (Anon., 2000). A significant reduction in the extent of the weed on the Ugandan shore was evident by November 1998, with many of the mats having sunk. These results were later repeated on the Kenyan and Tanzanian shores. An estimated 75% of the mats on the Kenyan side had sunk by December 1999 (Anon., 2000). The spectacular results of the biological control program on Lake Victoria using the two weevil species are the same as those achieved on Lake Kyoga (Uganda) (Ogwang and Molo, 1999) and on the lagoons of the Sepik River (Papua New Guinea) (Julien and Orapa, 1999). Similar results have been obtained in Sinaloa, Mexico where the release of N. eichhorniae and N. bruchi during 1995 to 1996 reduced 3,041 ha of waterhyacinth distributed over seven impoundments by 62% (to 1,180 ha) by 1998 (Aguilar, pers. comm.). These successes reaffirm earlier reports from Australia (Wright, 1979, 1981), Argentina (DeLoach and Cordo, 1983), India (Jayanth, 1987, 1988), and the Sudan (Girling, 1983; Beshir and Bennett, 1985).
Factors that Accelerate Success and Factors that Limit Control Factors associated with successful control include presence of the infestation in tropical and subtropical areas; infestations manifested as mono Cultures in free-floating mats that are able to sink when damaged; and mats that are stable (i.e., undisturbed) over long periods of time. Factors that might accelerate control include wave action, reduced growth (due to the action of biological control agents), and high nutrient levels (since high quality plants enhance insect population growth). Factors that limit control include removal of mats by herbicidal or mechanical means (thus disrupting agent populations), shallow water (damaged plants being unable to sink), ephemeral water bodies, toxicity effects in polluted waters, low temperatures at high-altitude or temperate sites, high nutrients at temperate sites, and limited releases (small, ino Culative releases as opposed to mass releases or serial releases) (Julien, 2001; Hill and Olckers, 2001). Recommendations for Future Work Future Needs for Importation or Evaluation Surveys done by Center et al. (1999a) confirmed that waterhyacinth populations not subjected to repeated control operations become stressed by biological control agents, particularly the two Neochetina species. On the other hand, water bodies subjected to Continual herbicidal control actions generally have small weevil populations, due to instability of the weevil’s food supply. Such sites produce healthier plants due to the reduced level of herbivore damage. The stressed plants typical of many unmanaged sites tend to be of lower nutritional quality than those at managed sites. The breeding condition of the female weevils correlates with host nutritional quality, so routine maintenance probably enhances the potential development of weevil populations by keeping host quality high, even though the actual populations are small. This suggests numerous possibilities for integrated approaches designed to overcome interference between the two Control methods. However, the present maintenance program is considered to be effective, efficient, and affordable. In contrast, an integrated program involving management of populations of biological control agents in concert with herbicide application would probably be more expensive, difficult to implement, and possibly less reliable. Hence, the present system is unlikely to Change. Therefore, new agents are needed to improve upon the level of biological control now realized. In particular, more mobile agents, with short life cycles and high reproductive capacities, are needed that can survive non-cyclical disruptions of waterhyacinth communities induced by herbicide applications. Currently, the candidates that best meet these criteria include the dolio Chopodid fly Thrypticus sp., planthoppers in the genera Taosa and Megamelus, and possibly the mirid E. catarinensis. Plans for Future Work Further work on the biological control of waterhyacinth is needed in five areas. First, available species should be fully evaluated. Second, additional natural enemies should be sought for use where existing control is less than desired. Third, more active approaches to biological control (e.g., mass or supplemental releases, serial releases) should be examined. Fourth, better methods to integrate biological control with other control options must be evaluated. Finally, the factors that accelerate success or limit control need further delineation. Despite a fairly long history of biological control of waterhyacinth in the world, and the number of successful programs now reported, much additional research is needed. As new agents are released there will be a need to quantify their impacts. In addition, some available agents have not been fully evaluated. Lack of a quantitative evaluation of O. terebrantis, for example, has resulted in it possibly being underrated as a control agent despite its significant effect on waterhyacinth on the Shire River in Malawi (Hill, unpublished data). A recent survey of the upper Amazon basin near Iquitos, Peru, identified several new candidate agents. The synergy observed between the insect damage and plant pathogens mandates further study. This brief trip was restricted to a small portion of the upper Amazon between Iquitos and Nauta. We do not consider this fruitful area to be fully explored and encourage further exploration. Surveys in other areas, such as the Pantanal region of Brazil and the Orino Co River system in Venezuela, also should be considered. Other insects that have been mentioned by explorers, for which basic information is not available, should be investigated to determine their field host plant ranges as a first step to assessing their potential for use in biological control efforts. These include the petiole-mining flies Eugaurax setigena Sabrosky (Diptera: Chloropidae), Hydrellia sp. (Diptera: Ephydridae), and Chironomus falvipilus Rempel (Diptera: Chironomidae); the flower-feeding carabid Calleida (= Brachinus); and the eriophyd mite Flechtmannia eichhorniae Keifer. The variable results given by biological control efforts against waterhyacinth in different areas have been ascribed to a lack of climate matching between the region of origin and the region of introduction (Hill and Cilliers, 1999). Investigations into the cold tolerances of the agents are required to determine their suitability for use in temperate areas. The biological control of waterhyacinth is perceived by water authorities to happen too slowly. Therefore, there have been a number of attempts to integrate biological control with other, quicker control options (herbicide application and mechanical control) (Delfosse et al., 1976; Center et al., 1982b, 1999a; Jones and Cilliers, 1999). The integration of two or more control options requires them to be compatible or, at least, not antagonistic. Further studies are needed to identify herbicides and adjuvants that are not toxic to the agents (e.g., Ueckermann and Hill, in press) and to determine more compatible methods of herbicide application. References APG (Angiosperm Phylogeny Group). 1998. An ordinal classification for the families of flowering Anonymous. 2000. Lake Victoria: Against the odds. Water Hyacinth News 1: 3-7. Baer, R. G. and P. C. Quimby, Jr. 1982. Some natural enemies of the native moth Arzama densa Barrett, S. C. H. 1977. Tristyly in Eichhornia crassipes (Mart.) Solms (water hyacinth). Biotropica 9: Barrett, S. C. H. 1980. Sexual reproduction in Eichhornia crassipes (water hyacinth). II. Seed Bennett, F. D. 1967. Notes on the possibility of biological control of the water hyacinth Eichhornia Bennett, F. D. 1972. Survey and assessment of the natural enemies of water hyacinth, Eichhornia Bennett, F. D. 1976. Current status of investigations on biotic agents of water hyacinth Eichhornia Bennett, F. D. and H. Zwölfer. 1968. Exploration for natural enemies of the waterhyacinth in northern Beshir, M. O. and F. D. Bennett. 1985. Biological control of waterhyacinth on the White Nile, Sudan, Bickel, D. J. 1986. Thrypticus and an allied new genus, Corindia, from Australia (Dipt.: Dolio Bock, J. H. 1968. The water hyacinth in California. Madrono 19(7): 281-283. Brou, V. A. Undated. Status in Louisiana of the introduced moth Sameodes albiguttalis (Warren) Buckingham, G. R. and S. Passoa. 1985. Flight muscle and egg development in waterhyacinth Center, T. D. 1976. The potential of Arzama densa (Lepidoptera: noctuidae) for control of Center, T. D. 1982. The waterhyacinth weevils. Neochetina eichhorniae and N. bruchi. Aquatics Center, T. D. 1984. Dispersal and variation in infestation intensities of waterhyacinth moth, Sameodes Center, T. D. 1987a. Do waterhyacinth leaf age and ontogeny affect intra-plant dispersion of Center, T. D. 1987b. Insects, mites, and plant pathogens as agents of waterhyacinth (Eichhornia Center, T. D. 1994. Biological control of weeds: Waterhyacinth and waterlettuce, pp. 481-521. In Center, T. D. and F. A. Dray. 1992. Associations between waterhyacinth weevils (Neochetina Center, T. D., and W. C. Durden. 1986. Variation in waterhyacinth/weevil interactions resulting from Center, T. D. and M. P. Hill. 1999. Host specificity of the pickerelweed borer, Bellura densa Walker Center, T. D. and N. R. Spencer. 1981. The phenology and growth of waterhyacinth (Eichhornia Center, T. D. and T. K. Van. 1989. Alteration of water hyacinth (Eichhornia crassipes [Mart.] Solms) Center, T. D., J. K. Balciunas, and D. H. Habeck. 1982a. Identification and descriptions of Sameodes Center, T. D., K. K. Steward, and M. C. Bruner. 1982b. Control of waterhyacinth (Eichhornia Center, T. D., A. F. Cofrancesco, and J. K. Balciunas. 1990. Biological control of wetland and aquatic Center, T. D., F. A. Dray, G. P. Jubinsky, and M. J. Grodowitz. 1999a. Biological control of Center, T. D., F. A. Dray, G. P. Jubinsky, and A. J. Leslie. 1999b. Waterhyacinth weevils (Neochetina Charudattan, R., S. B. Linda, M. Kluepfel, and Y. A. Osman. 1985. Bio Control efficacy of Cercospora Cock, M., R. Day, H. Herren, M. Hill, M. Julien, P. Neuenschwander, and J. Ogwang. 2000. Cofrancesco, Jr., A. F. 1984. Biological control activities in Texas and California, pp. 57-61. Cofrancesco, Jr., A. F. 1985. Biological control of waterhyacinth and alligatorweed in Galveston Cofrancesco, A. F., R. M. Stewart, and D. R. Sanders. 1985. The impact of Neochetina eichhorniae Cordo, H. A. 1999. New agents for biological control of water hyacinth, pp. 68-74. In Hill, M. P., Cordo, H. A. and C. J. DeLoach. 1975. Ovipositional specificity and feeding habits of the Cordo, H. A. and C. J. DeLoach. 1976. Biology of the waterhyacinth mite in Argentina. Weed Science Cronquist, A. 1988. The Evolution and Classification of Flowering Plants, 2nd edition. New York Cruttwell, R. E. 1973. Preliminary investigations on some insects causing minor damage to Delfosse, E. S. 1978. Effect on waterhyacinth of Neochetina eichhorniae [Col.: Curculionidae] Delfosse, E. S., D. L. Sutton, and B. D. Perkins. 1976. Combination of the mottled waterhyacinth DeLoach, C. J. 1975. Identification and biological notes on the species of Neochetina that attack DeLoach, C. J., and H. A. Cordo. 1976a. Life cycle and biology of Neochetina bruchi and DeLoach, C. J., and H. A. Cordo. 1976b. Ecological studies of Neochetina bruchi and N. eichhorniae DeLoach, C. J., and H. A. Cordo. 1983. Control of waterhyacinth by Neochetina bruchi (Coleoptera: DeLoach, C. J., H. A. Cordo, R. Ferrer, and J. Runnacles. 1980. Acigona infusellus, a potential De Quattro, J. 2000. Watch out water-hyacinth. Agricultural Research 48(3): 10-12. Ding, J., R. Wang, W. Fu, and G. Zhang. 2001. Water hyacinth in China: Its distribution, problems and Dyte, C. E. 1993. The occurrence of Thrypticus smaragdinus Gest. (Dipt.: Dolio Chopodidae) in Eckenwalder, J. E. and S. C. H. Barrett. 1986. Phylogenetic systematics of the Pontederiaceae. Forno, I. W. 1983. Life history and biology of a waterhyacinth moth, Argyractis subornata Freeman, T. E. and R. Charudattan. 1984. Cercospora rodmanii Conway. A Biocontrol Agent for Girling, D. J. 1983. Water hyacinth in the Sudan - early results. Biological Control News and Information Gopal, B. 1987. Water Hyacinth. Elsevier, New York. Gordon, R. D. and J. R. Coulson. 1974. Field observations of arthropods on waterhyacinth. Aquatic Gowanloch, J. N. 1944. The economic status of the waterhyacinth in Louisiana. Louisiana Gowanloch, J. N. and A. D. Bajkov. 1948. Water hyacinth program. Louisiana Department of Wildlife Goyer, R. A. and J. D. Stark. 1981. Suppressing water hyacinth with an imported weevil. Ornamentals Goyer, R. A. and J. D. Stark. 1984. The impact of Neochetina eichhorniae on waterhyacinth in southern Habeck, D. H. and D. M. Lott. 1993 (unpublished). Indentification of immature stages of weevils Hahn, W. J. 1997. Commelinanae. http://phylogeny.arizona.edu/tree/...ons/commelinanae/ Haigh, J. C. 1936. Notes on the water hyacinth (Eichhornia crassipes Solms) in Ceylon. Ceylon Haller, W. T., and D. L. Sutton. 1973. Effects of pH and high phosphorus concentrations on growth of Hansen, K. L., E. G. Ruby, and R. L. Thompson. 1971. Trophic relationships in the water hyacinth Harley, K. L. S. 1990. The role of biological control in the management of water hyacinth, Heard, T. A. and S. L. Winterton. 2000. Interactions between nutrient status and weevil herbivory Hill, M. P. and C. J. Cilliers. 1999. A review of the arthropod natural enemies,and factors Hill, M. P. and T. Olckers. 2001. Biological control initiatives against water hyacinth in South Africa: Hill, M. P., C. J. Cilliers, and S. Neser. 1999. Life history and laboratory host range of Eccritotarsus Hill, M. P., T. D. Center, J. N. Stanley, H. A. Cordo, J. Coetzee, and M. Byrne. 2000. The performance Hitchcock, A. E., P. W. Zimmerman, H. Kirkpatrick, Jr., and T. T. Earle. 1950. Growth and Holm, L. G., L. W. Weldon, and R. D. Blackburn. 1969. Aquatic weeds. Science 166: 699-709. Holm, L. G., D. L. Plucknett, J. V. Pancho, and J. P. Herberger. 1977. The World’s Worst Weeds: Jayanth, K. P. 1987. Suppression of water hyacinth by the exotic insect Neochetinaeichhorniae Jayanth, K. P. 1988. Biological control of water hyacinth in India by release of the exotic weevil Jones, R., and C. J. Cilliers. 1999. Integrated control of water hyacinth on the Nseleni/Mposa rivers and Julien, M. 2001. Biological control of water hyacinth with arthropods: a review to 2000, pp.8-20. In Julien, M. H. and M. W. Griffiths. 1998. Biological Control of Weeds. A World Catalogue of Agents Julien, M. H. and W. Orapa. 1999. Structure and management of a successful biological control Julien, M. H., and J. Stanley. 1999. Recent research on biological control for water hyacinth in Julien, M. H., M. W. Griffiths, and A. D. Wright. 1999. Biological Control of Water Hyacinth. ACIAR, Kiefer, H. H. 1979. Eriophyid studies C-17. Science and Education Adminstration, Agricultural Knipling, E. B., S. H. West, and W. T. Haller. 1970. Growth characteristics, yield potential, and Kohji, J., R. Yamamoto, and Y. Masuda. 1995. Gravitropic response. I. Eichhorniacressipes [sic] Kolbek, J. and J. Dostálek. 1996. Vegetation of water basins in the northern part of the Korean Labrada, R. 1996. Status of water hyacinth in developing countries, pp. 3-11. In Charudattan, R., R. Manning, J. H. 1979. Establishment of waterhyacinth weevil populations in Louisiana. Journal of Matthews, L. J. 1967. Seedling establishment of water hyacinth. Pest Articles and News Summaries (C) 13(1): 7-8. McVea, C. and C. E. Boyd. 1975. Effects of waterhyacinth cover on water chemistry, phytoplankton, Mitchell, D. S. and P. A. Thomas. 1972. Ecology of Water Weeds in the Neotropics. UNESCO, Paris, Morris, M.J. 1990. Cercospora piaropi recorded on the aquatic weed, Eichhornia crassipes, in South Morris, M. J., A. R. Wood, and A. den Breeÿen. 1999. Plant pathogens and biological control of Mullin, B. H., L. W. J. Anderson, J. M. DiTomaso, R. E. Eplee, and K. D. Getsinger. 2000. Invasive Muramoto, S., I. Aoyama, and Y. Oki. 1991. Effect of salinity on the concentration of some elements in O’Brien, C. W. 1976. A taxonomic revision of the new world subaquatic genus Neochetina. Annals of Ogwang, J. A. and R. Molo. 1999. Impact studies on Neochetina bruchi and Neochetinaeichhorniae O’Hara, J. 1967. Invertebrates found in water hyacinth mats. Quarterly Journal of the Florida Academy Parija, P. 1934. Physiological investigations on water-hyacinth (Eichhornia crassipes)in Orissa with Penfound, W. T. and T. T. Earle. 1948. The biology of the water hyacinth. Ecological Monographs 18: Perkins, B. D. 1974. Arthropods that stress water hyacinth. Pest Articles and News Summaries 20(3): 304-314. Richards, J. H. 1982. Developmental potential of axillary buds of water hyacinth, Eichhornia crassipes Sabrosky, C. W. 1974. Eugaurax setigena (Diptera: Chloropidae), a new stem miner in water hyacinth. Sands, D. P. A. and R. C. Kassulke. 1983. Acigona infusellus (Walker) (Lepidoptera:Pyralidae), an Schmitz, D. C., J. D. Schardt, A. J. Leslie, F. A. Dray, Jr., J. A. Osborne, and B. V.Nelson. 1993. The Seabrook, E. L. 1962. The correlation of mosquito breeding to hyacinth plants. Hyacinth Control Journal Silveira Guido, A. 1965 (Unpublished report). Natural enemies of weed plants. Final report on PL-480 Silveira Guido, A. 1971. Datos preliminaries de biologia y especificidad de Acigona ignitalis Hamps. Silveira Guido, A. and B. D. Perkins. 1975. Biology and host specificity of Cornops aquaticum (Bruner) Stanley, J. N. and M. H. Julien. 1999. The host range of Eccritotarsus catarinensis (Heteroptera: Stewart, R. M. 1985. Biological control of waterhyacinth in the California Delta, pp. 110-112. Stewart, R. M. 1987. Dispersing waterhyacinth bio Control agents in the Galveston District, pp. 105 Tabita, A. and J. W. Woods. 1962. History of hyacinth control in Florida. Hyacinth Control Journal Takhtajan, A. 1997. Diversity and Classification of Flowering Plants. Columbia University Press, Tharp, B. C. 1917. Texas parasitic fungi – new species and amended descriptions Mycologia 9: 105- Thorne, R. F. 1992. Classification and geography of flowering plants. Botanical Review 58: 225-348. Ueckermann, C. and Hill, M.P. 2001. Impact of herbicides used in water hyacinth control on the Ueki, K. 1978. Habitat and nutrition of waterhyacinth. Japan Agricultural Research Quarterly 12(3): 121- Ultsch, G. R. 1973. The effects of waterhyacinth (Eichhornia crassipes) on the microenvironment of USDA, NRCS (U.S. Department of Agriculture, Natural Resources Conservation Service). 1999. The Vogel, E. and A. D. Oliver, Jr. 1969a. Evaluation of Arzama densa as an aid in the control of Vogel, E. and A. D. Oliver, Jr. 1969b. Life history and some factors affecting the population of Warner, R. E. 1970. Neochetina eichhorniae, a new species of weevil from waterhyacinth, and Watson, M. A. 1984. Developmental constraints: effect on population growth and patterns of Watson, M. A., and Cook, C. S. 1982. The development of spatial pattern in clones of an aquatic Watson, M. A., and Cook, G. S. 1987. Demographic and developmental differences among clones of Webber, H. J. 1897. The water hyacinth, and its relation to navigation in Florida. Bulletin No. 18. U.S. Weber, H. 1950. Morphologische und anatomische studien über Eichhornia crassipes (Mart.) Weed Science So Ciety of America (WSSA). 1984. Composite list of weeds. Weed Science Wright, A. D. 1979. Preliminary report on damage to Eichhornia crassipes by an introduced weevil at Wright, A. D. 1981. Biological control of waterhyacinth in Australia, pp. 529-535. In Delfosse, E. S. Wright, A. D., and T. D. Center. 1984. Predicting population intensity of adultNeochetina eichhorniae Zeiger, C. F. 1962. Hyacinth – obstruction to navigation. Hyacinth Control Journal 1: 16-17. [ Contents ] [ Previous ] [ Next ] | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
| ||||
| |
The Bugwood Network and Forestry Images Image Archive and Database Systems The University of Georgia - Warnell School of Forestry and Natural Resources and College of Agricultural and Environmental Sciences - Dept. of Entomology Last updated on Thursday, July 19, 2018 at 01:38 PM Questions and/or comments to the | |||